二種土甲族幼蟲的分子鑒定
2021-11-11 06:44:02張建英丁榮榮
農業科學研究 2021年3期
張建英,李 彪,賈 龍,丁榮榮
(1.寧夏大學 生命科學學院,寧夏 銀川 750021;2.寧夏大學 農學院,寧夏 銀川 750021)
土甲族(Opatrini)隸屬鞘翅目(Coleoptera)擬步甲科(Tenebrionidae)擬步甲亞科(Tenebrioninae)[1],在寧夏分布有10屬35種[2]。土甲族昆蟲是我國荒漠半荒漠地區主要的農、林、牧業害蟲和倉儲害蟲[3]。四五月份成蟲交配產卵,孵化成幼蟲[4],幼蟲一般棲息在土壤之中,口器較發達,食性雜[2],為害植物嫩芽、根部和種子,損害植物生長,甚至致其死亡。土甲族昆蟲的幼蟲期是取食量最大、對農林牧業為害最嚴重的發育階段。因此,高效、準確地對該類昆蟲幼蟲進行種的鑒定已成為該類昆蟲防治與利用的基礎,也是該類群昆蟲防治與利用研究的核心內容之一。雖然目前對于土甲族昆蟲的研究很多,在成蟲分類研究方面較為完善,但對于土甲族昆蟲幼期種類鑒定的研究卻鮮有報道,其主要原因是該族昆蟲幼期標本不易獲得,并且屬間或種間差異小,給種類鑒定造成了很大困難。
目前,國際上對昆蟲的鑒定大多采用形態學方法,形態學鑒定是將成蟲的形態學特征作為依據進行鑒定,而幼期的特征差異小,僅從形態特征入手,鑒定困難,必須等到幼蟲老熟時,即形態特征骨化穩定后才能鑒定。因此幼蟲鑒定一直是分類學的一個難題,采取的辦法往往是將幼蟲在實驗室內進行人工飼養,待其發育成成蟲后再進行鑒定,費時費力,嚴重影響物種鑒定的速度。20世紀80年代以來,隨著分子生物學技術和生物信息學的發展以及PCR技術的建立與完善,一系列分子生物學技術被廣泛應用于生物鑒定,極大地彌補了傳統形態學鑒定方法存在的缺陷,DNA條形碼技術就是其中具有代表性的一種[5]。加拿大動物學家Hebert等[6]首先倡導將條形編碼技術應用于生物物種鑒定。DNA在生物體生長發育過程中不會改變,以DNA序列作為檢測對象,不受個體發育過程、不同發育形態和生長環境等的影響,且DNA條形碼技術具有可操作性強等特點,目前已廣泛應用于物種鑒定[7]。
土甲族昆蟲生活史普遍在1年左右。野外獲得低齡幼蟲后,因其鑒別特征尚未硬化定型,專業人員很難做出準確鑒定,往往通過實驗室飼養,待其發育成老熟幼蟲或成蟲后再進行鑒定,而這個過程短則數月,長則1年,鑒定周期很長。為此,本研究利用DNA條形碼技術以線粒體COⅠ基因、線粒體16S基因為分子標簽,對土甲族偽堅土甲屬粗背偽堅土甲(Scleropatrum horridum horridum)和笨土甲屬阿笨土甲(Penthicus alashanica)成蟲、低齡幼蟲和高齡幼蟲等8個樣品的線粒體COⅠ基因、線粒體16S基因序列進行比對分析,若同種土甲成蟲(雌性和雄性)和幼蟲(低齡和高齡)之間的16S和COⅠ基因序列相似度達到98%及以上,說明可以通過DNA條形碼技術擴增未知幼蟲的16S和COⅠ基因序列,然后與已知成蟲的16S和COⅠ基因序列進行比對,根據相似度來判斷未知幼蟲為哪種已知成蟲的幼蟲,從而鑒定未知幼蟲種類。Hajibabaei等[8]對鱗翅目隸屬3科521種的DNA條形碼進行分析鑒定,證明DNA條形碼的準確性極高;Virgilio等[9]對鞘翅目、雙翅目、半翅目、膜翅目、鱗翅目和直翅目進行了DNA條形碼技術鑒定的效率分析,認為在無數據庫參照時其物種鑒定的效率會受影響;岳巧云等[10]應用COⅠ基因鑒定形態近似的鞘翅目幼蟲,結果準確、可靠。該研究的意義在于:昆蟲幼蟲分類研究中采用DNA條形碼技術,相比傳統的實驗室內飼養幼蟲待其鑒別特征骨化后再進行鑒定的傳統幼蟲鑒定方法,可以有效減少鑒定時間和成本,鑒定結果也比較準確、客觀、簡便。采用線粒體16S基因和線粒體COⅠ基因同時作為DNA條形編碼的標記基因,突破了單分子標記的局限性。
1 材料與方法
1.1 實驗材料
1.1.1 供試蟲源 本研究用于檢測的粗背偽堅土甲和阿笨土甲成蟲采集于寧夏賀蘭山國家級自然保護區;幼蟲經人工室內飼養獲得。
1.1.2 試劑 Taq Plus DNA聚合酶、10×PCR Buffer(含Mg2+)、dNTP(10 mmol/L)、滅菌去離子水、引物DNA、6×DNA Loading Dye、DNA Ladder Mix(100~10 000 bp)、DNA Ladder Mix (100~3 000 bp)、50×TAE、瓊脂糖H、SanPrep柱式DNA膠回收試劑盒、無水乙醇、異丙醇、1×TE、BigDye Terminator v1.1、POP-7TMPolymer、HiDi Formamide、EDTA、NaAc、NaOH、Ezup柱式動物組織基因組DNA抽提試劑盒
1.1.3 儀器 冰箱、潔凈工作臺、PCR儀、凝膠成像儀、測序儀、冷凍離心機、臺式高速離心機、電泳儀、電泳槽、微型旋渦混合儀、數顯恒溫水浴鍋、移液器、紫外可見分光光度計等。
1.2 實驗方法
1.2.1 室內人工飼養方法 室內飼養成蟲時,用較大的容器盛沙土,將雌雄蟲按一定比例放置其中進行飼養。沙土必須進行100~120℃高溫處理24 h,目的是殺死沙土中的螨類。沙土用水拌濕,土壤含水量在14%~18%為宜。將拌好的沙土裝入容器,上面留出1/3體積的空余,在濕土上覆蓋薄薄一層干沙土。以燕麥片、黃瓜、麩皮、苜蓿等作為主要飼料。當卵達到一定數量時,用軟毛筆將卵轉移到孵卵容器中,孵化卵的沙土濕度不能過大。當卵孵化成幼蟲后,將其輕輕轉移到新容器中飼養。飼養幼期土甲較飼養成蟲困難,幼蟲和蛹對沙土水分有一定要求,容器內沙土水分不宜過大,否則會使幼蟲和蛹發黑腐爛;但也不能沒有水分,當沙土水分減少后,可通過吸水紙向其中滴水以保持一定濕度。預蛹期時老熟幼蟲會在容器底部打造蛹室,在此期間不能搖動容器。飼養過程中若發現寄生螨需及時處理,可在漏斗中用干燥的沙土來回沖刷蟲體,從而去除體表的寄生螨;寄生菌需隔離處理。除預蛹期和蛹期外,需定期更換沙土從而達到保濕、除菌的目的。用透明容器飼養需注意避光。
1.2.2 DNA提取 采用Ezup柱式動物組織基因組DNA抽提試劑盒分別提取粗背偽堅土甲雌性成蟲、雄性成蟲、低齡幼蟲、高齡幼蟲和阿笨土甲雌性成蟲、雄性成蟲、低齡幼蟲和高齡幼蟲8個樣品的DNA。
1.2.3 PCR擴增條件及產物檢測 選用線粒體COⅠ基因、線粒體16S基因的部分片段作為分子標記,委托生工生物工程(上海)股份有限公司設計合成引物進行PCR擴增(表2)。

表2 引物序列
PCR反應體系包括模板DNA1μL、上下游引物(10μmol/L)各1μL、Dntp(mix)(10 mmol/L)1μL、Taq Buffer(10×)2.5μL、Taq酶(5 U/μL)0.2μL、dd H2O加至25μL。擴增條件:第1步95℃預變性5 min;第2步94℃變性30 s,63℃(每循環降0.5℃)退火30 s,72℃延伸30 s共10個循環;第3步95℃變性30 s,58℃退火30 s,72℃延伸30 s共30個循環;第4步72℃修復延伸10 min;第5步4℃保溫。
產物檢測:取5μL PCR擴增產物,以標準DNA Marker為參照,在1%瓊脂糖凝膠上進行電泳分離,150 V、100 mA條件下電泳10~20 min,在紫外檢測儀上觀察拍照。按純化試劑盒操作說明將目的片段電泳條帶單一且明亮的PCR產物二次純化,用測序儀對PCR產物進行雙向測序。
1.2.4 序列分析 本研究測定了土甲族2種昆蟲的16S基因和COⅠ基因部分序列,用Chromas軟件觀察每對序列峰值圖,結合Contig Express軟件進行人工校閱得到所測序列。用分子進化遺傳分析軟件MEGA 7分別統計2個基因的序列組成,計算堿基組成,統計轉換(transition,TS)和顛換(transversion,TV)數,計算轉化/顛換比率,分析2種昆蟲間的DNA序列差異。將測得的DNA序列輸入NCBI(https://www.ncbi.nlm.nih.gov/)網站,進行Blast核苷酸序列的同源性分析,用Clustal X 1.83軟件對2種基因14條序列進行多重比對。
2 結果與分析
2.1 DNA提取
通過分光光度計對測得的各樣品DNA質量進行檢測,得到基因組DNA溶液OD值為1.7~2.0。DNA提取檢測結果顯示除6號樣外,其他7個樣品均得到清晰明亮且單一的目的條帶,滿足后續PCR擴增要求。其中6號樣沒有得到目的條帶,結果見圖1,這可能因為在樣品處理過程中6號樣基因組DNA發生降解,從而導致未能成功提取出6號樣基因組DNA。
圖1 DNA提取檢測圖
2.2 PCR擴增
對除6號樣外的7個樣品進行16S和COⅠ基因PCR擴增,得到的電泳圖如圖2、圖3所示。對于16S基因片段,7個樣品均得到清晰明亮的目的條帶;對于COⅠ基因片段,7個樣品亦均得到清晰明亮的目的條帶,2個基因片段結果均為單一條帶,可用于后續的基因測序分析。本實驗采用特異性引物進行PCR擴增,擴增結果表明:PCR擴增產生大小相同的1條帶,無其他條帶干擾(圖2、圖3),因此所得到的序列應該是線粒體序列,而非核中線粒體假基因。
圖2 16S基因PCR產物電泳圖
圖3 COⅠ基因PCR產物電泳圖
2.3 基因序列組成及變異
2.3.1 16S基因序列組成及變異 在2種土甲的16S基因序列中,有62個保守位點,935個變異位點,變異率為93.4%,645個簡約信息位點,285個自裔位點,結果表明種間序列變異較大。堿基T、C、A、G的平均含量分別為32.0%、15.3%、45.1%和7.6%。A+T的平均含量為77.1%,明顯高于G+C含量(22.9%)。堿基T、C、A、G在每個氨基酸密碼子的不同位點差異不大,第1位點A+T含量(78.0%)高于G+C含量(22.0%),第2位點A+T含量(76.7%)高于G+C含量(23.3%),第3位點A+T含量(76.4%)高于G+C含量(23.6%),16S基因在各類昆蟲中富含A、T,如鱗翅目A+T的含量為78.4%,蜉蝣目A+T的含量為66.3%[11],本實驗結果與之相似,另外所有擬步甲科昆蟲的序列總和中A、T堿基所占總堿基的百分比均高于G、C堿基所占總堿基的百分比,由此可知所有序列都表現出明顯的AT偏向性(表3),這表明所有序列都符合昆蟲線粒體的堿基組成結構。堿基替換是DNA序列堿基進化的重要衡量指標,有堿基轉換和堿基替換兩種形式[12]。從堿基替換的結果看,TV的發生遠遠多于TS,全組數據TS/TV的平均值為0.3,在第1、2、3位點的比值分別為0.3、0.4、0.4。其中轉換主要發生在C←→T之間,顛換主要發生在A←→T和A←→C之間,其他類型的替換相比上述幾種替換發生較少(表4)。
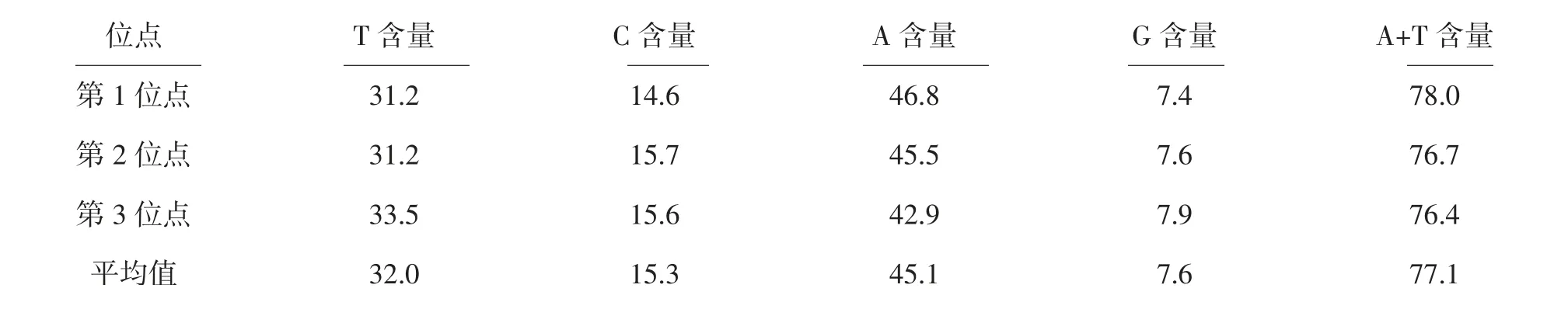
表3 土甲族2種昆蟲的16S基因序列組成 %
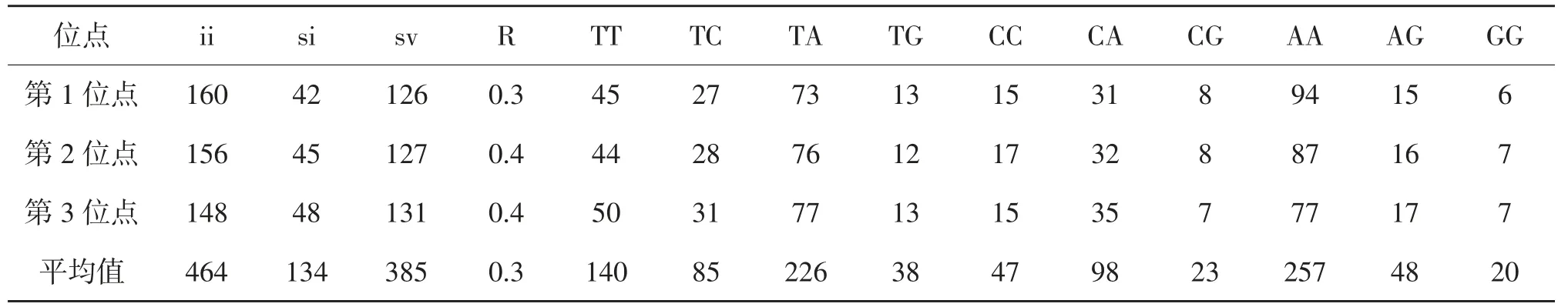
表4 土甲族2種昆蟲的16S基因堿基替換統計
2.3.2 COⅠ基因序列組成及變異 在2種土甲的COⅠ基因序列中,有15個保守位點,1 021個變異位點,變異率為98.2%,698個簡約信息位點,321個自裔位點,結果表明種間序列變異較大。堿基T、C、A、G的平均含量分別為35.0%、17.5%、30.8%和16.7%。A+T的平均含量為65.8%,明顯高于G+C含量(34.2%)。第1位點的A+T含量為65.3%,第2位點的A+T含量為62.4%,第3位點的A+T含量為69.7%。第3位點表現出比較強的AT含量偏向性,A+T含量較前兩個位點高,該位點堿基G的含量最少為12.8%。所有擬步甲科昆蟲的序列總和中A、T堿基所占總堿基的百分比為63.1%[13],與本實驗結果相符,都表現出明顯的AT偏向性(表5)。這表明所有序列符合昆蟲線粒體堿基組成結構。從堿基替換的結果看,TV的發生略多于TS,TS/TV的平均值為0.5,在第1、2、3位點的比值分別為0.5、0.5、0.5。其中轉換主要發生在C←→T之間,顛換主要發生在A←→T之間,其他類型的替換相比上述幾種替換發生較少(表6)。
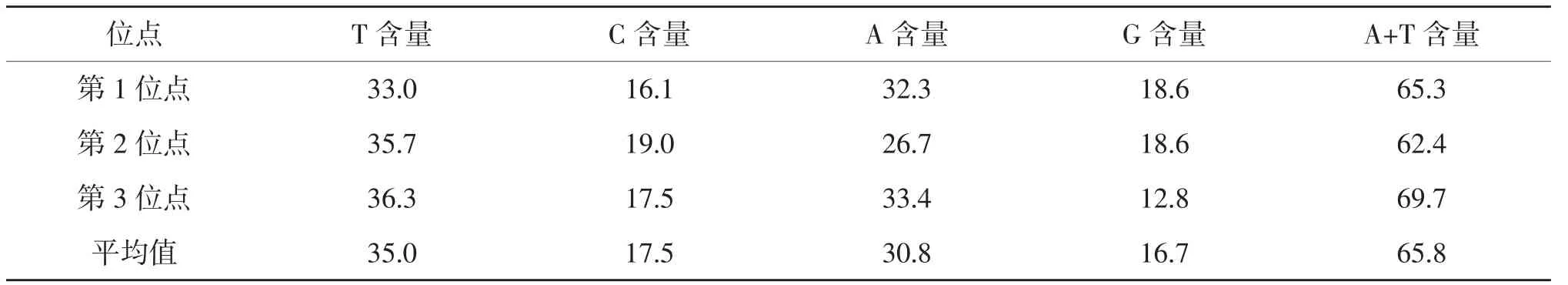
表5 土甲族2種昆蟲的COⅠ基因序列組成 %

表6 土甲族2種昆蟲的COⅠ基因堿基替換統計
2.4 同源性相似度比對分析
2.4.1 16S基因同源相似度 由表7和表8可知,粗背偽堅土甲雄性成蟲與低齡幼蟲及高齡幼蟲的相似度達到100%,與雌性成蟲相似度達100%,雌性成蟲與低齡幼蟲及高齡幼蟲相似度達到100%,低齡幼蟲與高齡幼蟲相似度也達到100%。阿笨土甲雌性成蟲與低齡幼蟲相似度達99.11%,與高齡幼蟲相似度達98.97%,低齡幼蟲與高齡幼蟲相似度達99.56%。2種土甲種間16S基因相似度:成蟲之間最高為93.72%,幼蟲之間最高為93.33%,成蟲與幼蟲之間最高為93.55%。2種土甲種內雌性成蟲的16S基因與低齡幼蟲和高齡幼蟲的同源相似度均達到98%以上,粗背偽堅土甲雄性成蟲的16S基因與低齡幼蟲和高齡幼蟲的同源相似度均達到98%以上。說明16S基因在不同種的各個蟲態之間差異較大,在同一種內不論是雌性還是雄性成蟲,它們與不同齡期幼蟲都有極高的16S基因同源相似度。

表7 種內16S基因BLAST比對結果
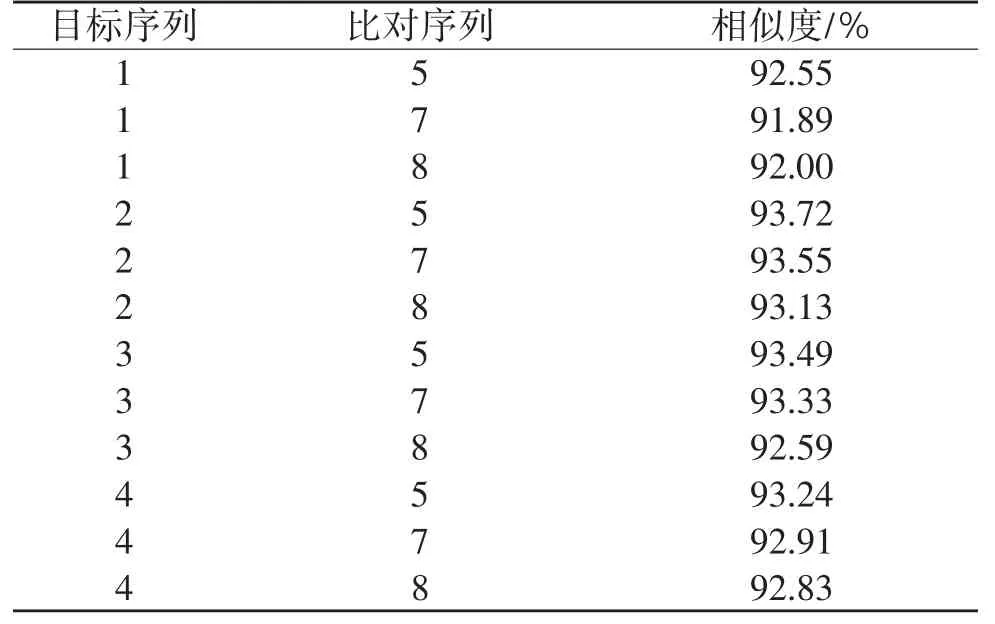
表8 種間16S基因BLAST比對結果
2.4.2 CO I基因同源相似度 由表9和表10可知,粗背偽堅土甲雌性成蟲與低齡幼蟲相似度達98.84%,與高齡幼蟲相似度達99.50%,與雄性成蟲相似度達98.49%;雄性成蟲與低齡幼蟲相似度達98.94%,與高齡幼蟲相似度達99.39%,低齡幼蟲與高齡幼蟲相似度達99.24%。阿笨土甲雌性成蟲與低齡幼蟲相似度達98.63%,與高齡幼蟲相似度達99.24%,低齡幼蟲與高齡幼蟲相似度達98.79%。2種土甲種間COⅠ基因相似度:成蟲之間最高為91.57%,幼蟲之間最高為90.53%,成蟲與幼蟲之間最高為91.75%。2種土甲雌性成蟲的COⅠ基因與低齡幼蟲和高齡幼蟲的同源相似度均達到98%以上,粗背偽堅土甲雄性成蟲的COⅠ基因與低齡幼蟲和高齡幼蟲的同源相似度均達到98%以上。說明COⅠ基因在不同種的各個蟲態間差異較大,在同一種內不論是雌性還是雄性成蟲,它們與不同齡期幼蟲都有極高的COⅠ基因同源相似度。
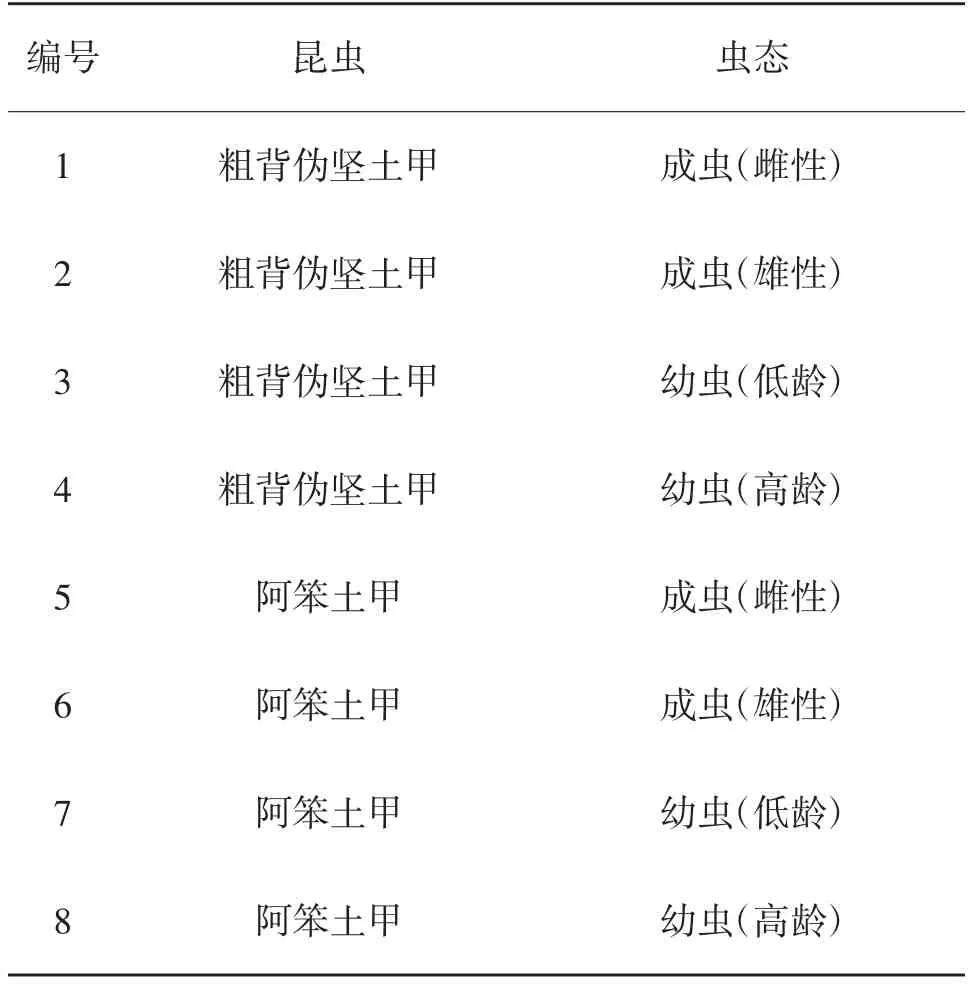
表1 樣品編號及昆蟲、蟲態對應關系
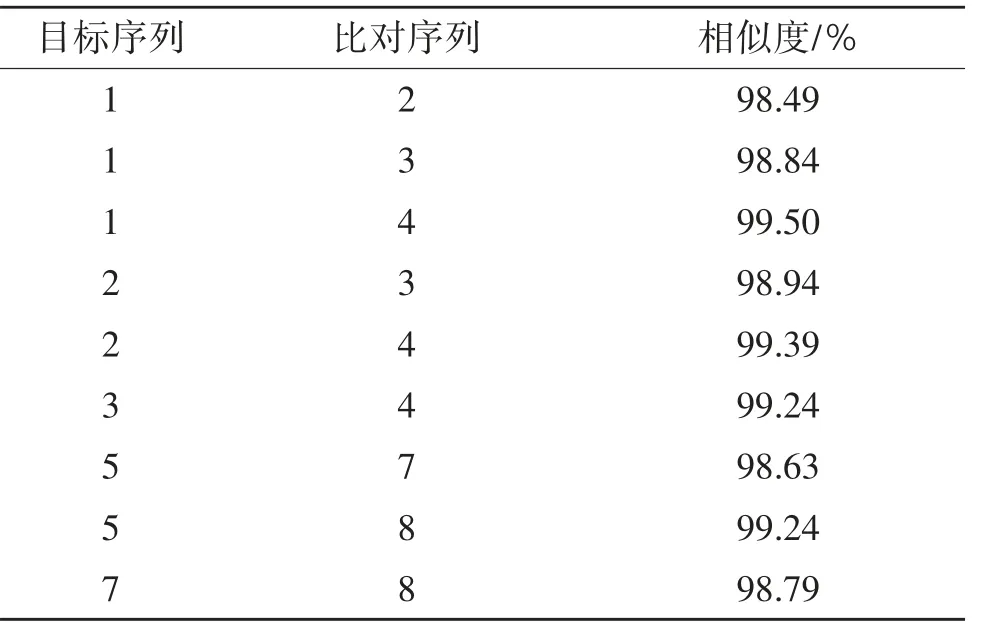
表9 種內COⅠ基因BLAST比對結果
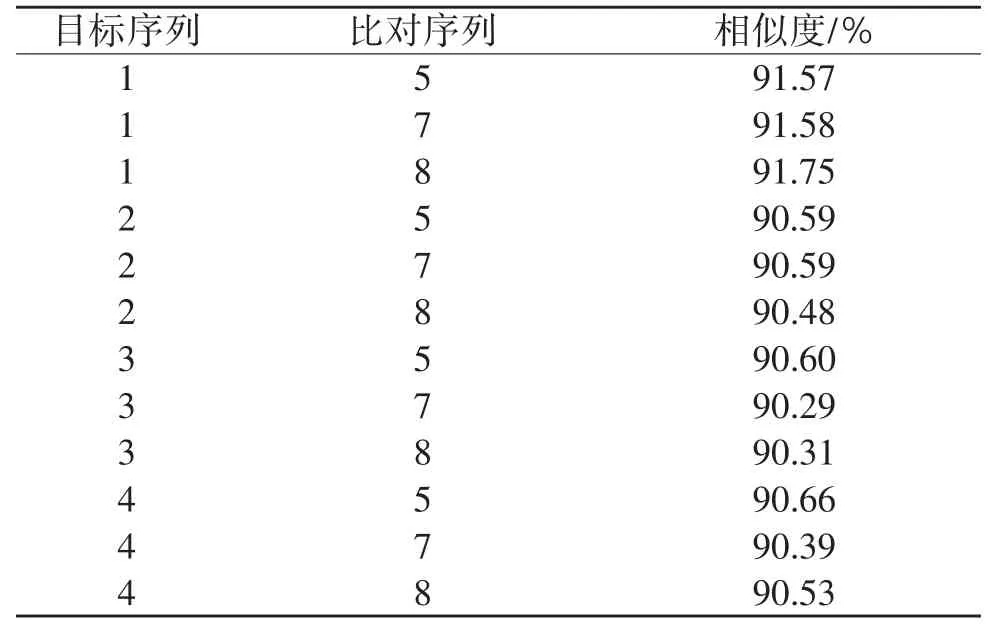
表10 種間COⅠ基因BLAST比對結果
3 結論與討論
目前DNA條形碼技術主要被用于鑒定近緣種或者建立數據庫分析某個大的類群[14-15]。在昆蟲研究工作中,非昆蟲分類學專家很難通過形態特征對一些昆蟲進行鑒定,而分子鑒定可以有效解決這一問題,DNA條形碼技術就是其中的代表之一,對昆蟲特定基因片段進行測序及比對,確定其分類學地位。
運用DNA條形碼技術鑒定物種有一個關鍵點就是基因的選擇,基因的選擇直接關系到鑒定結果的準確性。研究表明,許多生物包括昆蟲類群存在核中線粒體基因的拷貝,即線粒體假基因Numts(nuclear mitochondria DNA segments)[16]。隨著線粒體假基因研究的推進,人們開始懷疑線粒體基因作為系統發育研究分子標記的準確性。因為mtDNA與線粒體假基因同源,在核基因組中進化速率很低,所以在用通用引物進行擴增時很容易被擴增出來[17],如果線粒體假基因在進行系統發育研究分析時被當作線粒體基因序列來進行分析,分析結果可能會變得不準確或者錯誤[18-19]。COⅠ基因因其突變速度快、容易擴增、母性遺傳等特點成為分子系統發育研究中應用較為廣泛的分子標記。本研究采用COⅠ基因和16S基因作為分子標記,對2種土甲進行分子分類鑒定研究。通過對2種土甲不同蟲態的7條16S基因序列與7條COⅠ基因序列進行分析,根據堿基組成分析結果,2種土甲的堿基序列均表現出一定的AT偏向性,其中16S基因序列中A+T的平均含量為77.1%,G+C的平均含量為22.9%,這與劉曉麗[11]報道的擬步甲線粒體16S基因序列堿基構成的特點相符;COⅠ基因序列中A+T的平均含量為65.9%,G+C的平均含量為34.1%,這與趙亞茹[13]報道的擬步甲科昆蟲COⅠ基因的A+T含量相符。這表明本研究測得的基因序列是正確的。從2種土甲不同基因序列的同源相似度分析結果來看,粗背偽堅土甲種內16S基因序列同源相似度最高達100%,阿笨土甲種內16S基因序列同源相似度最高達99.56%,粗背偽堅土甲與阿笨土甲種間16S基因序列同源相似度最高為93.72%;粗背偽堅土甲種內COⅠ基因序列同源相似度最高達99.50%,阿笨土甲種內COⅠ基因序列同源相似度最高達99.24%,粗背偽堅土甲與阿笨土甲種間COⅠ基因序列同源相似度最高為91.75%。以上結果表明,不同物種間沒有完全一致的序列,同一物種不同蟲態不同齡期的個體間DNA序列差異很小。2種土甲種內不同個體間16S基因序列最大差異為1.03%,小于種類鑒定界限(2%);2種土甲種間16S基因序列最小差異為6.28%,高于限值(2%)。2種土甲種內不同個體間COⅠ基因序列最大差異為1.51%,小于種類鑒定界限(2%);2種土甲種間COⅠ基因序列最小差異為8.25%,高于限值(2%)。結果顯示2種土甲種內個體間與種間存在明顯的差異間隔,種內最大差異小于限值(2%),種間最小差異明顯高于限值(2%),與研究報道的種間差異>2%的結果一致[20]。由序列同源相似度可知1、2、3、4號為同一種,5、7、8號為同一種,這與形態學鑒定結果一致,表明DNA條形碼技術可以用來進行土甲族昆蟲幼期鑒定研究。
本研究采用16S與COⅠ兩種基因進行測定分析,突破單分子標記的局限性,當然本研究存在一定的不足之處,如研究標本采自同一地區,標本來源單一,若有不同采集地的標本進行對比研究鑒定,結果將更有說服力;本研究只是鑒定了幼蟲期的低齡和高齡幼蟲,對其他齡期的研究尚未進行。貢獻在于:為幼蟲種類鑒定提供新途徑,采用基于COⅠ基因和16S基因的DNA條形碼技術與常規形態學鑒定方法相結合的鑒定方式,可以更加準確快捷地進行種類鑒定研究。本實驗獲得的土甲族昆蟲的COⅠ基因序列可以作為該蟲的條形碼,進行快捷準確的鑒定,從而為害蟲及時有效防治提供科學依據。DNA條形碼技術除了可以彌補傳統形態學分類的諸多局限性外,還可以為分類學家發現新種提供一個新的方法[21],如果DNA條形碼技術能夠在土甲族昆蟲鑒定上得到充分利用并與常規形態學相結合,會給土甲族昆蟲的幼期鑒定帶來新突破。
