桂林巖溶石山青岡群落植物功能性狀的種間和種內(nèi)變異研究
2021-11-18 02:42:26姚玉萍姚義鵬梁士楚李月娟梁歡歡趙慶年黃勇焙林才健
生態(tài)學(xué)報 2021年20期
何 雁,姚玉萍,姚義鵬,姜 勇,*,梁士楚,李月娟,梁歡歡,趙慶年,黃勇焙,林才健
1 廣西高校野生動植物生態(tài)學(xué)重點(diǎn)實(shí)驗(yàn)室,廣西師范大學(xué),桂林 541006
2 廣西師范大學(xué)生命科學(xué)學(xué)院,桂林 541006
3 桂林航天工業(yè)學(xué)院,桂林 541004
一個多世紀(jì)以來,生物多樣性的形成和維持機(jī)理即群落構(gòu)建機(jī)制一直是生態(tài)學(xué)研究的核心論題。相比大多數(shù)基于植物分類和數(shù)量的研究,植物功能性狀考慮了共存物種的冗余和互補(bǔ),反映了物種在功能屬性方面的差異,已成為揭示群落物種共存和多樣性維持機(jī)制的可靠途徑[1- 2]。植物功能性狀是指植物體具有的與其定植、生長、存活和死亡等緊密相關(guān)的一系列核心植物功能屬性,且這些屬性與環(huán)境適應(yīng)后表現(xiàn)出的性狀變異不僅能夠顯著影響生態(tài)系統(tǒng)功能,而且可以將群落結(jié)構(gòu)與群落環(huán)境、生態(tài)系統(tǒng)過程等聯(lián)系起來[3- 4]。通過植物功能性狀可以探明環(huán)境變異與物種分布之間的機(jī)理性關(guān)聯(lián)[5- 7],而探討功能性狀在種間和種內(nèi)尺度上是否存在差異及其形成原因是理解群落構(gòu)建的重要環(huán)節(jié)[8]。
目前,已有大量的研究表明種內(nèi)變異對群落動態(tài)變化以及生態(tài)系統(tǒng)特征具有顯著的影響,其與種間變異同等重要[9- 10]。Siefert等[11]對植物群落種內(nèi)性狀變異相對程度的全球薈萃分析表明,種內(nèi)性狀變異平均占群落內(nèi)總性狀變異的25%,占群落間總性狀變異的32%。不少研究表明,種內(nèi)性狀變異可能是物種定居新環(huán)境或抵抗環(huán)境變化能力的主要因素[12- 13]。種內(nèi)性狀變異可通過遺傳變異和表型可塑性來響應(yīng)局部選擇壓力使物種內(nèi)性狀發(fā)生適應(yīng)性變化,相較于利用物種性狀均值進(jìn)行的種間研究更能反映出植物種群的性狀變異能力[14- 15]。而Niu等[16]對西藏高山草甸的研究也表明了,種內(nèi)性狀變異在植物群落中的功能重要性會隨著環(huán)境的惡劣程度而增加。因此,將種內(nèi)性狀變異和種間性狀變異相結(jié)合才能更真實(shí)地反映群落構(gòu)建過程中物種對生境變化和資源競爭的響應(yīng),并更準(zhǔn)確地理解物種多樣性維持機(jī)制及生態(tài)系統(tǒng)特征[17- 19]。基于功能性狀的群落構(gòu)建機(jī)制通常將群落視為是確定性過程或隨機(jī)性過程或兩者結(jié)合的物種分選過程的結(jié)果[20- 22]。確定性過程側(cè)重于兩個選擇性過程:環(huán)境過濾和相似性限制(生態(tài)位分化)[23]。環(huán)境過濾被視為定向選擇的一種形式,即非生物環(huán)境選擇特定的性狀值,從而導(dǎo)致生境內(nèi)的性狀趨同[24]。相似性限制則被視為密度制約的一種選擇形式,通過競爭和其他生物相互作用以避免與相鄰物種的性征值過于相似[25]。而如果群落主要受到擴(kuò)散限制的影響,則隨機(jī)過程將起主導(dǎo)作用[21,26]。通常,檢驗(yàn)群落內(nèi)物種共存機(jī)制的方法可利用實(shí)際觀測的性狀分布與零模型隨機(jī)模擬得出性狀分布進(jìn)行比較[27]。近幾年來基于功能性狀結(jié)合零模型探討群落構(gòu)建機(jī)制的研究已經(jīng)有很多,但檢測水平大多還停留在物種水平、種群水平[18,28],綜合考慮種內(nèi)變異基于個體水平的零模型檢測還少有報道。
桂林巖溶石山地貌區(qū)生態(tài)系統(tǒng)特征典型,其巖溶發(fā)育強(qiáng)烈、生態(tài)系統(tǒng)穩(wěn)定性差、巖石裸露率較高,地形破碎、土層淺薄、土壤貧瘠、水分滲漏嚴(yán)重和石漠化加劇,是世界巖溶區(qū)域演化最具典型性和普遍性的代表地之一[29- 31]。由于該區(qū)域的生態(tài)環(huán)境惡劣,植物的生長與繁殖受到了限制[32]。青岡(Cyclobalanopsisglauca)為殼斗科青岡屬常綠闊葉樹種,具有較強(qiáng)的耐貧瘠、耐旱性、石生性和嗜鈣性等特點(diǎn),容易成為巖溶生態(tài)系統(tǒng)頂極群落的建群種,在巖溶森林植被中具有重要的地位和作用[33- 34]。鑒于此,青岡群落為探討巖溶森林植被的性狀變異及其背后的生態(tài)學(xué)機(jī)理提供了良好的研究對象。通過以桂林巖溶石山青岡群落木本植物為研究對象,基于4個植物功能性狀(葉干物質(zhì)含量LDMC、葉厚度LT、葉面積SLA和木材密度WD),采用混合線性模型結(jié)合方差分解的統(tǒng)計檢驗(yàn)分析功能性狀在種間和種內(nèi)2個尺度變異程度及比重;其次,針對其在種內(nèi)和種間2個尺度的變異及比重的不同,基于零模型(null model)方法檢驗(yàn)桂林巖溶石山青岡優(yōu)勢群落構(gòu)建機(jī)制,主要探討以下3個科學(xué)問題:(1)植物功能性狀的種間和種內(nèi)變異程度及比重如何?(2)結(jié)合個體性狀值將種內(nèi)變異納入考慮的群落機(jī)制檢測是否得到改善?(3)桂林巖溶石山青岡群落構(gòu)建過程中的潛在驅(qū)動機(jī)制是什么?通過對以上問題的研究,希望能初步揭示桂林巖溶石山青岡群落種內(nèi)種間性狀分布模式和維持機(jī)制,為桂林巖溶石山森林保護(hù)與管理提供科學(xué)依據(jù)。
1 材料和方法
1.1 研究區(qū)概況
研究區(qū)位于桂林巖溶區(qū)處于演替后期的青岡群落內(nèi),地處廣西壯族自治區(qū)東北部(110°14′—110°42′ E,24°43′—25°20′ N)。該區(qū)域?qū)儆谥衼啛釒駶櫦撅L(fēng)氣候,雨量充沛,氣候溫和,具有顯著的巖溶地區(qū)特性,年平均氣溫為18—19 ℃,最冷1月份平均氣溫為8 ℃,最熱8月份平均氣溫為28 ℃,全年無霜期達(dá)309 d,年降雨量為1856.7 mm,降雨量全年分配不均勻,春夏濕潤多雨,年平均蒸發(fā)量為1458.4 mm[35]。該區(qū)域喬木層主要以青岡為建群種,伴生種主要有扁片海桐(Pittosporumplanilobum);灌木層主要以一葉萩(Flueggeasuffruticosa)、粗糠柴(Mallotusphilippensis)、紅背山麻桿(Alchorneatrewioides)、干花豆(Fordiacauliflora)、檵木(Loropetalumchinense)等為主;木質(zhì)藤本以龍須藤(Bauhiniachampionii)為主。
1.2 群落調(diào)查
2017年7—9月在桂林陽朔縣葡萄鎮(zhèn)碑頭村(BTC)、桂林陽朔縣白沙鎮(zhèn)富里灣(FLW)、桂林市郊蘆笛巖張家村(ZJC)和桂林市郊演坡山(YPS) 4個采樣點(diǎn)共建立了20個20 m×20 m的樣方,各樣方基本概況如表1所示。調(diào)查時,將每個20 m×20 m的樣方劃分成4個10 m×10 m的小樣方,對每個小樣方進(jìn)行常規(guī)群落學(xué)調(diào)查,內(nèi)容主要包括:對喬木層植株進(jìn)行每木檢尺,記錄種名、空間坐標(biāo)、高度、胸徑、基徑、冠幅等指標(biāo);灌木層植株記錄種名、空間坐標(biāo)、高度、基徑等指標(biāo)。同時,記錄各樣方的土壤類型、海拔、巖石裸露率、礫石直徑、干擾程度等生境特征。所有樣方共記錄了4190株植物個體,隸屬36科,57屬,105種。其中喬木2372株,灌木1818株。
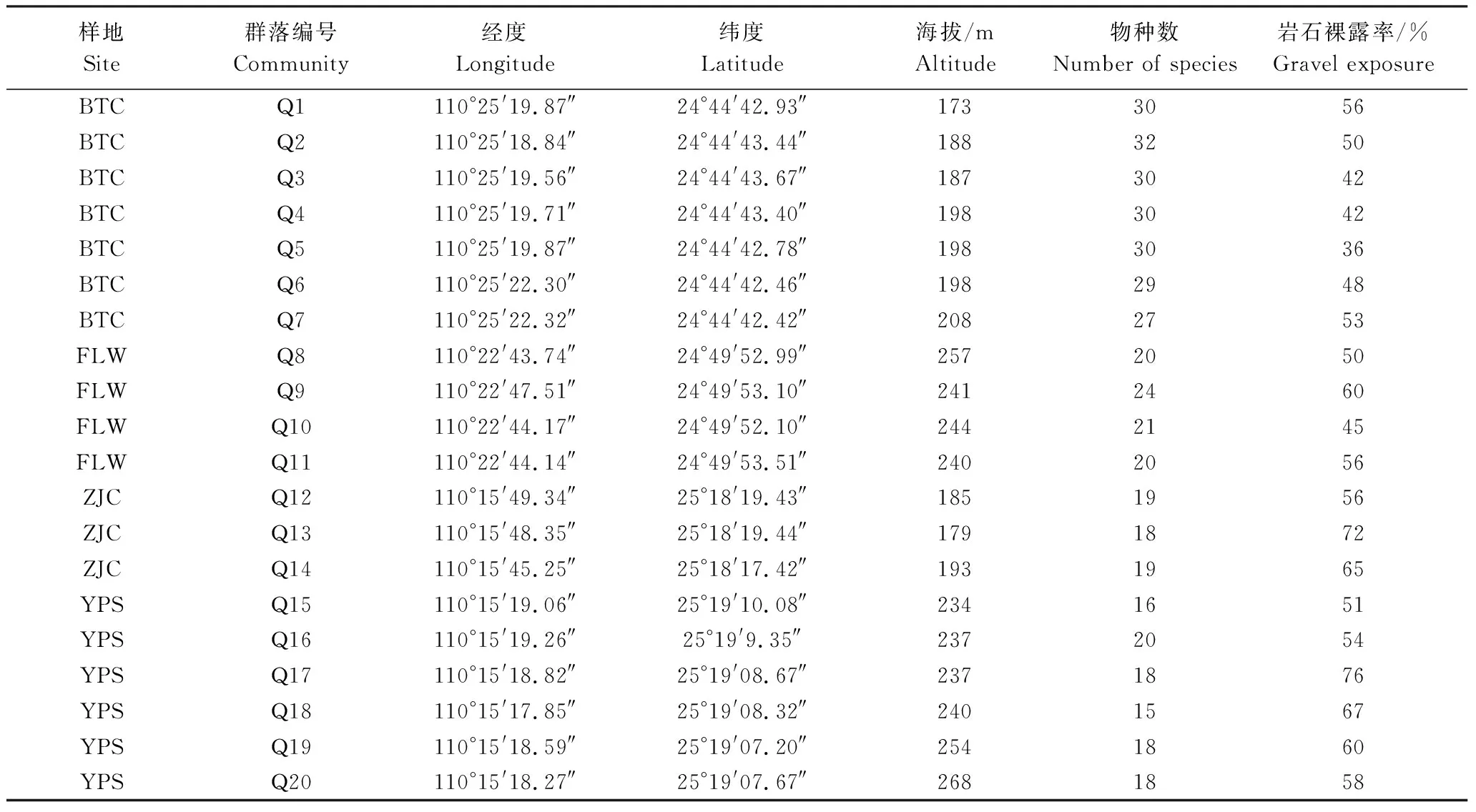
表1 樣地基本概況Table 1 Basic information of different sampling sites
1.3 功能性狀數(shù)據(jù)采集和測定
1.3.1功能性狀數(shù)據(jù)采集
由于植株的葉片生長狀況、個體發(fā)育階段、冠層位置以及蔭蔽度等會對植物的葉性狀差異造成影響,因此,本研究中采集了樣方內(nèi)所有胸徑≥1 cm的木本植物個體樹冠頂端3片完全展開的健康向陽葉片,為避免年份和季節(jié)等時間因素造成植物的功能性狀變異,研究數(shù)據(jù)均于2017年7—9月采集完成。木材樣本采自與葉片樣本相同的個體,使用枝剪截取樹冠外圍3小段非當(dāng)年生小枝。為保持葉片和枝條的水分飽和,室內(nèi)功能性狀測定均在樣本采回8小時內(nèi)完成。本研究共計有12570片葉片和木材樣品,來自105個種的4190株植物個體。
1.3.2功能性狀數(shù)據(jù)的測定
本研究選取葉干物質(zhì)含量(LDMC)、葉厚度(LT)、比葉面積(SLA)和木材密度(WD) 4個功能性狀進(jìn)行植物功能性狀的種間和種內(nèi)變異研究。其中,SLA和LDMC是葉片經(jīng)濟(jì)譜的核心性狀,代表植物獲取資源的能力;LT決定著葉片物理抵抗能力;WD是與機(jī)械穩(wěn)定性、生長速率以及耐旱和耐蔭策略相關(guān)的關(guān)鍵性狀[15, 36]。功能性狀各指標(biāo)測定方法如下:LDMC用精度為0.0001 g的電子天秤(BSM- 220.4,Zhuojing,China)稱量葉片的鮮重,然后將葉片樣品編號置于80 ℃烘箱中烘48 h 至恒重,稱量并記錄其葉片干重,根據(jù)葉片干重(g)與葉片鮮重(g)的比值獲得葉干物質(zhì)含量;LT用精度為0.01 mm的電子數(shù)顯卡尺(SF2000,Guilin,China)測量其葉厚度,每個葉片避開葉片主脈測量其前端、中端和末端,然后將三者的平均值作為所測葉片的葉厚度值;SLA由便攜式葉面積儀(Yaxin- 1241,Yaxin,China)測得葉片葉面積,由電子天秤測得烘干后的葉片干重,根據(jù)葉片面積(cm2)與葉片干重(g)的比值獲得比葉面積;WD根據(jù)Swenson和Enquist的研究[37],利用測定的枝條密度代替木材密度,即取回的枝條將其表面的樹皮削去后,用電子天平稱量并記錄枝條鮮重,利用量筒排水法測其體積,然后對其編號并放入80 ℃烘箱中烘干至恒重后,稱量并記錄枝條干重,根據(jù)枝條干重與體積的比值計算枝條密度(g/cm3)。
1.4 土壤取樣和理化性質(zhì)測定
本研究選取土壤含水量(SWC)、有機(jī)質(zhì)(SOM)、pH值(pH)、全氮(TN)、速效氮(AN)、全磷(TP)、速效磷(AP)、全鉀(TK)、速效鉀(AK)等9個土壤理化性質(zhì)進(jìn)行測定。在20 m×20 m的樣方中心位置處用環(huán)刀采集0—20 cm的土樣用于測定土壤含水量。土壤化學(xué)成分則采用“梅花五點(diǎn)法”采樣,在每個樣方的四個頂點(diǎn)和中心位置清除表層枯落物和腐殖質(zhì)層后,用土鉆鉆取0—20 cm表層土樣1 kg左右,運(yùn)用四分法混合均勻后裝入密封袋帶回實(shí)驗(yàn)室用于土壤化學(xué)性質(zhì)的測定。將采集的新鮮土樣放置于室內(nèi)陰涼通風(fēng)處自然干燥,待風(fēng)干后,剔除其中的根系、石塊、鈣核及動植物殘體等雜物后倒入研缽中研細(xì),過不同孔徑(0.15、0.25、1.4 mm)的土壤篩,對預(yù)處理后的土樣進(jìn)行分析測定。測定方法參照《土壤農(nóng)業(yè)化學(xué)常規(guī)分析方法》[38]。每個土壤樣品重復(fù)測定3次后取其平均值作為本研究分析的數(shù)據(jù)。
1.5 統(tǒng)計與分析
首先,本研究篩選出個體數(shù)大于3的71個主要物種進(jìn)行植物功能性狀的種間和種內(nèi)2個尺度上的變異分析。考慮數(shù)據(jù)需要滿足方差齊性和正態(tài)性,本研究分別對葉干物質(zhì)含量(LDMC)、葉厚度(LT)、比葉面積(SLA)和木材密度(WD)這4個性狀進(jìn)行對數(shù)轉(zhuǎn)換。利用混合線性模型和方差分解方法分析其在種間和種內(nèi)2個尺度上貢獻(xiàn)程度。該模型通過限制最大似然來擬合,相應(yīng)的性狀作為響應(yīng)變量,兩個尺度(即種間和種內(nèi))作為嵌套隨機(jī)因素。方差成分之間的比例表示每個尺度導(dǎo)致的變化的比例貢獻(xiàn)。針對4個性狀在種間和種內(nèi)2個尺度上貢獻(xiàn)的不同,本研究采用①性狀的范圍(Range)矩陣(作為檢驗(yàn)環(huán)境篩選的標(biāo)準(zhǔn))和②性狀軸上最近鄰體距離變異系數(shù)(CV_NND)矩陣(作為檢驗(yàn)相似性限制的標(biāo)準(zhǔn))在種間和種內(nèi)2個尺度上進(jìn)行觀測值與零期望9999次隨機(jī)模擬值比較分析。預(yù)計環(huán)境過濾和相似性限制會使觀測到的性狀值范圍和最近鄰體距離變異系數(shù)低于零期望值,本研究采用Wilcoxon符號單側(cè)秩檢驗(yàn)法比較觀測群落與隨機(jī)群落是否存在顯著差異性,進(jìn)而從統(tǒng)計學(xué)角度驗(yàn)證群落構(gòu)建機(jī)制理論。考慮到環(huán)境過濾對相似性限制檢驗(yàn)可能產(chǎn)生的附加影響,在隨機(jī)篩選性狀數(shù)據(jù)時執(zhí)行一個“限制零模型”,即從受約束的物種庫中隨機(jī)篩選物種性狀的分布范圍與指定群落性狀分布范圍相同。理論上,與隨機(jī)分布相比,環(huán)境過濾在理論上會改變實(shí)際群落的性狀均值以及引起性狀的范圍變窄;而相似性限制則會使物種沿性狀軸分布得更加均勻,因而會降低實(shí)際群落的最近鄰體距離變異系數(shù)[22, 39]。對于每個群落,利用SES值(standardized effect size)量化了實(shí)際群落性狀分布的標(biāo)準(zhǔn)值對隨機(jī)群落性狀分布零期望偏離的程度:
SES=(Iobs-Inull) /σnull
(1)
其中,Iobs代表實(shí)際群落的觀測值,Inull和σnull分別代表隨機(jī)群落的平均值和標(biāo)準(zhǔn)差,而SES值的正負(fù)則刻畫了實(shí)際群落性狀分布的標(biāo)準(zhǔn)值對隨機(jī)群性狀分布零期望偏離的方向。
功能性狀與土壤因子的關(guān)系采用多元逐步線性回歸分析,以土壤含水量(SWC)、土壤有機(jī)質(zhì)(SOM)、土壤pH值(pH)、全氮(TN)、速效氮(AN)、全磷(TP)、速效磷(AP)、全鉀(TK)、速效鉀(AK)等9個土壤理化性質(zhì)指標(biāo)為自變量,以葉干物質(zhì)含量、葉厚度、比葉面積、木材密度的群落加權(quán)平均性狀值(CWM)為因變量,根據(jù)AIC值、模型決定系數(shù)(R2)及P值大小選擇最優(yōu)模型,篩選出影響桂林巖溶石山青岡群落植物功能性狀的關(guān)鍵環(huán)境因子。以上所有數(shù)據(jù)統(tǒng)計分析與制圖均在R 4.0.2軟件中完成,其中混合線性模型的擬合用“nlme”包,方差分解用“ade”包的“varcomp”函數(shù),性狀的群落加權(quán)平均計算用“FD”包。
2 結(jié)果和分析
2.1 植物功能性狀的分布和變異
如表2所示,4個功能性狀的變異程度存在很大的差異,其中SLA的變異最大(60.81%),其次是LT(37.58%)和LDMC(25.13%),WD的變異最小(21.03%)。而方差分解和混合線性模型結(jié)果表明:4個植物功能性狀在種間和種內(nèi)2個尺度變異的大小表現(xiàn)出一致的規(guī)律(圖1),均體現(xiàn)種間>種內(nèi)。三個葉性狀(SLA,LT和LDMC)的變化主要來源于種間(50.42%—71.90%),種內(nèi)的貢獻(xiàn)較小(23.65%—42.61%);而木材性狀(WD)的變化在種間(47.82%)和種內(nèi)(44.86%)的貢獻(xiàn)相近。總體來看,桂林巖溶石山青岡群落植物功能性狀的種間變異(59.48%)要高于種內(nèi)變異(34.61%),種間變異是植物功能性狀變異的主要來源,但種內(nèi)變異亦不容忽視。
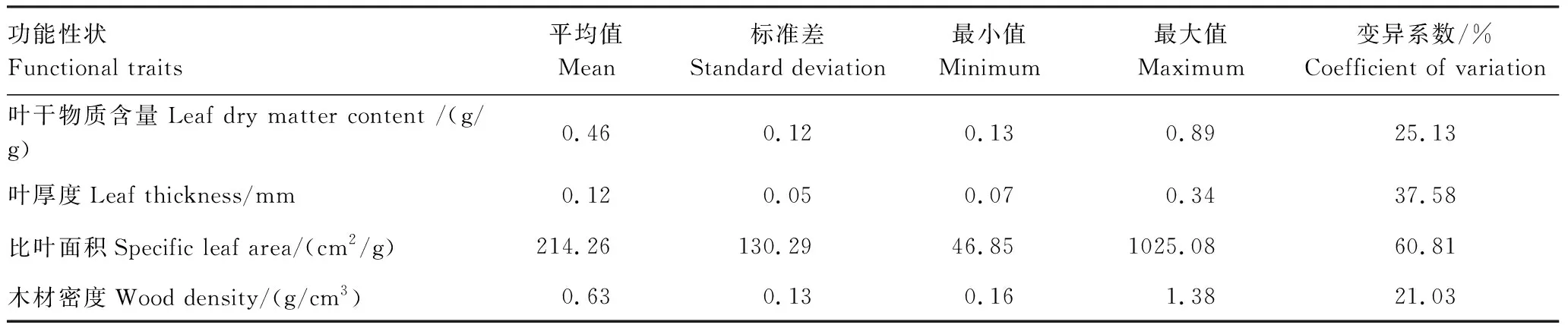
表2 4個植物功能性狀的分布特征Table 2 Distribution characteristics of 4 plant functional traits
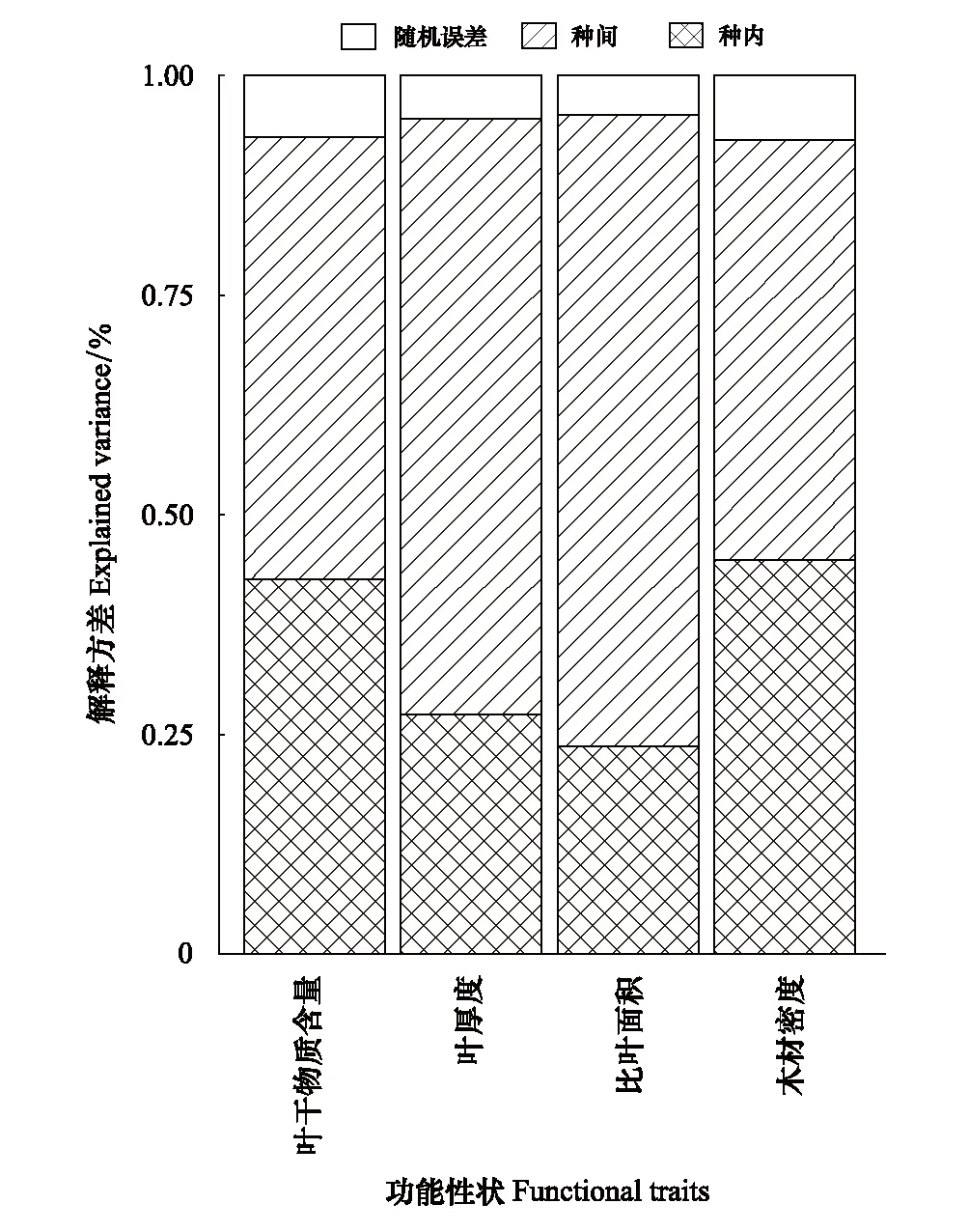
圖1 不同功能性狀在種間和種內(nèi)尺度上的方差分解圖 Fig.1 Variance partitioning of different functional traits at interspecific and intraspecific scales
2.2 零模型檢驗(yàn)
如圖2所示,在結(jié)合物種性狀均值種間尺度上的環(huán)境過濾檢驗(yàn)中,SLA和WD的分布范圍與隨機(jī)零模型相比顯著減小,呈現(xiàn)聚集分布,表明環(huán)境因子對植物的性狀分布具有限制作用;但是LDMC和LT的分布范圍與隨機(jī)零模型相比差異性不顯著。在種間尺度的相似性限制檢驗(yàn)中,僅WD的最近鄰體距離變異系數(shù)與隨機(jī)零模型相比顯著減小,呈現(xiàn)均勻分布,這與相似性限制理論預(yù)測的結(jié)果一致;但其余性狀與隨機(jī)零模型相比差異不顯著,呈現(xiàn)隨機(jī)分布。
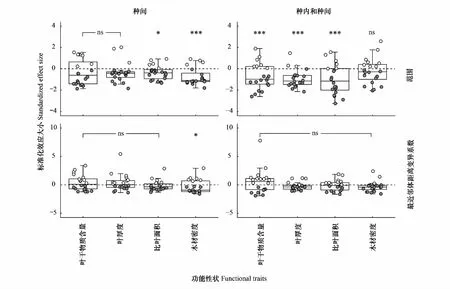
圖2 功能性狀在不同群落的零模型比較Fig.2 Comparison of null models of functional traits in different communities*代表觀測群落與零模型隨機(jī)群落在Wilcoxon符號秩檢驗(yàn)下呈顯著差異,* P <0.05,*** P <0.01,ns代表差異不顯著
在結(jié)合個體性狀值的種內(nèi)和種間尺度的環(huán)境過濾檢驗(yàn)中,LDMC,LT和SLA的分布范圍與隨機(jī)零模型相比顯著減小,呈現(xiàn)聚集分布;WD的分布范圍與隨機(jī)零模型相比差異不顯著。在種內(nèi)和種間尺度上的相似性限制檢驗(yàn)中,LDMC,LT,SLA和WD的最近鄰體距離變異系數(shù)與隨機(jī)零模型相比差異都不顯著,呈現(xiàn)隨機(jī)分布。
2.3 功能性狀與土壤因子的關(guān)系
多元逐步線性回歸分析結(jié)果表明,桂林巖溶石山青岡群落植物功能性狀與土壤有機(jī)質(zhì)、含水量、pH和氮磷鉀含量等環(huán)境因子密切相關(guān)(表3)。其中,葉干物質(zhì)含量與土壤含水量、全磷和全鉀含量顯著相關(guān);葉厚度與土壤全磷、含水量、有機(jī)質(zhì)和全鉀含量顯著相關(guān);比葉面積與土壤pH顯著相關(guān);木材密度與土壤全磷、全氮和全鉀含量顯著相關(guān)。

表3 功能性狀與土壤養(yǎng)分多元逐步回歸分析Table 3 Multiple linear stepwise regression analysis of function traits with soil nutrients
3 討論
3.1 植物功能性狀的種間和種內(nèi)變異分析
從研究結(jié)果來看,不同性狀的變異程度表現(xiàn)出了較大的差異,這表明不同功能性狀受到的環(huán)境作用的影響程度是存在差異的。通過4個性狀在種內(nèi)和種間水平上的變異檢測,進(jìn)一步表明了桂林巖溶石山青岡群落植物功能性狀的主要變異來源是種間變異。其次,相比木材性狀(WD),三個葉性狀(LDMC,LT和SLA)擁有更高的種間變異。植物性狀是由遺傳因素與環(huán)境條件共同決定的[40],不同遺傳背景的物種(分類單元不同)其性狀在種間差異較大,尤其是葉性狀[41]。本文葉性狀(SLA,LDMC和LT)在種間變異分析結(jié)果符合上述客觀規(guī)律,說明其主要受到遺傳因素限制,具有穩(wěn)定的變化特征。另一方面,本研究發(fā)現(xiàn)性狀的可塑性降低可能是由于傾向不利生境的專性化而引起的,表明低變異的物種更常見于惡劣的生境[42]。桂林巖溶石山相對惡劣的生境特點(diǎn),導(dǎo)致了其更傾向于選擇性狀可塑性低的物種[10, 33],這意味著植物對特殊環(huán)境的變化敏感,因而可能存在特殊的適應(yīng)環(huán)境方式。但木材性狀WD種內(nèi)變異的貢獻(xiàn)接近其種間的貢獻(xiàn),根據(jù)生態(tài)位理論,植物功能性狀的變異程度受環(huán)境過濾和相似性限制作用的共同影響[43- 44]。環(huán)境過濾作用能夠降低性狀的變異程度,使得具有相似性狀的物種在局域生境中共存,共存物種的性狀在功能上往往表現(xiàn)出趨同的一面,而相似性限制作用使這些共存物種通過種內(nèi)變異來增強(qiáng)生態(tài)位的分化,從而呈現(xiàn)某些生態(tài)策略的趨異性來降低生物競爭強(qiáng)度[28, 45]。對比本研究中的4個植物功能性狀,種間尺度的變異雖然占據(jù)了主導(dǎo)地位,但也有如WD種內(nèi)變異貢獻(xiàn)(44.86%)接近其種間貢獻(xiàn)(47.82%)的性狀。這表明了性狀的種內(nèi)變異并非是無足輕重,而是重要且不容忽視的變異來源之一。故在今后的基于性狀的生態(tài)學(xué)研究中,不能簡單地使用物種水平的功能性狀平均值代替?zhèn)€體數(shù)據(jù)而忽略種內(nèi)變異,應(yīng)基于個體水平采樣并整合功能性狀的種內(nèi)和種間變異來研究植物對環(huán)境的適應(yīng)策略,以期更好地揭示植物群落構(gòu)建和生物多樣性維持機(jī)制。
3.2 種內(nèi)種間變異對群落構(gòu)建的響應(yīng)
通過比較零模型結(jié)果來看,種內(nèi)變異對改善群落構(gòu)建機(jī)制的檢測有著一定的影響。基于個體水平將種內(nèi)變異納入考慮的環(huán)境過濾檢測確實(shí)得到了改善,三個葉性狀都表現(xiàn)出更強(qiáng)的收斂模式,極顯著低于零模型模擬。但與Siefert等[19]在美國紐約廢棄農(nóng)田的植物群落中的實(shí)驗(yàn)結(jié)果相似,在使用個體性狀數(shù)據(jù)對相似性限制檢測沒有得出相似的結(jié)論,性狀在局域群落中更趨向于隨機(jī)分布而不是均勻分布。這一研究結(jié)果表明在桂林巖溶石山青岡群落種內(nèi)和種間尺度上性狀分布除了受到環(huán)境過濾這個確定性過程的主要影響外,還受到一定的以擴(kuò)散限制為主的中性隨機(jī)過程的影響。這可能與桂林巖溶石山相對特殊(極端)的生境有關(guān)。一方面,研究表明物種在所處生境受到的脅迫性越高所需的耐受性也就越高,那么其性狀專化程度亦會相應(yīng)的表現(xiàn)得較高[42, 46],因此巖溶區(qū)的植物群落容易表現(xiàn)出較強(qiáng)烈的環(huán)境過濾作用;另一方面,正如Conradi等[47]在溫帶草原上的田間試驗(yàn)發(fā)現(xiàn):隨機(jī)過程是營養(yǎng)貧瘠土壤上群落物種組成的有效驅(qū)動力,因此在巖溶石山相對貧瘠的土壤環(huán)境上隨機(jī)過程必然也在起著一定的作用。同時,通過多元逐步線性回歸的功能性狀與土壤因子的關(guān)系分析,發(fā)現(xiàn)土壤有機(jī)質(zhì)、含水量、pH和氮磷鉀含量等環(huán)境因子與功能性狀之間密切相關(guān),這一結(jié)果也很好地佐證了環(huán)境過濾在桂林巖溶石山青岡群落構(gòu)建中的驅(qū)動作用。
此外,本研究發(fā)現(xiàn)在種間尺度的零模型檢驗(yàn)中,WD不僅體現(xiàn)環(huán)境篩選,同時體現(xiàn)限制相似性作用,但在種內(nèi)和種間尺度的零模型檢驗(yàn)中WD卻顯示隨機(jī)的分布格局。根據(jù)Albert等[48]提出的“性狀空間變異分割假說”,隨著空間尺度的增加,環(huán)境異質(zhì)性和個體數(shù)量逐步增加,表型可塑性也隨之增加,從而導(dǎo)致種內(nèi)變異的程度逐步增大,而種內(nèi)變異與環(huán)境過濾過程呈負(fù)相關(guān),與相似性限制作用成正比。因此,本研究推斷這可能是由于考慮了種內(nèi)變異,相似性限制作用的檢測得到了加強(qiáng),其與環(huán)境過濾的作用力相互抵消,導(dǎo)致了隨機(jī)格局的產(chǎn)生。盡管確定性機(jī)制似乎是群落構(gòu)建的重要因素,但在自然系統(tǒng)中,只要其他因素施加有意義的影響,它們的信號就可能被稀釋和掩蓋,要令人滿意地解釋在自然群落中觀察到的群落構(gòu)建模式,必須綜合考慮確定性過程和隨機(jī)過程的共存模式[49]。共存理論的最新發(fā)展認(rèn)識到,隨機(jī)性過程和基于生態(tài)位的確定性過程在塑造群落中同時起作用[43, 50],但是要恒量這兩個過程在不同物種組,不同生境和生物群落以及不同空間尺度的相對貢獻(xiàn)仍然是一個困難。因此,在今后基于功能性狀的生態(tài)學(xué)研究中,注重在個體水平采樣將種內(nèi)性狀變異和種間性狀變異相結(jié)合的同時,亦要注重構(gòu)建生態(tài)位分化的確定性過程和中性作用的隨機(jī)性過程的耦合關(guān)系模型,量化環(huán)境過濾、相似性限制和擴(kuò)散限制作用的相對貢獻(xiàn)率,結(jié)合空間結(jié)構(gòu)、環(huán)境因子和系統(tǒng)發(fā)育深入探究植物功能性狀變異的來源與潛在驅(qū)動機(jī)制,以期更好地揭示植物對環(huán)境的適應(yīng)策略以及植物群落構(gòu)建和生物多樣性維持機(jī)制。
4 結(jié)論
綜上所述,本研究的結(jié)論如下:(1)桂林巖溶石山青岡群落植物功能性狀的種間變異最大,但種內(nèi)變異亦不容忽略;(2)基于個體性狀值來考慮種內(nèi)變異是有必要的,雖然沒有提高相似性限制作用的檢測能力,但環(huán)境過濾作用的檢測明顯得到了改善;(3)對于桂林巖溶石山青岡群落,環(huán)境過濾作用確實(shí)是群落構(gòu)建的重要驅(qū)動機(jī)制,與相似性限制作用和隨機(jī)作用共同塑造了以青岡為優(yōu)勢物種的群落構(gòu)建,不過要具體量化不同過程在群落構(gòu)建過程的相對重要性還是一個困難。因此,未來的研究中,應(yīng)注重在個體水平采樣將種內(nèi)性狀變異和種間性狀變異相結(jié)合,進(jìn)一步構(gòu)建生態(tài)位分化的確定性過程和中性作用的隨機(jī)性過程耦合關(guān)系模型,不斷引入和開發(fā)新模型新方法來量化環(huán)境過濾、相似性限制和擴(kuò)散限制作用的相對貢獻(xiàn)率,加強(qiáng)群落構(gòu)建機(jī)制的定量分析研究,為揭示植物對環(huán)境的適應(yīng)策略和理解群落構(gòu)建的具體過程提供深刻的見解。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學(xué)學(xué)報(自然科學(xué)版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
中國生殖健康(2019年11期)2019-01-07 01:28:02
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
