雷州半島紅樹林群落特征及其分布格局
2021-11-13 03:42:36唐秋霞王友紹
生態科學 2021年5期
唐秋霞, 王友紹
雷州半島紅樹林群落特征及其分布格局
唐秋霞1, 2, 3, 4, 5, 王友紹1,2, 3, 4, *
1. 中國科學院南海海洋研究所熱帶海洋環境國家重點實驗室, 廣州 510301 2. 中國科學院大亞灣海洋生物綜合實驗站, 深圳 518121 3. 南方海洋科學與工程廣東省實驗室(廣州) 廣東 廣州 511458 4. 中國科學院南海生態環境工程創新研究院, 廣東 廣州 510301 5. 中國科學院大學, 北京 100039
以我國雷州半島紅樹林為調查與研究對象, 采用樣方調查方法對紅樹林區系組成、群落結構和分布特征開展調查分析。調查結果表明: 雷州半島紅樹林的群落可分為24個類型, 其中白骨壤群落面積占比高達60.55%, 占有絕對優勢; 雷州半島紅樹林包括真紅樹植物8科13種, 占全國真紅樹植物物種的50%, 半紅樹植物8科10種, 占全國半紅樹植物物種的83.33%; Shannon—Wiener多樣性指數為0.09—1.08, 部分研究區域之間數值差異較大; 從向陸林緣至向海林緣, 物種多樣性沿潮間帶的變化趨勢比較一致, 為高潮間帶>中潮間帶>低潮間帶; 白骨壤為雷州半島重要的優勢種, 在高、中和低潮間帶均有分布, 聚集分布為高潮間帶<中潮間帶<低潮間帶; 與2008年雷州半島國家“908”專項紅樹林調查結果相比, 該地區所記錄的紅樹物種由14種增加到了23種, 受2008年寒害所影響而枯死的紅海欖又逐漸恢復; 附城鎮1995年引進無瓣海桑, 與鄉土種白骨壤、秋茄一起成為當地的優勢種, 說明無瓣海桑的引進對于當地紅樹林群落恢復具有一定的促進作用。雖然雷州半島物種組成較為豐富, 但是紅樹林群落結構仍然不夠合理, 對殘次天然純林進行改造和人工多物種造林, 是日后雷州半島紅樹林群落恢復和重建的重要方向; 同時也要加強鄉土紅樹植物物種恢復, 維持雷州半島紅樹林生態系統物種多樣性與生態平衡。
雷州半島; 紅樹林; 物種組成; 空間分布; 群落結構
0 前言
紅樹林是生長于熱帶和亞熱帶海岸和河口潮間帶、受周期性潮水浸淹的以紅樹植物為主體的常綠灌木或喬木組成的潮灘濕地木本生物群落, 具有高生產力、高歸還率、高分解率和高抗逆性特性(即: 具有“四高”特性)[1], 是兼具海洋和陸地特性的特殊類型的生態系統[2], 也是地球上具有高生產力和高生物多樣性的濕地類型之一[3], 為鳥類、魚類和其他海洋生物提供了豐富的食物和良好的棲息環境, 除此之外, 它還有助于維持河口海灣生態環境、防風護岸、凈化環境、保護、維持濕地生態系統[4]和維護近海漁業的穩產高產[5], 素有“海上森林”之稱。
紅樹林調查研究的兩個基本問題是面積變化和種類組成。近年來, 利用遙感技術研究監測紅樹林濕地面積和景觀的變化逐漸成為研究熱點[6], 如紅樹林濕地的分布和其變化特征[7-12]、紅樹林濕地的景觀演變[13-14]、紅樹林生物量潛力的估算[15]等; 種類組成方面, 主要側重于紅樹林的群系組成、群落結構特征和物種多樣性研究上[16-18]。與遙感的便捷相比, 野外調查耗時、耗力, 且成本也更高。
雷州半島的紅樹林面積在1956年為14027 hm2, 由于人類活動和自然變化的干擾, 其紅樹林面積1985年僅為5800 hm2; 通過人工恢復等手段, 紅樹林面積于1993年增至12422.9 hm2; 而后又因為近海圍墾養殖等, 紅樹林面積銳減, 到2001年下降至7305.8 hm2[19]。目前, 雷州半島仍是我國大陸擁有紅樹林面積最大和具有豐富物種多樣性的區域[20]。多年來, 對于雷州半島紅樹林的研究主要集中于某個時期或局部區域種群和群落特征水平上[19-20, 22], 本研究基于雷州半島紅樹林實地調查數據, 從整體尺度對其紅樹林區系組成、群落結構的動態變化特征進行分析, 以期為雷州半島紅樹林生物多樣保護和可持續發展提供科學依據。
1 材料與方法
1.1 研究地點
雷州半島(20°12′—21°35′ N, 109°30′—110°55′ E)位于中國大陸最南端, 東臨南中國海, 南與瓊州海峽隔海相望, 西臨北部灣。其地勢平緩, 西北高、東南低, 海拔多在1000 m以下; 屬于熱帶海洋季風氣候, 有明顯的干、濕季之分, 沿海地區夏秋季多臺風[20]; 半島的紅樹林多在河口、海灣和島嶼的背風一側分布, 地質主要為砂質海灘, 僅僅在河口還有海灣地區為泥沙質灘涂地[21]; 半島的東岸為不規則半日潮, 西岸為規則全日潮。
1.2 研究方法
2018年7—8月, 我們對雷州半島紅樹植物群落的結構特征進行了調查, 共選取具有代表性的斷面30個(圖1), 按照各地紅樹林區群落的分布情況沿垂直岸線方向設置斷面, 每條斷面間隔一定的距離設置樣方(高潮間帶、中潮間帶和低潮間帶), 共設置樣方256個, 每個樣方面積為10 m×10 m, 分別測量樣方內紅樹植物株高、冠幅、基干周長和胸徑, 并記錄植物種類、株數和郁閉度[22]。
1.3 數據處理
物種多樣性指數采用Shannon—Wiener多樣性指數[23]:
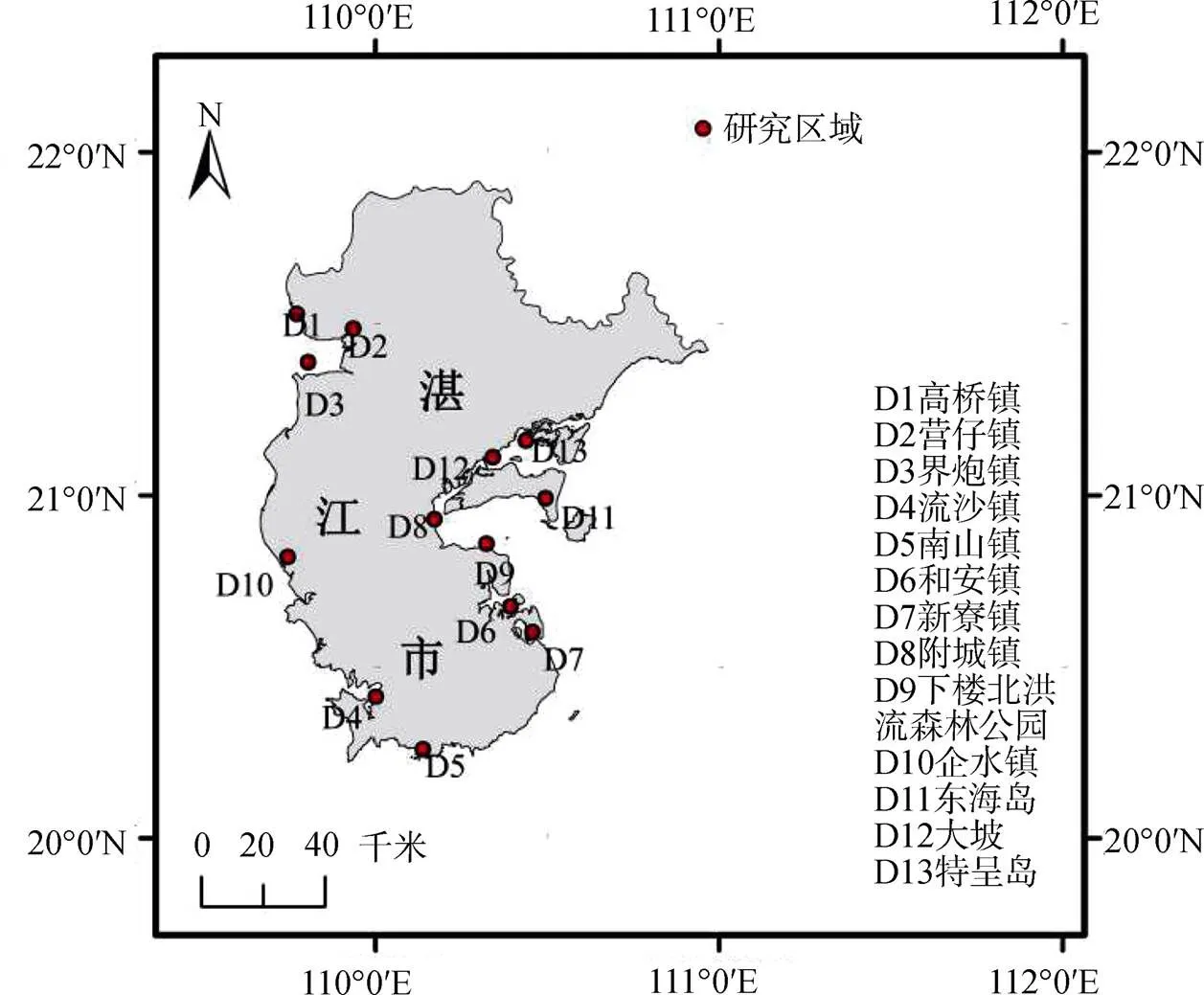
圖 1 各紅樹林片區在雷州半島的地理位置示意圖
Figure 1 Geographical map of each mangrove area in Leizhou Peninsula
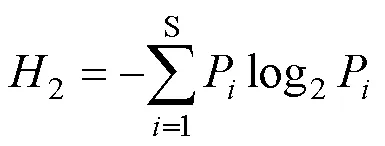
物種優勢度可以通過重要值來體現。重要值()為相對多度()、相對頻度()和相對顯著度之和()[22]:


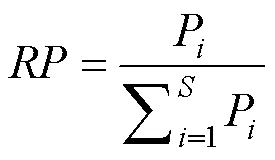
為物種數目;為所有物種的個體數之和;n第個種個體數量;
采用擴散型指數()、聚集度指標()、負二項指數()、擴散系數(S/x)和平均擁擠指數()5個指標來度量紅樹林種群的分布[24-25]。
采用SPSS 22、Excel軟件進行數據的統計和分析, 使用ArcGIS 10.2、Origin軟件進行圖件繪制。
2 結果與分析
2.1 雷州半島紅樹林群落分布與組成
表1表明, 雷州半島紅樹林群落可分為24個類型, 白骨壤群落、桐花樹群落、紅海欖群落、白骨壤+桐花樹群落等為主要群落類型, 占紅樹林總面積的94.53%, 其中白骨壤群落占比高達60.55%, 占有絕對優勢, 余下的13個群落類型占比皆低于1%。
紅樹林面積占比大于1%的群落分布特征如表2所示, 此外群落類型C12、C18—C21只在營仔鎮分布, 沿河岸上、中游處呈條帶狀分布。雷州半島紅樹林群落總體結構較為簡單, 多為小喬木狀或灌木林; 可分為天然林、人工林(附城鎮)和人工混合林(和安鎮)。從向陸林緣至向海林緣, 真紅樹植物群落都有分布, 其中白骨壤群落作為先鋒群落, 其面積最大; 在特呈島有較原始的白骨壤林, 冠幅較大(大于5 m×5 m), 基徑較粗, 而其他地區白骨壤基徑大多小于10 cm(如圖2所示)。半紅樹植物群落多見于高潮線之上, 面積均不大。
表3表明, 雷州半島紅樹林包括真紅樹植物8科13種, 占全國真紅樹植物物種的50%, 半紅樹植物8科10種, 占全國半紅樹植物物種的83.33%。雷州半島紅樹林物種占廣東紅樹林區系27種的85.19%, 高于粵東地區的66.7%[17]、淇澳島的81.5%[6]。
2.2 雷州半島調查區域各紅樹林區的物種多樣性指數和種群優勢度
表4為雷州半島各紅樹林調查區域的真(半)紅樹植物總數和物種多樣性指數, D1(高橋鎮)、D6(和安鎮)和D7(新寮鎮)的總物種數雖然最高, 但是主要組成物種白骨壤(高橋鎮為桐花樹和白骨壤)約占總數90%, 物種組成很不均勻、多樣性指數低; D13(特呈島)的總物種數最少, 為9種; D3(界炮鎮)的總物種數雖然只有10種, 但是其物種組成比較均勻、多樣性指數最高; D8(附城鎮)、D9(下樓北洪流森林公園)和D11(東海島)的物種組成比例也較為均勻。

表1 雷州半島紅樹林群落構成及面積比例
*表示該群落類型的面積占比低于0.3%。
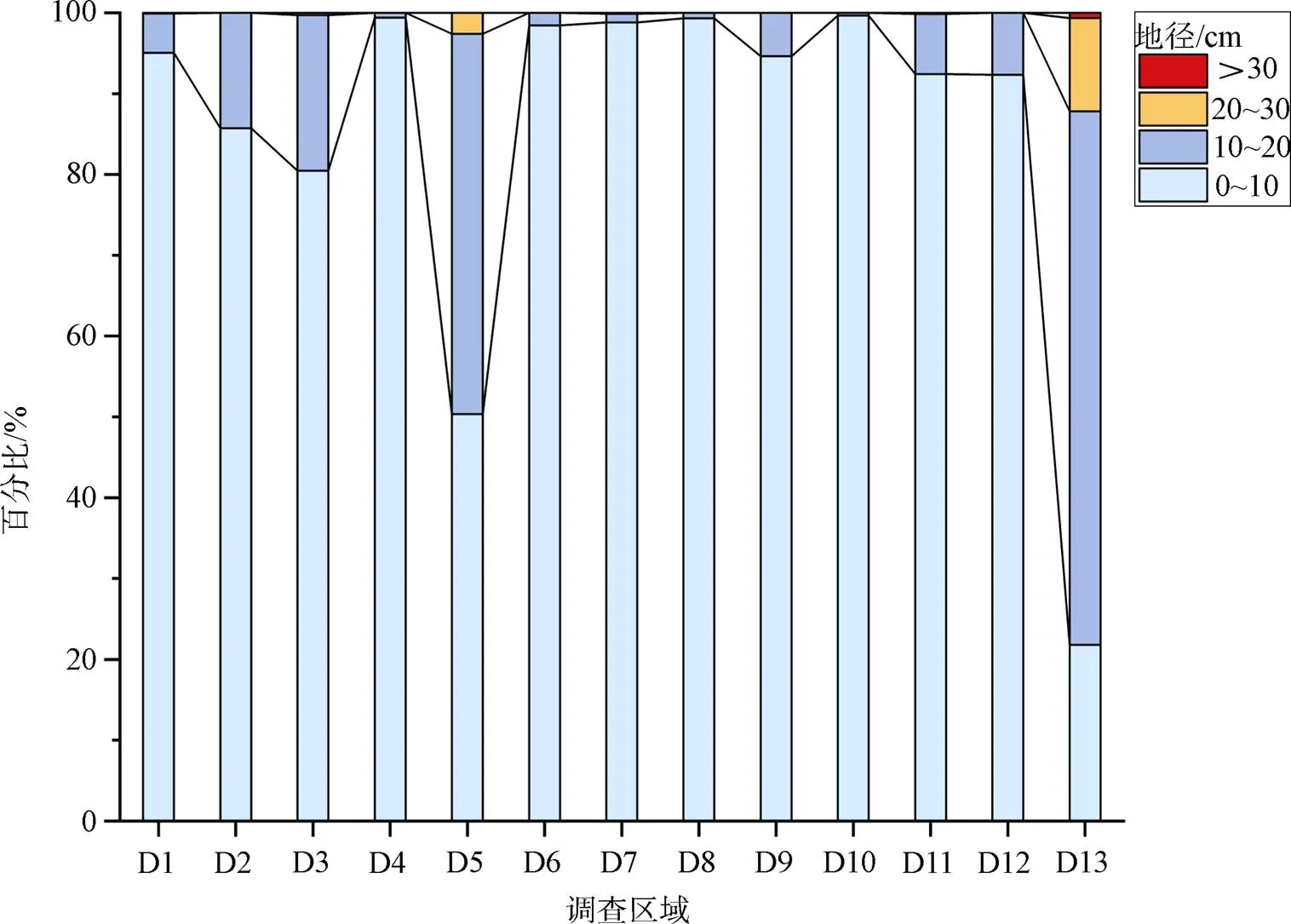
圖2 雷州半島各調查區域白骨壤地徑的組成比例
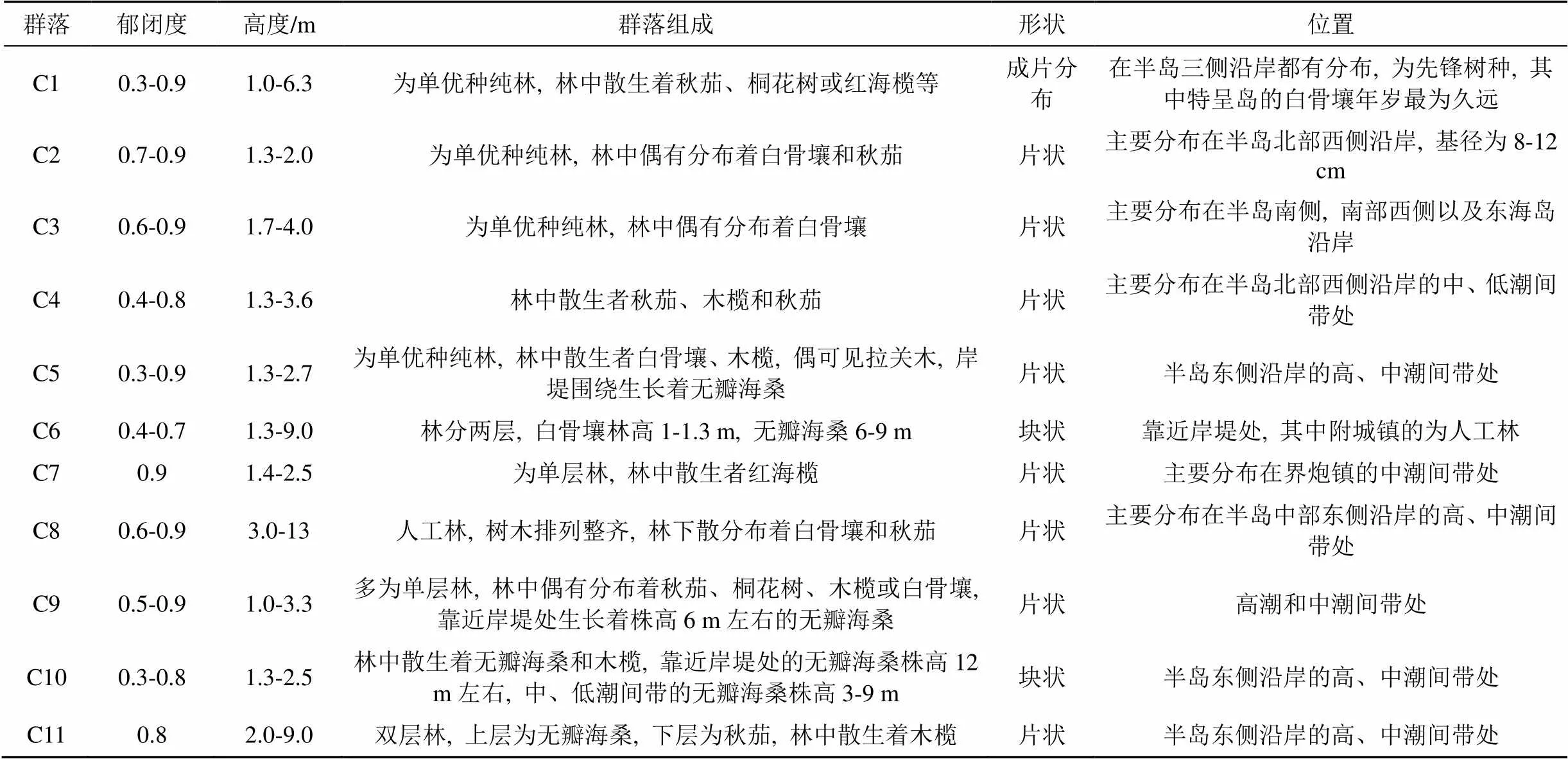
Table 2 Different composition of the base diameter ratio of Avicennia?marina in survey areas of the Leizhou Peninsula

表3 雷州半島調查區域紅樹林種類組成
“樣地調查”欄中標注“是”的11種紅樹植物是樣地調查所得, 標注“否”的12種則是野外踏查所得數據。
由圖2可看出, 白骨壤作為先鋒樹種, 適應生境能力強, 在不同潮帶均分布, 為雷州半島重要優勢種; 作為外來種的無瓣海桑生長良好, 在雷州半島中部東側成為優勢種之一; 角果木、老鼠簕、木欖、桐花樹和秋茄只在半島幾個潮灘形成優勢種; 半紅樹植物闊苞菊和苦郎樹只在營仔鎮形成優勢種; 紅海欖在整個雷州半島分布較為均勻。
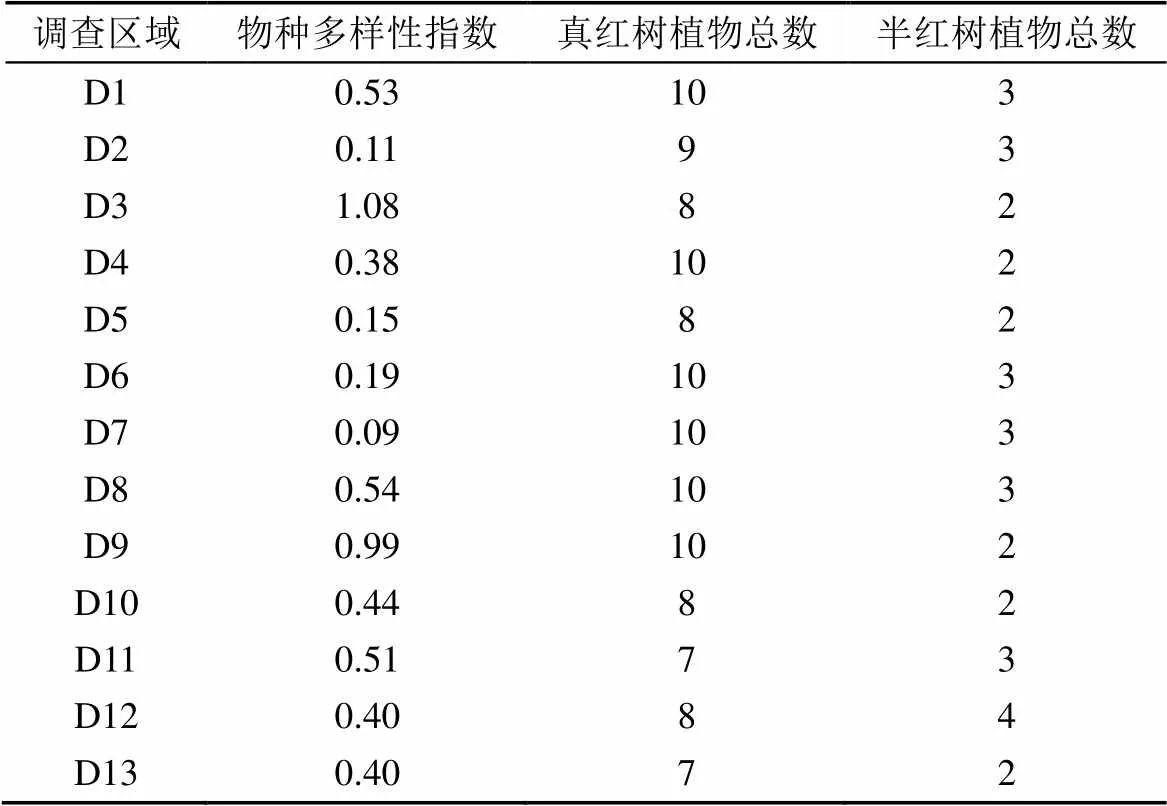
表4 雷州半島調查區域紅樹植物物種多樣性指數和真(半)紅樹植物總數
2.3 調查區域物種多樣性指數在潮灘上的變化
物種多樣性指數在潮間帶上的變化趨勢與當地的鹽度、沉積物、潮汐浸淹和波浪能量等海洋環境因素有關[26]。雷州半島各個紅樹林調查區的物種多樣性指數沿潮間帶的變化如圖4所示, H位于向陸林緣, L位于向海林緣。總體而言, 物種多樣性沿潮間帶的變化趨勢比較一致, 為高潮間帶>中潮間帶>低潮間帶。中、高潮間帶多為白骨壤和其他樹種的混交林, 低潮間帶則多為白骨壤純林。雷州半島真紅樹植物物種主要為白骨壤、桐花樹、秋茄和紅海欖: 白骨壤為群落先鋒物種, 在不同潮間帶均可見到, 是一個多潮帶的廣布物種; 紅海欖主要分布于中高潮間帶的內緣; 桐花樹和秋茄在中高潮間帶分布較廣。紅樹樹種的橫向分布主要是正面反映潮間帶潮汐浸淹頻率及相應產生的物理化學環境梯度的生理適應性, 但也不排除其他機制(包括胚軸潮汐分選、胚軸差別侵害、種間競爭和偶然性等[26-27])。
2.4 白骨壤在雷州半島的分布格局
除部分潮間帶之外, 白骨壤在雷州半島大多數紅樹林區中都有重要生態功能和社會經濟價值, 有些甚至遠遠超過其他樹種的, 故可以以白骨壤種群的分布格局來反映雷州半島紅樹林群落特征。
采用5種指標對雷州半島調查區域不同潮間帶白骨壤種群調查數據分析, 得到從向陸林緣至向海林緣白骨壤種群的分布格局(表5)。通過對擴散系數S/x中的檢驗變量、擴散指數中的檢驗變量X進行計算, 結果表明各種群均達到顯著性水平(t>t,X>X), 即不同潮間帶的白骨壤種群格局類型總體呈聚集分布的特點。根據指數值的大小對白骨壤種群在高、中和低潮間帶的聚集強度進行比較, 各指標的排序結果相近, 都顯示白骨壤種群在低潮間帶的聚集程度最高, 其次到中潮間帶, 最低為高潮間帶。
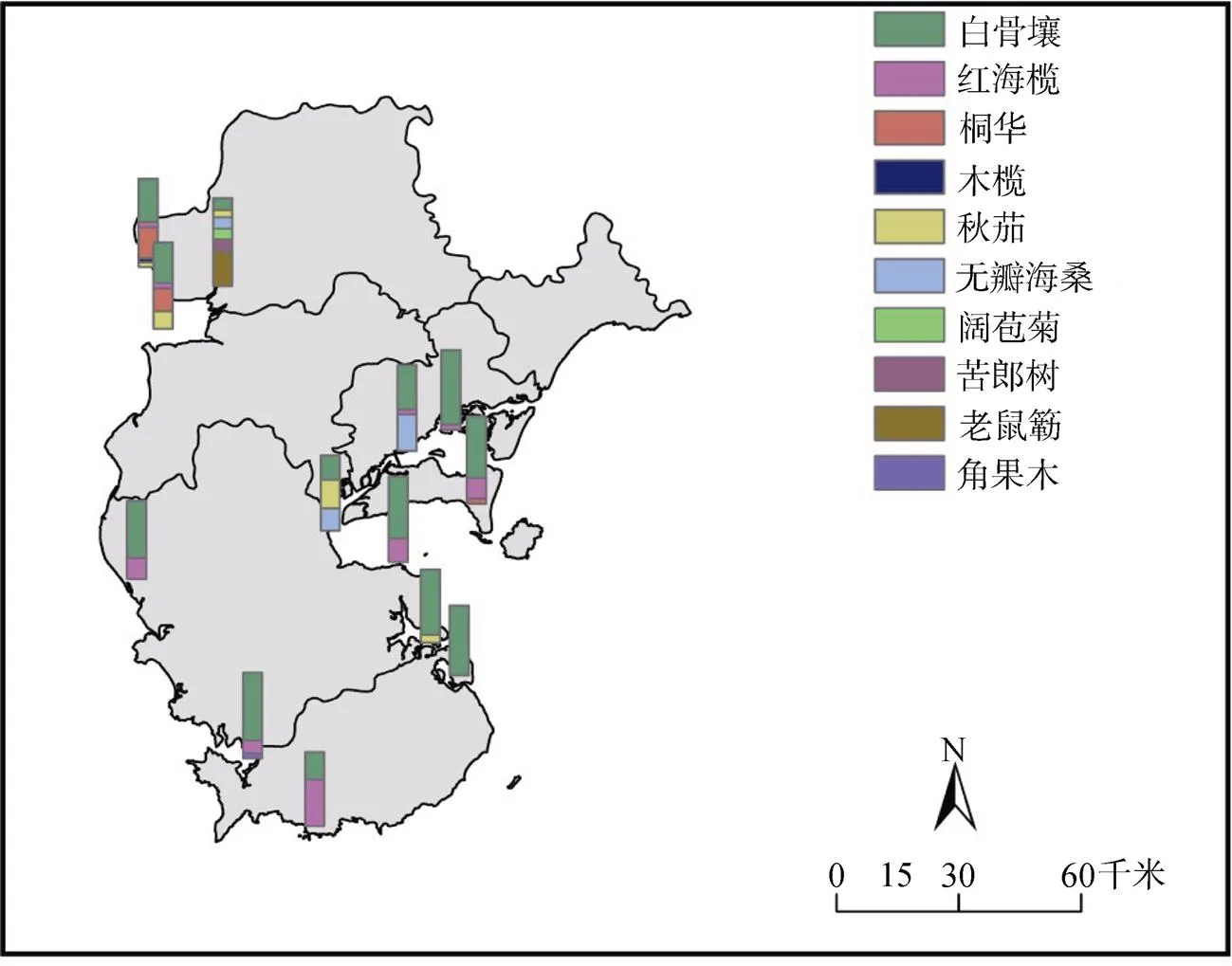
圖3 雷州半島調查區域紅樹植物優勢度
Figure 3 Dominance of mangrove plants in survey areas of the Leizhou Peninsula
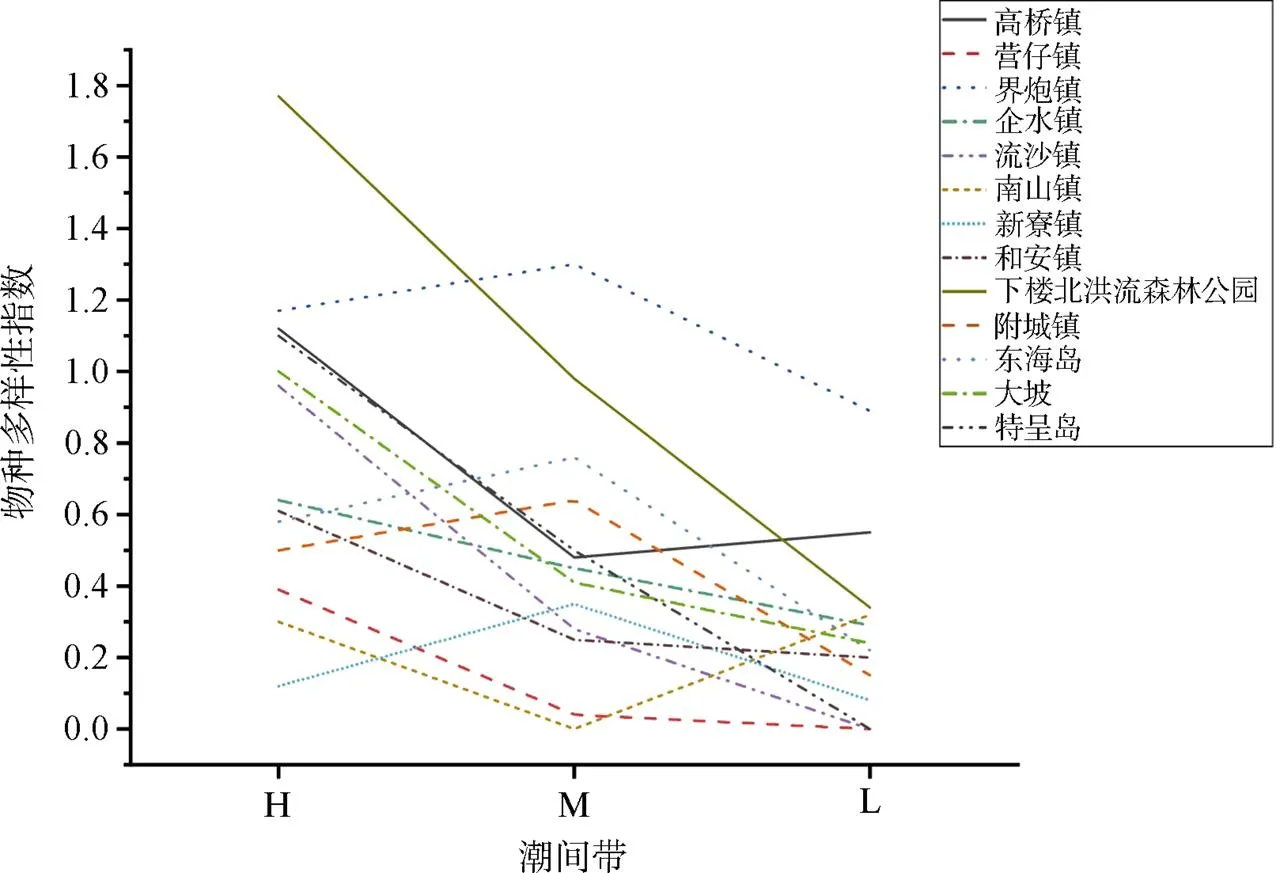
圖4 各紅樹林調查區物種多樣性指數沿潮灘的變化
Figure 4 Changes of Shannon index along tide in different mangrove in areas
潮灘外沿風浪較大, 環境較惡劣, 物種多樣性指數低, 相比于其他紅樹植物, 具有發達且附著能力強的呼吸根的白骨壤可以在漲潮之后的缺氧環境中生長[28]; 低潮間帶的白骨壤大多為較低矮的幼樹, 株高皆低于2 m, 直徑小于5 cm, 聚集分布更利于個體生存, 同時也有利于種群適應不良環境[29]。近陸林緣的物種多樣性最高, 個體對于環境比如光、養分的要求也進一步增強, 種內和種間的競爭激烈, 因而導致白骨壤種群密度下降, 聚集程度也最小。
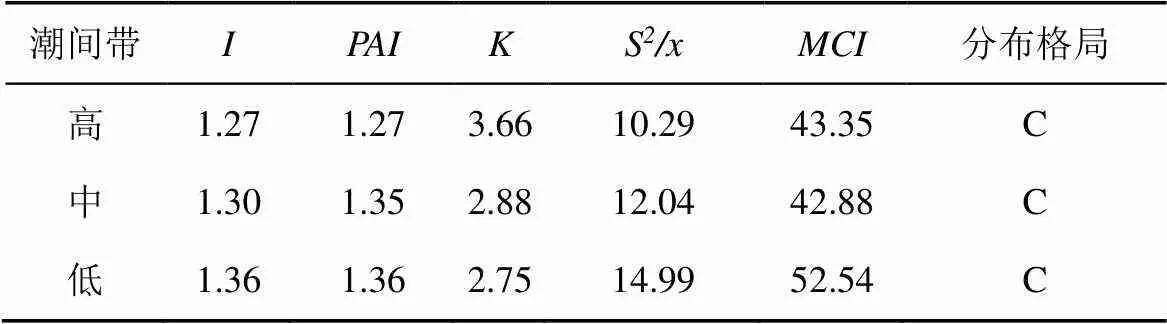
表5 調查區域不同潮間帶白骨壤種群分布格局指數
表示擴散型指數,表示聚集度指標,表示負二項指數,S/x表示擴散系數,表示平均擁擠指數, C表示聚集分布。
3 討論
3.1 雷州半島紅樹林調查區物種多樣性和真紅樹物種的變化
由表6可以看出, 與2008年雷州半島國家“908”專項紅樹林野外調查相比, 雖然每個研究區域真紅樹物種總數都增加了, 但是物種多樣性都降低了。造成這種現象的原因可能有兩個方面: 一方面, 在2008年, 各個區域的真紅樹物種總數雖然較少, 但是分布得比較均勻; 隨著時間的變化, 群落發生正向演替, 種內和種間競爭加劇, 導致林間分布稀疏, 群落密度下降, 演替后期紅樹植物物種如紅海欖、木欖和海漆等出現, 但數量較少, 且大多都沒有形成優勢種; 另一方面, 人類活動和全球氣候變化干擾, 也會對紅樹林的群落結構產生影響[30-32], 如區域不合理開發和利用等導致紅樹林減少、生物多樣性降低, 若隨著海平面上升, 且紅樹林沉積物的沉降速度小于海平面上升的速度, 這將將對紅樹林生存造成潛在的嚴重威脅[33-34], 先鋒紅樹植物將逐漸消失或向高潮間帶遷移[1], 尤其在我國絕大部分海岸地區建有固化防浪堤, 阻礙紅樹林向高潮間帶遷移[35]。
3.2 雷州半島群落類型的變化
在2008年雷州半島國家“908”專項紅樹林野外調查中, 紅樹林群落類型主要有13種, 白骨壤群落、桐花樹群落、秋茄群落、白骨壤+桐花樹群落和桐花樹+秋茄群落為主要的群落類型, 面積占比超過87%; 與2008年調查所得的結果相比, 白骨壤群落面積占比顯著增加; 雷州半島紅樹林群落類型變化較大, 新增了11種群落類型, 除了紅海欖群落之外, 其余面積占比皆低于3%。

表6 雷州半島部分紅樹林調查區物種多樣性指數和真紅樹物種的變化
“2008年數據”(“2018年數據”); √: 存在于調查區域, -: 不存在于調查區域。
由于2008年寒害的影響, 雷州半島大部分地區的紅海欖已經枯死[31], 造成單優種紅海欖林稀少, 多為紅海欖和其他紅樹物種的混交林, 且除高橋鎮之外, 其余的紅海欖皆為小樹。十年之后, 紅海欖在半島的地理分布相對比較均勻(如圖3所示), 面積占比5.86%, 在企水鎮、流沙鎮、南山鎮和東海島等區域都形成優勢種之一。其中, 在南山鎮的一個斷面中, 紅海欖為最主要優勢種, 其群落的面積占比具有絕對的優勢。中、高潮位區林高2.5—3 m, 林下分布著大量的紅海欖幼苗; 向海林緣處紅海欖株高較高, 皆為3.5—4 m, 林中偶有分布著株高2.7—3 m, 胸徑5—8 cm的白骨壤, 林下幼苗少。此紅海欖群落的年齡結構為典型的金字塔型, 即隨著樹齡的增加, 紅海欖個體所占比例相應地減少, 但種群在迅速增長。
3.3 雷州半島紅樹林群落的特征
雖然雷州半島擁有群落演替后期的真紅樹物種紅海欖、木欖、欖李和海漆, 但是只有紅海欖在雷州半島分布較廣, 木欖僅在高橋鎮的紅樹林自然保護區內形成優勢種, 而欖李和海漆多在向陸林緣零星分布, 規模很小; 總體上半島的天然半紅樹植物物種豐富度都較低, 且主要為黃槿、苦郎樹和闊苞菊, 群落的規模也不大; 演替先鋒群落—白骨壤、桐花樹和白骨壤+桐花樹群落的面積占比為71.88%; 受2008年寒害影響而死亡的紅海欖在十年間逐漸恢復, 主要是在原有斑塊上擴展, 多為單優種純林, 除此之外, 作為外來引種的無瓣海桑在一些群落上也主要為純林, 物種多樣性指數低, 群落結構穩定性差, 對于外界壓力的承受極限小, 需要進行人為干預。這表明雷州半島的紅樹林群落結構總體上仍然不夠合理, 無論是天然林還是人工林, 都多為純林; 或是白骨壤和其他群落演替初、中期的紅樹植物如桐花樹、秋茄的混合林, 群落演替后期的紅樹植物如紅海欖、木欖和欖李等無法形成優勢種, 整個雷州半島內, 只有高橋鎮的部分群落擁有較為完整的演替序列, 這可能與過去數十年間頻繁的人類活動比如圍墾養蝦、過度開發, 以及極端天氣等所造成的紅樹林面積變小、破碎化以及群落萎縮有很大的關系。
4 結論
與2008年國家“908”專項紅樹林調查結果相比, 經過10多年紅樹林林保護和發展, 雷州半島調查區域紅樹物種已由之前14種增加到23種, 增加了4種真紅樹植物物種, 分別為角果木、拉關木、木欖和尖葉鹵蕨(有些物種由此次踏查發現), 拉關木為引進的外來物種, 目前規模較小, 主要分布于高潮位區, 株高多在2 m以下, 地徑8 cm左右; 天然角果木僅在流沙鎮有較大規模分布, 與白骨壤和紅海欖成為當地紅樹群落的優勢種, 高橋鎮角果木林為人工造林, 且未能形成優勢種; 雖然作為紅樹群落演替后期物種之一的木欖在雷州半島各個紅樹林區域都有分布, 但除高橋鎮的木欖規模最大之外, 總體上都只是在近陸林緣零星分布; 尖葉鹵蕨僅在極少數地區的林下幾株分布; 5種半紅樹物種: 黃槿、苦郎樹、鈍葉臭黃荊、玉蕊和海濱角果木, 其中黃槿、苦郎樹和闊苞菊分布范圍廣, 為雷州半島主要的半紅樹物種。
雖然雷州半島物種組成較為豐富, 但是紅樹林群落結構仍然不夠合理, 2008、2018年的紅樹林調查結果都表明, 白骨壤群落在整個半島的面積占比都占有絕對的優勢, 沿潮間帶的分布都為聚集分布; 在大部分群落中, 無論是天然林或人工引種林, 皆以物種多樣性指數低、結構穩定性差的單優種純林為主; 紅海欖、木欖和欖李等這些群落演替后期的紅樹植物皆無法形成優勢種, 半紅樹和伴生植物種群的規模也都比較小, 表明在未來時間內, 如果繼續保持當前的紅樹林恢復模式, 雷州半島的紅樹林面積將會持續增加, 但是物種多樣性指數仍可能下降。若遇上比如海平面上升、極端天氣變化等情況, 雷州半島的紅樹林將會面臨嚴重威脅, 需要進一步加強全球氣候變化對紅樹林影響研究。
雷州附城鎮1995年人工造林, 引進外來物種無瓣海桑, 到如今人工林胸徑為13—20 cm, 株高12 m左右, 且與鄉土種白骨壤、秋茄成為當地的優勢種之一, 物種多樣性指數為0.54, 表明引進外來物種可以使紅樹林群落得到較好的發展, 但是也有研究表明, 無瓣海桑的引進會在一定程度上抑制鄉土種秋茄等生存和發展[6]。因此, 在今后雷州半島紅樹林的保護和恢復工作中, 除了考慮氣候、地質和水文等重要環境因素之外, 應將重點放在群落的物種多樣性的構建上, 借鑒紅樹林生態恢復成功模式, 盡量選取當地鄉土種來造林[35-36]; 同時, 雷州半島紅樹林易受海平面上升、極端天氣影響較為嚴重的區域[30, 35], 紅樹林恢復還要考慮到紅樹植物物種耐水淹、抗低溫和耐干旱等[1, 37-38]。對白骨壤純林或其他的殘次、低矮純林進行改造和人工多物種造林, 是日后雷州半島紅樹林群落恢復和重建的重要方向; 除此之外, 也應該進一步加強鄉土種紅樹植物物種恢復, 即在選取鄉土種造林時, 可以優先考慮原先生境受到威脅的物種, 遷地保育, 以維持雷州半島紅樹林生態系統的物種多樣性與生態平衡。
[1] 王友紹. 紅樹林分子生態學[M]. 北京: 科學出版社, 2019.
[2] 林鵬. 中國紅樹林生態系[M]. 北京: 科學出版社, 1997.
[3] 黎夏, 葉嘉安, 王樹功, 等. 紅樹林濕地植被生物量的雷達遙感估算[J]. 遙感學報, 2006, 10 (3): 387–396.
[4] BAYEN S, WURL O, KARUPPIAH S, et al. Persistent organic pollutants in mangrove food webs in Singapore [J]. Chemosphere, 2005, 61(3): 303–313.
[5] 林鵬, 陳榮華. 紅樹林有機碎屑在河口生態系統中的作用[J]. 生態學雜志, 1991, 10(2): 45–48+64.
[6] 邱霓, 徐頌軍, 邱彭華, 等. 珠海淇澳島紅樹林群落分布與景觀格局[J]. 林業科學, 2019, 55(1): 1–10.
[7] 李天宏, 趙智杰, 韓鵬. 深圳河河口紅樹林變化的多時相遙感分析[J]. 遙感學報, 2002, 6(5): 364–369+403.
[8] COHEN M L C, BEHLING H, LARA R J, et al. Impact of sea-level and climatic changes on the Amazon coastal wetlands during the late Holocene[J]. Vegetation History and Archaeobotany, 2009, 18(6): 425–439.
[9] 辛琨, 黃星. 海南東寨港紅樹林景觀變化與原因分析[J]. 濕地科學與管理, 2009, 5(2): 56–57.
[10] 吳培強, 張杰, 馬毅, 等. 近20a來我國紅樹林資源變化遙感監測與分析[J]. 海洋科學進展, 2013, 31(3): 406– 414.
[11] ANKITA M, MANI M R, VETHAMONY P. Assessment of the land use/land cover (LU/LC) and mangrove changes along the Mandovi-Zuari estuarine complex of Goa, India [J]. Arabian Journal of Geosciences, 2015, 8(1): 267–279.
[12] HU Luojia, LI Wenyu, XU Bing. Monitoring mangrove forest change in China from 1990 to 2015 using Landsat-derived spectral-temporal variability metrics[J]. International Journal of Applied Earth Observation and Geoinformation, 2018, 73(10): 88–98.
[13] 崔麗娟, 李偉, 張曼胤, 等. 福建洛陽江口紅樹林濕地景觀演變及驅動力分析[J]. 北京林業大學學報, 2010, 32(2): 106–112.
[14] 韓淑梅. 海南東寨港紅樹林景觀格局動態及其驅動力研究[D]. 北京: 北京林業大學, 2012.
[15] CHRISTOPHE P, PIERRE C, FRANCOIS F. Predicting and mapping mangrove biomass from canopy grain analysis using Fourier-based textural ordination of IKONOS images[J]. Remote Sensing of Environment, 2007, 109(3): 379–392.
[16] 林鵬. 中國紅樹林研究進展[J]. 廈門大學學報(自然科學版), 2001, 40(2): 592–603.
[17] 李皓宇, 彭逸生, 劉嘉健, 等. 粵東沿海紅樹林物種組成與群落特征[J]. 生態學報, 2016, 36(1): 252–260.
[18] 何斌源, 范航清, 王瑁, 等. 中國紅樹林濕地物種多樣性及其形成[J]. 生態學報, 2007, 27(11): 4859–4870.
[19] 韓維棟, 高秀梅, 盧昌義, 等. 雷州半島的紅樹林植物組成與群落生態[J]. 廣西植物, 2003, 23(2): 127–132+138.
[20] 韓維棟. 雷州半島紅樹林及其保護策略研究[D]. 廈門: 廈門大學, 2002.
[21] 楊娟, GAO J, 劉寶林, 等. 雷州半島紅樹林邊緣效應及其對海岸有機碳庫的影響[J]. 海洋學報(中文版), 2012, 34(5): 161–168.
[22] 李楠, 王友紹, 林立, 等. 湛江特呈島紅樹植物群落結構特征[J]. 生態科學, 2010, 29(1): 8–13.
[23] 孔凡洲, 于仁成, 徐子鈞, 等. 應用Excel軟件計算生物多樣性指數[J]. 海洋科學, 2012, 36(4): 57–62.
[24] 蘭國玉, 雷瑞德. 植物種群空間分布格局研究方法概述[J].西北林學院學報, 2003, 18(2): 17–21.
[25] 李學斌, 陳林, 李國旗, 等. 干旱半干旱地區圍欄封育對甘草群落特征及其分布格局的影響[J]. 生態學報, 2013, 33(13): 3995–4001.
[26] 張喬民. 我國熱帶生物海岸的現狀及生態系統的修復與重建[J]. 海洋與湖沼, 2001, 32 (4): 454–464.
[27] 林鵬,傅勤. 中國紅樹林環境生態及其經濟利用[M]. 北京: 高等教育出版社, 1995.
[28] 梁士楚. 廣西英羅灣紅樹植物群落的研究[J]. 植物生態學報, 1996, 20(4): 310–321.
[29] 韓路, 王海珍, 彭杰, 等. 塔里木河上游天然胡楊林種群空間分布格局與動態研究[J]. 西北植物學報, 2007, 27(8): 1668–1673.
[30] 陳鷺真, 王文卿, 張宜輝, 等. 2008年南方低溫對我國紅樹植物的破壞作用[J]. 植物生態學報, 2010, 34 (2): 186–194.
[31] 李玫, 廖寶文, 管偉, 等. 廣東省紅樹林寒害的調查[J]. 防護林科技, 2009, 26(2): 29–31.
[32] GILMAN E L, ELLISON J, DUKE N C, et al. Threats to mangroves from climate change and adaptation options: A review[J]. Aquatic Botany, 2008, 89(2): 237–250.
[33] AMIR A A. Mitigate risk for Malaysia’s mangroves [J]. Science, 2018, 359(6382): 1342–1343.
[34] LOVELOCK C E, CAHOON D R, FRIERSS D A, et al. The vulnerability of Indo-Pacific mangrove forests to sea-level rise [J]. Nature, 2015, 526(15538): 559–563.
[35] 王友紹主編. 紅樹林生態系統評價與修復技術[M] . 北京: 科學出版社, 2013.
[36] 廖寶文, 李玫, 陳玉軍, 等著. 中國紅樹林恢復與重建技術[M]. 北京: 科學出版社, 2010.
[37] CHENG Hao, WANG Youshao, FEI Jiao, et al. Differences in root aeration, iron plaque formation and waterlogging tolerance in six mangroves along a continues tidal gradient[J]. Ecotoxicology, 2015, 24(7-8): 1659–1667.
[38] PENG Yalan, WANG Youshao, FEI Jiao, et al. Ecophysiological differences between three mangrove seedlings () exposed to chilling stress[J]. Ecotoxicology, 2015, 24(7-8): 1722–1732.
[39] GUAN Guifang, WANG Youshao, CHENG Hao, et al. Physiological and biochemical response to drought stress in the leaves ofand[J]. Ecotoxi-cology, 2015, 24(7-8): 1668–1676.
Characteristics and distribution pattern of mangrove community in the Leizhou Peninsula
TANG Qiuxia1, 2, 3, 4, 5, WANG Youshao1, 2, 3,4, *
1. Key Laboratory of Tropical Marine Environmental Dynamics, South China Sea Institute of Oceanology, Chinese Academy of Sciences, Guangzhou 510301, China 2. Daya Bay Marine Biology Research Station, Chinese Academy of Sciences, Shenzhen 518121, China 3. Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou), Guangzhou 511458, China 4. Innovation Academy of South China Sea Ecology and Environmental Engineering, Chinese Academy of Sciences, Guangzhou 510301, China 5. University of Chinese Academy of science, Beijing 100039, China
Based on the mangrove community in the Leizhou Peninsula by survey and data analysis, it was also researched for mangrove species composition, community structure and distribution characteristics in the Leizhou Peninsula. The results showed that there were 24 species of mangrove communities in the Leizhou Peninsulathe, andcommunity was absolutely the first dominant species with an area contribution of 60.55%. A total of 13 species of true mangroves (accounting for 50% of the true mangrove species in China) and 10 species of semi-mangrove (accounting for 83.33% of the semi-mangrove species in China) were recorded in the survey areas. Shannon-Wiener index ranged from 0.09 to 1.08, and there were significant correlations in these indices with some research areas. The trend of Shannon index along the intertidal zone was smaller from high tide to low tide, which was relatively consistent.,contributing continuously on the beach from the landward to the seaward, was an important dominant species in the Leizhou Peninsula, whose aggregation at population level became higher from high tide to low tide. Compared with the results of the national "908" project for mangrove survey in 2008, the mangrove species recorded in the Leizhou Peninsula increased from 14 to 23, a large area ofthat died being the cold damage in 2008 was gradually recovered.(which introduced and artificial planting in Fucheng town in 1995) and the native true mangrove species (and), became the dominant species in this region, which meant the introduction ofhad some positive effect in restoring local mangrove communities. Although the species composition of the Leizhou Peninsula was rich, the structure of the mangrove community was still not rational. Improvement in residual secondary natural forests and plantations was the focus of the restoration and reconstruction of mangrove communities in Leizhou Peninsula. In the meantime, the planting of local mangrove species should be highlighted, which can protect mangrove species diversity and maintain the balance of mangrove ecosystems in the Leizhou Peninsula.
the Leizhou Peninsula; mangroves; species composition; spatial distribution; community structure
10.14108/j.cnki.1008-8873.2021.05.004
Q945.78
A
1008-8873(2021)05-023-10
2020-04-08;
2020-05-02
國家重點研發計劃(國科技基礎資源調查專項)(2017FY100700); 國家自然科學基金重點項目(U1901211和41876126); 中國科學院國際伙伴計劃項目(133244KYSB20180012); 中國科學院A類戰略性先導科技專項(XDA23050200和XDA19060201)
唐秋霞(1993—), 女, 廣東茂名人, 碩士研究生, 主要從事紅樹林生態學研究, E-mail:tangqiuxiala@163.com
通信作者:王友紹, 男, 二級教授, 主要從事海洋生態環境與生物資源研究, E-mail: yswang@scsio.ac.cn
唐秋霞, 王友紹. 雷州半島紅樹林群落特征及其分布格局[J]. 生態科學, 2021, 40(5): 23–32.
TANG Qiuxia, WANG Youshao. Characteristics and distribution pattern of mangrove community in the Leizhou Peninsula[J]. Ecological Science, 2021, 40(5): 23–32.
猜你喜歡
歌海(2024年6期)2024-03-18 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
兒童故事畫報·自然探秘(2022年3期)2022-04-27 00:17:10
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
瘋狂英語·初中天地(2018年6期)2018-11-24 02:39:24
汽車觀察(2018年10期)2018-11-06 07:05:26
歌海(2018年4期)2018-05-14 12:46:15
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
