鈣調素結合位點調控環核苷酸門控離子通道研究進展
2021-11-12 09:16:26余冰清嚴漢池
湖北農業科學 2021年20期
余冰清,嚴漢池
(天津大學生命科學學院,天津 300072)
1 CNGC 的發現及其結構研究
細胞信號傳導是一種可以響應并適應各種內部及外部因素的進化上的保守機制。環核苷酸門控離子通道(Cyclic nucleotide-gated ion channels,CNGC)是一類陽離子通道,它可以非選擇性地直接被環核苷酸(cAMP 和cGMP)這類細胞內信使小分子活化[1]。
對于CNGC 通道的研究最早在動物中進行。但是,與哺乳動物宿主中環狀核苷酸信號傳導的研究進展相反,這一主題在植物中的研究還較少。植物中的CNGC 不僅能夠調節花粉的正常發育并且能維持體內離子的穩態,另外,在植物對生物和非生物脅迫的應答反應中也起重要作用[2]。在植物中,1998年在大麥(Hordeum vulgare)中篩選鈣調素(Calmod?ulin,CaM)結合伴侶時偶然發現了第一個CNGC 基因。隨后,在其他植物中也鑒定出CNGC,例如擬南芥(Arabidopsis thaliana)、大 麥、水 稻[3](Oryza sati?va)、玉米(Zea mays)等。對CNGC 家族的研究表明,植物CNGC 基因可能在功能上以群依賴的方式區分。例如,可以依據系統進化關系將擬南芥CNGC家族分成5 個亞家族組(I、II、III、IVa 和IVb)。擬南芥CNGC 家族中包括20 個成員[4],目前對于植物環核苷酸門控離子通道的研究已經較為清晰地闡明其分子結構,并且發現它與Shaker 型鉀離子電壓門控離子通道的結構相似[5],其核心結構包括6 個跨膜α 螺旋(S1-S6),并且在S5 和S6 的中間有孔狀結構,即P loop,該區域是植物CNGC 區別于其他離子通道蛋白的重要標志,因為其影響離子的選擇性[6]。C末端具有環核苷酸結合區域(Cyclic Nucleotide mo?nophosphates Binding Domain,CNBD)和鈣調素結合區域(Calmodulin Binding Domain,CaMBD),CNBD由3 個α 螺旋(αA、αB、αC)和2 個β 折疊(β1 和β2)構成,β1 和β2 構成的片層結構位于αA 和αB 中間,CaMBD結合Ca2+后可以反饋調節CNGC的通道活性,當Ca2+濃度高時,Ca2+與CaM結合,CNGC通道關閉。
盡管有關CNGC 的報道較多,但由于大多數研究都集中在CNGC 的基因組表征、系統發育比較和潛在作用上,因此對其激活和功能機制的了解較少。動物的環核苷酸結合域位于肽鏈的C 端,鈣調蛋白結合域位于N 端;許多研究證明了植物的CaMBD 和CNBD 都位于C 末端[6](圖1)。序列比較分析結果顯示,植物的CaM 結合域和CNBD 結合域有部分重疊。Arazi 等[7]利用生物化學手段研究了煙草(Nicotiana tabacum)的CNGC 與模式植物擬南芥的CNGC,比較準確地詮釋了環核苷酸結合域(CNBD)與鈣調蛋白結合域(CaMBD)相重疊的論斷。也就是說,植物CNGC 與動物之間的主要區別在于植物CNGC 中包含了重疊的鈣調蛋白結合域(CaMBD)和環狀核苷酸結合域(CNBD),該結構上的區別表明,動植物的CNGC 之間調節蛋白質活性的機制和功能有所不同。然而近期有關多個鈣調蛋白結合位點正向和負向調節擬南芥CNGC12 的研究對于植物CNGC 調控模式產生了新的見解[8],為CNGC 調控新模型的建立提供了新的可能性。
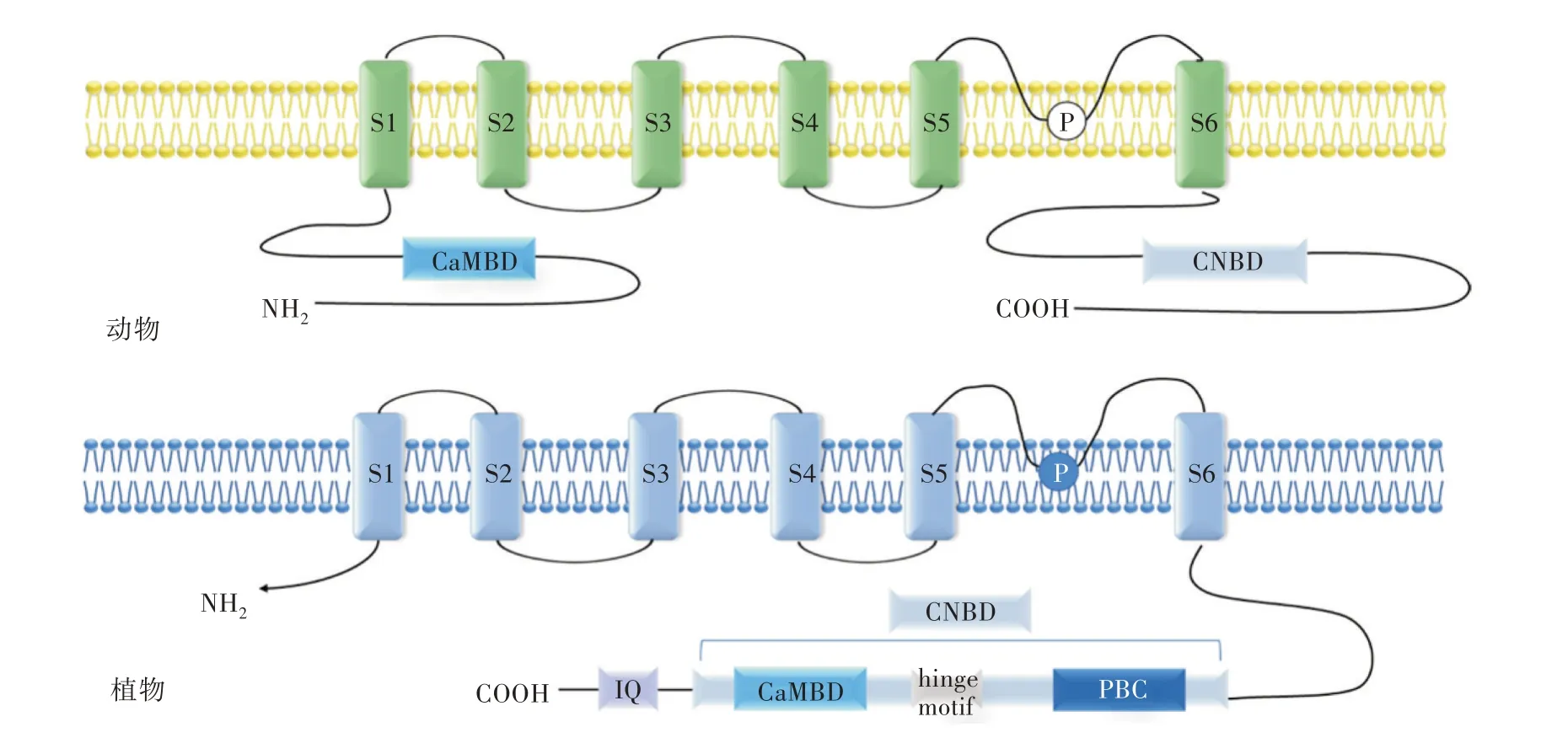
圖1 動植物環核苷酸門控離子通道(CNGC)的拓撲結構
2 CNGC 的調控因子——cNMP、Ca2+/CaM
CNGC 是一類陽離子通道,其特點在于具有不同程度的離子傳導選擇性,這些通道可以允許單價、二價陽離子的擴散,而CNGC 的活性受兩個方面的調控,一方面受cGMP、cAMP 的調節,另一方面受CaM 與CNBD 的可逆結合調節,結合后蛋白的變構導致通道的開放和閉合。
2.1 cNMP
環狀核苷酸(cNMP)是動植物體內非常重要的第二信使。cNMP 在調控離子動態變化中起重要作用,環核苷酸通過與環核苷酸結合結構域(CNBD)結合來調節其開放。現有的證據已經表明,環狀核苷酸門控通道是植物細胞中主要的cNMP 效應子。這些通道是重要的細胞開關,可將細胞內環狀核苷酸濃度的變化轉化為膜電位和離子濃度的變化。環狀核苷酸(cNMP),如3',5'-環腺苷酸(cAMP)和3',5'-環鳥苷酸(cGMP)與所有活生物體中的信號轉導相關[9]。
2.2 Ca2+/CaM
鈣調素(CaM)是細胞內Ca2+信號傳導途徑中的主要信號轉導分子,它作為一種重要的鈣受體蛋白,可以通過與Ca2+結合進而調節細胞內一些鈣調結合蛋白(CaMBPs)的活性,介導調節通過Ca2+濃度變化引發的各種生理生化反應。
在 調 控 方 面,Ca2+/CaM 結 合CNGCs 是CNGC 家族首次被鑒定出來的生物化學功能。目前的研究顯示,Ca2+/CaM 病原菌入侵細胞后會引起胞質內Ca2+含量增多,一方面激活下游Ca2+依賴型的生理生化反應,進而調控細胞一系列的相關生理過程;另一方面,被激活的CaM 能夠反饋抑制鈣信號。CNGC 之所以能夠被胞質內增多的Ca2+/CaM 抑制,可能是由于Ca2+/CaM 阻礙了環核苷酸激活CNGC,因為該信號級聯通過病原菌響應。電生理學試驗顯示,在施加CaM 拮抗劑后,持續的胞外刺激會導致Ca2+不斷內流[10]。用CaM 拮抗劑處理植物細胞,在病原菌刺激下Ca2+含量依然會升高,也就是說Ca2+/CaM 對于CNGC 的反饋抑制可能阻礙了Ca2+的持續內流,因此,其對于維持胞內Ca2+含量穩定具有重要作用[11]。
雖然,研究者已經了解到CaM/Ca2+可以調控其活性,但是,與動物相比,對其活性調控的具體機制還不了解。已有的鈣調蛋白與擬南芥環核苷酸門控離子通道的結合的研究顯示,CaM 與其動物中的對應部分相似,對植物的CNGC 具有負調控作用。另一方面,CaM 結合親和力的改變會導致CNGC 調控情況的復雜變化。
2.3 動植物CNGC 活性受cNMP 和CaM 調控
動物CNGC 主要分布于化學感受器和光感受器中,在轉換膜外信號中起作用。在嗅覺和視覺信號轉導過程中,CNGC 通道通過cAMP 或cGMP 調節通道活性,引起Na+流入,從而介導嗅覺神經元的嗅覺轉換以及桿狀細胞中光感受器的光轉換[12]。而植物CNGC 被激活的方式有2 種,除了被直接激活外,也可以通過與環核苷酸(cAMP/cGMP)可逆性結合的方式被激活,但其門控作用主要依賴于cAMP 和cGMP 的存在[13],同時受Ca2+調控。環核苷酸信號的產生和消退取決于腺嘌呤環化酶和鳥嘌呤環化酶,以及可以將環狀核苷酸水解成ATP 和GTP 的磷酸二酯酶[14]。當環狀核苷酸與通道蛋白結合時,它會誘導蛋白構象的變化,導致P 區域的旋轉,然后通道大門打開,從而促使陽離子跨膜移動[15]。
如上所述,CaMBD 位于與CNBD 的C 末端α-螺旋重疊的部位。CaMBD 具有4 個氨基酸殘基,為Trp-Arg-Thr-Trp(WRTW),是CaM 結合所必需的。CaMBD 在CNBD 內的位置允許CaM 與cNMP 作為通道電導的變構門控中的配體競爭。最近,在模式植物AtCNGC20 中發現通過獨特的異亮氨酸-谷氨酰胺(IQ)CaM 結合基序與CaM 結合,該基序與CNBD中的α-螺旋相鄰但不重疊[8]。一些番茄和擬南芥CNGC 家族的域組織分析試驗也證明了一些成員是含有IQ CaM 結合基序的,該結果首次讓人們意識到植物CNGC 可能含有2 個替代的CaM 結合基序(CaMBD 和IQ CaM 結合基序)。除此之外,分析結果還顯示,它類似于動植物中的電壓依賴性Ca2+通道,表明通過CaM 對植物CNGC 的配體調控可能比以往預期的更加復雜,因為上述2 個結構域(CaMBD和IQ)具有不同的功能,具體調控模式還需進一步研究分析。
3 CaM 對CNGC 調節的復雜模式
迄今為止研究的所有植物和動物CNGC 具有至少1 個鈣調蛋白(CaM)結合結構域。研究表明,動植物CNGC 的N 端和C 端都延伸至細胞質中,除了CaMBD 的位置不同外,其他基本相同,其中CNBD在多肽的C 末端,由3 個α 螺旋(αA、αB、αC)和2 個β 折疊(β1 和β2)構成,由β1 和β2 組成的片層結構位于αA 和αB 之間。動物CNGC 的CaMBD 位于肽鏈的N 末端的上游,但植物中CaMBD 位置與動物不同,它位于多肽的C 末端,即CNBD 截短的αC 螺旋的起點處。因此,由于植物CNBD 是重疊的結合域,它既參與鈣調蛋白的結合,又參與環核苷酸的結合。
3.1 動物CNGC 的變構調節模式
首先,對于動物中cNMP 對CNGC 的調節,響應激活胞內嘌呤核苷酸環化酶(NCs)的細胞外信號合成cNMP,它們分別由腺苷三磷酸腺苷(ATP)和鳥苷三磷酸鳥苷(GTP)催化cAMP 和cGMP 合成。細胞內cNMP 的濃度受磷酸二酯酶(PDEs)調節,磷酸二酯酶進而將cAMP 和cGMP 代謝為無活性的核苷酸單磷酸[16]。cNMP 結合并激活環狀核苷酸結合蛋白(CNBP),包括依賴于cAMP 和cGMP 的蛋白激酶(PKA 和PKG),這被認為是動物中環狀核苷酸信號傳導的主要作用因子。通過環狀核苷酸來激活在動物各種生理過程中起基本作用的通道,其中包括環狀核苷酸門控通道(CNGC)[17]。具體調控過程如下:氣味分子與化學感受器中的G 蛋白耦聯型受體可以通過結合進而激活腺苷酸環化酶,產生cAMP,打開cAMP 門控陽離子通道(cAMP-gated cation channel),并引起Na+內流,膜去極化,從而產生神經沖動,最終形成嗅覺或味覺。
哺乳動物CNGC 變構調節的機制被充分表征,其中環核苷酸單磷酸酯cAMP 和cGMP 與C 端CNBD的結合提供了通道傳導的門控機制[18],而哺乳動物CNGC 功能的變構抑制又由CaM 結合調節,這降低了通道對cAMP/cGMP 的親和力[19]。哺乳動物CNGC具有多種CaM 結合位點,目前在6 種不同CNGC 同種型的N 末端及C 末端均已發現至少9 種CaMBD,然而,到目前為止,試驗只提供了N 端CaMBD 的功能作用的證據。通常,CaM 是在升高的細胞內Ca2+水平下,在CNGCs 的反饋調節中通過結合到1 個或多個CaMBD,從而變構性抑制CNGC 電導[20]。
3.2 鈣調蛋白結合位點調控植物CNGC 的復雜模式
植物中的CNGC 是信號轉導級聯反應(Signal transduction cascades)的組成部分。通過這種級聯反應,它們將細胞外部信號轉換成可以穿過細胞膜并作用于細胞的陽離子流[12]。如前文所述,植物CNGC 能夠直接被激活,還受cAMP/cGMP 以及Ca2+調控。
cAMP/cGMP 分別通過跨膜(tmAC)或可溶性(sAC)腺苷酸環化酶/跨膜(tmGC)或可溶性(sGC)胍基環化酶合成。它們都被磷酸二酯酶(PDE)水解成無活性的AMP 和GMP。根據現有技術的實際研究發現,CNGC 是植物細胞中cNMP 的主要靶標。環狀核苷酸可以直接與環狀核苷酸結合域(CNBD)結合,以誘導蛋白的構像變化,調節CNGC 孔的打開或關閉,并促進陽離子跨膜移動。
先前的研究假設CaMBD 允許CaM 作為配體與環狀核苷酸單磷酸在通道電導的變構門控中競爭結合CNBD[21]。首次提出該區域時,它被視為植物CNGC 的惟一CaMBD,就是說,由于植物CNBD 是1個重疊的結合域,CaM 與植物CNGC 的結合會干擾CNGC 與環狀核苷酸的結合,競爭性的結合進而影響通道的激活[7],表明由CaM 調節的動植物CNGC之間具有不同的調節機制。CaM、環核苷酸和Ca2+不僅各自都能與CNGC 相互作用,而且可以通過整合的方式控制離子通過植物離子通道。當環核苷酸與CNGC 結合而被激活時,通道打開導致胞外Ca2+內流。一方面,胞內Ca2+的增加將導致Ca2+依賴蛋白激酶(CDPKs)的激活,進而導致下游靶蛋白的磷酸化;另一方面,由Ca2+激活的鈣調蛋白將會與環核苷酸競爭性地結合CNGC,這就一定程度上阻止了環核苷酸與CNGC 結合,通道活性受到抑制,從而限制了Ca2+的進一步內流。
此前的研究均假設植物的CNGC 受Ca2+傳感器鈣調蛋白(CaM)負調控,并且先前的工作也都只集中在與植物CNGC 的CNBD 重疊的C 末端的CaMBD上。而近幾年研究發現,模式植物AtCNGC20 通過與αC-螺旋相鄰卻并不重疊的獨特異亮氨酸-谷氨酰胺(IQ)基序與CaM 結合[22],表明植物CNGC,如哺乳動物CNGC,可具有多種CaMBD。
3.3 動植物CNGC 在鈣離子變構調節上的異同
為了進一步了解CNGCs 的調節和生理作用,DeFalco 等[8]對模式植物擬南芥CNGC12 的CaM 結合特性和功能進行了全面的表征。擬南芥CNGC12的αC-螺旋區域具有低保守性,因此研究者認為其無法結合CaM,然而,該研究證明這種同種型在其胞質N 和C 末端都具有新的CaMBD,提供了單個植物CNGC 同種型具有多個CaMBD 的證據。表明擬南芥CNGC12 在其胞質N 和C 末端含有多個CaMBD,包括NT、CT 和IQ(圖2)。在CNBD 之外的CNGC12的C 末端存在新的CaMBD(CT),該區域以前并未在任何植物CNGC 中表征過。先前已報道植物CNGC同種型中沒有N 末端CaMBD,然而在N 末端區域的覆蓋測定中也觀察到明顯的CaM 結合(NT)。由于其試驗確定了CaM 與IQ 基序的結合是通道功能所必需的,不僅表明了CaM 可以正向和負向調節CNGC12,同時說明了CaM 對植物CNGC 調節模式較為復雜,它與先前提出的競爭性配體模型相反,這表明有可能植物和動物CNGC 之間存在相似性。
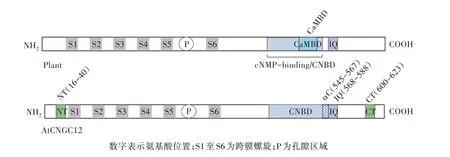
圖2 模式植物擬南芥CNGC12 的CaMBD 位置(包括NT 和CT 位點的描述)
鑒于對其CaMBD 的生化和生理解剖,DeFalco等[8]提出了CaM 調節CNGC12 的模型。該模型假定CNGC12 通過IQ 基序以Ca2+獨立的方式進行組成性調節,而Ca2+/CaM 可以順序結合多個CaMBD。該模型顯示CNGC12 直接介導Ca2+的內流,但是不能排除CNGC12 信號觸發其他通道介導的Ca2+電流的可能性。該模型包含了一些推測性的特征,這些特征與目前的研究結果一致,但仍需要進一步研究驗證,其中一些特征與各種動物通道有明顯的相似性,例如CaM 介導的亞基間和亞基間相互作用的橋接以及CaM 在多個CaMBD 之間的動態運動[23],類似的CaM 調節特征廣泛分布在各種動物電壓或Ca2+門控的陽離子通道中,例如很多脊椎動物的陽離子通道中就存在類似機制[24]。
結構性、功能性、生理和電生理學研究的結合將有助于闡明Ca2+對CaM 介導的CNGC 調控的復雜模式。考慮到CNGC 和CaM 在免疫中的重要作用[25],進一步表征CaM-CNGC 相互作用對于理解免疫應答過程中CNGC 介導的Ca2+信號傳導至關重要。
4 展望
環核苷酸門控通道作為植物細胞中環核苷酸的靶點,越來越受到人們的關注。尤其是近年來細胞斑塊鉗位、植物基因組測序以及基因組學方法的進一步發展,促進了CNGC 編碼基因的識別和調控等方面研究的進展,為研究人員闡明通道的結構與功能間的聯系提供了有效方法和技術支撐。信號轉導通路缺陷的突變體的存在,使人們對CNGC 生理功能的認識取得了迅速進展。
幾乎所有的植物信號過程都有Ca2+的參與,然而至今為止,植物中只有為數不多的鈣離子通道家族被發現。另外,植物中的CNGC、GLR 等鈣通道最早是通過動物中的同源蛋白進行序列相似性比對而挖掘出來的。在對CNGC 進行研究時,如果能考慮動植物間的相似點和區別,將對闡明植物CNGCs 功能及其調控機理有很大的幫助。所以探究CNGCs相應的配體、門控分子和活性調節機制是今后研究的一個重要目標。
總而言之,要完全理解CNGC 的功能,還需要采取多種研究手段。結合更多的結構功能、生理和電生理研究將有助于闡明Ca2+/CaM 介導的CNGC 調控的復雜模式。將鈣信號整合到植物中更大的信號網絡中的研究應該得到更多的關注。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
