經濟蟹類性腺脂質需求的研究進展
2021-11-05 03:59:14張鳳英馬凌波
動物營養學報 2021年10期
傅 愔 張鳳英 趙 明 馬凌波
(中國水產科學研究院東海水產研究所,農業部遠洋與極地漁業創新重點實驗室,上海 200090)
我國甲殼類水產品資源豐富,種類繁多。隨著人民生活水平不斷提高,水產品在動物性食品中的占有率日趨壯大,而蟹作為傳統的水產佳肴一直備受喜愛,其養殖規模亦隨之膨脹。性腺發育是蟹類繁殖的基礎,良好的性腺發育水平可以提高蟹類的繁殖效率,同時為獲得大量優質的苗種提供保證。性腺是蟹類最主要的食用部位之一,性腺發育水平也決定其食用價值。相比未成熟的扣蟹,性成熟的雌蟹和雄蟹滋味更為鮮美,營養更為豐富。蟹類通常在生殖蛻殼后快速發育,根據卵細胞組成與形態的變化,雌蟹第1次卵巢發育分為6個時期(Ⅰ~Ⅵ期),卵巢發育從生殖蛻皮后的Ⅱ期開始,到Ⅴ期基本發育成熟,直至產卵[1-2]。對于雄蟹,精巢發育主要劃分為5個時期(Ⅰ~Ⅴ期),從Ⅰ期精原細胞期至Ⅳ期精子期精子成熟。
脂質作為蟹類性腺發育需要的主要營養素之一,直接影響其性腺的發育,關系到后續的交配繁殖、親本生殖力,甚至是胚胎發育以及內源營養期。此外,脂質作為蟹黃與蟹膏的主要成分,決定了成蟹的營養與食用品質。蟹類性腺中含有豐富的脂質,其中與性腺發育密切相關的脂質涉及各類脂肪酸,特別是二十碳五烯酸(EPA)、二十二碳六烯酸(DHA)、花生四烯酸(ARA)以及磷脂、固醇等。而在實際生產中,脂質的缺乏、過量、不平衡以及氧化均會導致蟹類性腺發育出現一系列的問題,嚴重制約蟹類養殖業的發展。目前,我國最主要的經濟養殖蟹類為中華絨鰲蟹(Eriocheirsinensis)、三疣梭子蟹(Portunustrituberculatus)和擬穴青蟹(Scyllaparamamosain)[3],其養殖產量逐年增高,2019年產量分別達到77.9萬、11.4萬和16.0萬t[4]。養殖過程中脂質的不合理攝入導致親蟹性腺發育不良、性早熟或者晚熟、肝胰腺白化癥、抱卵率下降,甚至流產等問題[5],進而造成成蟹營養價值降低、性腺發育不良、胚胎死亡率高、幼體育成活率低、優質苗種不能滿足產業需求等嚴重后果。因而,研究蟹類繁殖營養、關注性腺發育過程時脂質的調控作用是重點。本文聚焦中華絨螯蟹,同時綜述三疣梭子蟹和擬穴青蟹以作對比,就蟹類性腺發育階段脂質組成及變化規律、脂質在氧化供能與內分泌調控方面的生理作用,以及飼料脂質對性腺發育的影響3個方面作歸納總結,印證蟹類繁殖期科學合理地補充脂質營養對性成熟的促進作用。
1 蟹類性腺發育過程中脂質組成的變化
1.1 蟹類脂質的分類
蟹類中常見的脂質營養素包括不同飽和度、不同鏈長的脂肪酸,磷脂和固醇等。脂肪酸若依照鏈長分類,則12個碳以上均為長鏈脂肪酸。依照飽和度分類,可分為飽和脂肪酸、單不飽和脂肪酸(MUFA)和多不飽和脂肪酸(PUFA),其中PUFA也包含了有3個或以上雙鍵,且碳鏈長度大于20的高不飽和脂肪酸(HUFA)。典型的長鏈飽和脂肪酸有棕櫚酸、硬脂酸等,MUFA有油酸等,PUFA有亞油酸、亞麻酸、ARA等,而HUFA有EPA和DHA等。此外,根據不同的雙鍵位置,PUFA還能被分為n-3系列和n-6系列,亞麻酸、EPA和DHA均屬于n-3系列,而亞油酸、ARA屬于n-6系列。類脂中,磷脂是磷酸復合脂,為生物膜的重要組成成分,主要有甘油磷脂與鞘磷脂兩類。而環戊烷多氫菲衍生的膽固醇,是蟹類最主要的固醇類化合物。
1.2 蟹類性腺脂質的組成與變化
生殖期蟹類大量合成和積累高價值的脂質,用于構建生殖生理結構。中華絨螯蟹卵巢發育過程中,總脂含量呈先升高后降低的趨勢,于Ⅲ期達到最高[6]。而其主要脂肪酸含量也于Ⅲ期達到峰值,但飽和脂肪酸、DHA和磷脂含量于Ⅳ期達到峰值(表1)。擬穴青蟹雌蟹的卵巢發育過程中,磷脂和總脂含量也在Ⅳ期最高,此后降低,而各脂肪酸和固醇含量均在Ⅴ期達到最大值[7-8](表2)。目前公布的數據中,還未有關于三疣梭子蟹不同性腺發育時期各種脂質成分的變化,這是值得今后研究補充的空白。
值得關注的是,完全成熟時,性腺的脂質組成因性別和種類而異(表3)。中華絨螯蟹雄蟹精巢中以飽和的棕櫚酸、雌蟹卵巢以單不飽和的油酸較為突出[9-11],以儲存為主。各PUFA中,雌性中華絨螯蟹中亞油酸含量較高,雄蟹ARA和EPA含量較高[9]。此外,中華絨螯蟹雄蟹成熟后副性腺異常發達,占據生殖系統的絕大部分,副性腺發育過程中長鏈PUFA、固醇等脂質的含量在Ⅱ期最高,而它們在性腺中的含量于Ⅳ期最高[12]。
與中華絨螯蟹相比,三疣梭子蟹[9]和擬穴青蟹[13-14]有更高含量的EPA、DHA和PUFA,而青蟹的PUFA比例最高,并且含有大量磷脂[15]。并且,PUFA的n-3/n-6比值在2種海水蟹中亦較高[16]。此外,3種蟹類雄蟹生殖腺中的ARA均高于雌蟹[9,13-14]。
1.3 脂質的轉移促進性腺成熟
性腺發育時期,脂質從蟹的肝胰腺轉運至性腺,積累以促進性腺成熟。蟹類性腺和肝胰腺中脂質組成及含量隨著性腺發育階段的變化有一定的規律性。肝胰腺脂質在中華絨螯蟹雌蟹性腺發育Ⅲ期達到最大值29.85%,然后開始下降,直至產卵[6]。就卵巢脂質含量而言,中華絨螯蟹卵巢脂質則從Ⅱ期到Ⅳ期一直保持增長,并且在Ⅳ期達到最大值19.05%,然后在產后(Ⅴ期)降到最低值6.59%[6]。遠海梭子蟹(Portunuspelagicus)肝胰腺Ⅲ期的EPA含量增高,僅次于飽和脂肪酸[17],而三疣梭子蟹卵巢中的脂質含量也呈現出類似中華絨螯蟹的規律[18]。欖綠青蟹(Scyllaolivacea)卵巢中總脂、飽和脂肪酸、MUFA和PUFA的含量從Ⅰ期增加到Ⅲ期,Ⅵ期呈下降趨勢[19]。根據以上試驗結果,可以認為在卵巢發育過程中,Ⅲ期雌蟹卵母細胞迅速增長,為內源性卵黃合成期,此時肝胰腺脂質含量也達到高峰。而卵巢的脂質則在完全成熟期,當卵巢體積接近最大時才達到高峰。
性腺成熟與肝胰腺中脂肪酸轉運蛋白、脂肪酸結合蛋白、酰基轉移酶等的表達有關[20],表明其可能參與了脂質從肝胰腺到性腺的轉運。就雄蟹而言,發現脂肪酸結合蛋白在中華絨螯蟹睪丸快速生長中參與了脂質的轉運[21]。而在中華絨螯蟹雌蟹卵巢快速生長期脂肪酸結合蛋白也起到了類似的轉運作用[22]。此外,中華絨螯蟹卵巢發育過程肝胰腺和卵巢中催化三酰甘油酯合成的甘油-3-磷酸酰基轉移酶的表達水平從Ⅰ期開始升高,到Ⅳ期達到最高,然后急劇下降[23]。三疣梭子蟹的肝胰腺中發現至少4種脂肪酸結合蛋白用于介導脂質轉運[24]。青蟹肝胰腺轉運脂肪時,首先將吸收或存儲的脂質轉換為氧化穩定性更好、更易消化吸收的磷脂,同時伴隨著肝胰腺中的游離脂肪酸逐步增加,及時轉運到發育中的卵巢[16]。
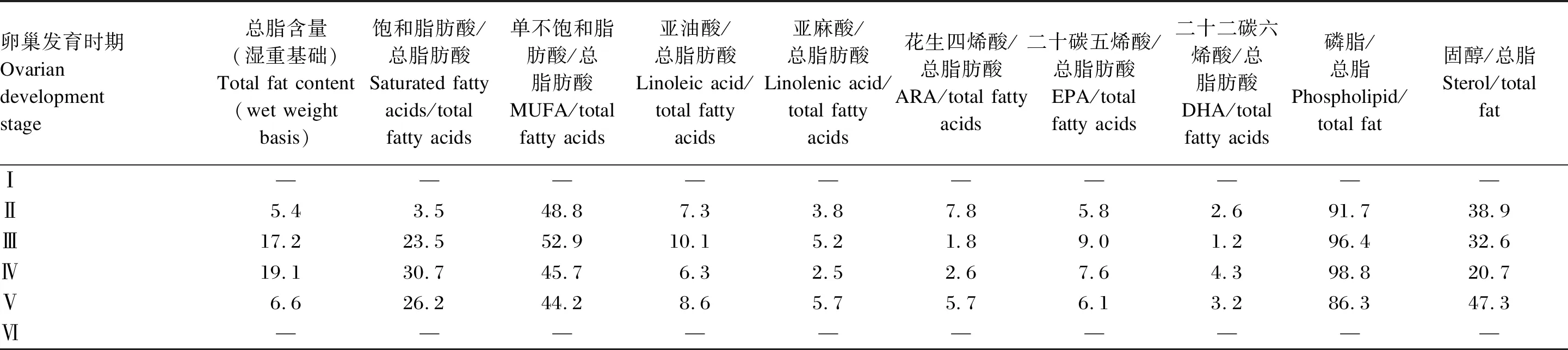
表1 中華絨螯蟹雌蟹卵巢發育過程中各脂質組成的變化
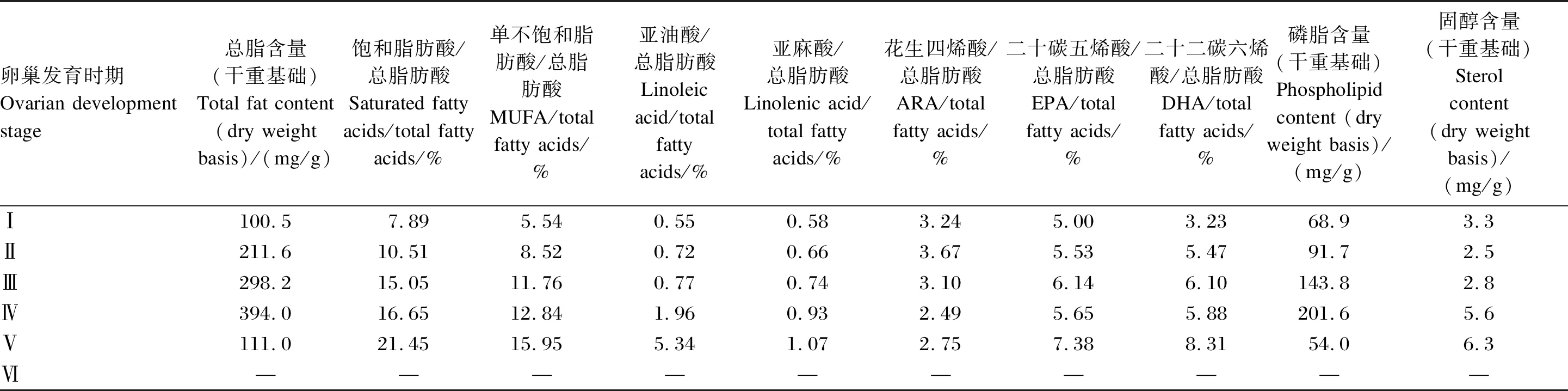
表2 擬穴青蟹雌蟹卵巢發育過程中各脂質組成的變化
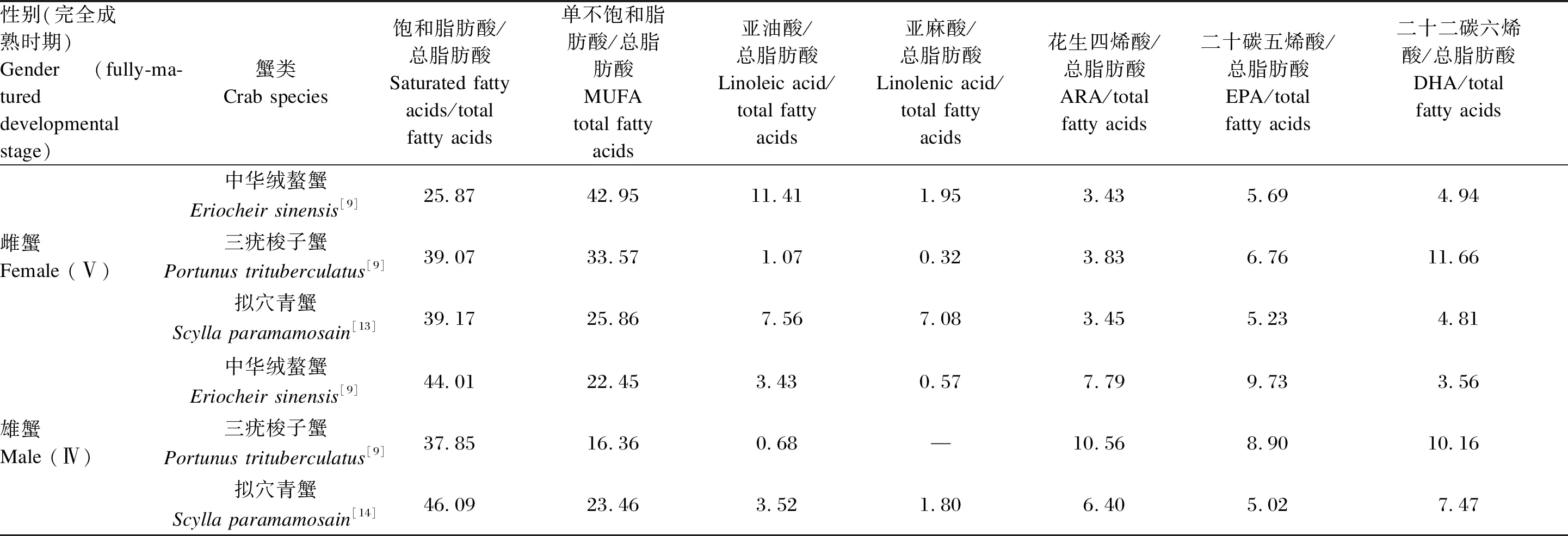
表3 3種經濟蟹類的雌、雄蟹成熟性腺脂肪酸的相對含量
參與轉運的脂質之中,PUFA是促進性腺發育的關鍵脂肪酸類別,其大量轉運主要集中在中華絨螯蟹雌蟹卵巢發育Ⅲ期后[25]。成永旭等[26]發現抱卵中華絨螯蟹肝胰腺中EPA和DHA含量明顯降低,而卵巢中的長鏈PUFA含量增加[27],這是由于抱卵蟹肝胰腺中高含量的DHA、EPA、ARA等部分轉移至卵巢,用于卵巢成熟、受精卵的發育、卵膜卵柄的形成等[10]。研究認為,不抱卵和流產蟹肝胰腺的PUFA未能有效轉運至卵巢[28]。在不抱卵的中華絨螯蟹的卵巢中發現EPA和DHA含量較抱卵蟹低,而抱卵蟹流產后肝胰腺PUFA的含量明顯上升,其中變化最顯著的為DHA與EPA的含量之和,大約上升3.7倍[10],推測其滯留在肝胰腺未能轉移至卵巢,使卵巢發育不良,且胚胎得不到足夠的營養和能量。
1.4 性腺脂質積累的抗氧化保護
生殖期親蟹性腺對其自身的抗氧化保護需求要高于其他時期,維生素、礦物質和類胡蘿卜素等都對積累的脂質起到一定的抗氧化保護作用。維生素C和維生素E能抑制中華絨螯蟹雌蟹卵子脂質過氧化發生,降低脂質過氧化物丙二醛含量,提高血清超氧化物歧化酶和谷胱甘肽過氧化物酶的活性[29]。維生素C、維生素E的協同作用還可防止細胞和亞細胞膜中磷脂和固醇等不飽和脂肪酸的過氧化反應,使中華絨螯蟹體內自由基達到動態平衡。蝦青素能幫助維持中華絨螯蟹[30]卵巢的總脂含量,使之呈上升趨勢,這種抗氧化作用同樣適用于三疣梭子蟹[31]。礦物質的抗氧化保護主要體現在硒元素上,能激活擬穴青蟹的谷胱甘肽過氧化物酶,保護性腺脂質[32]。此外,β-胡蘿卜素在性腺成熟過程中于青蟹卵巢中的沉積也能有效清除自由基,阻止生物膜上含PUFA的脂質和蛋白質氧化[33]。由此可知,適量補充維生素C、維生素E、蝦青素、硒、β-胡蘿卜素等抗氧化營養素有利于保護親蟹性腺的脂質營養組成。
2 脂質在性腺發育過程中的生理功能
2.1 脂質氧化為性腺發育供能
脂質的消化分解為蟹類性腺發育提供能量。中華絨螯蟹肝胰腺的脂肪酶活性在卵巢成熟期逐漸上升[34]。三疣梭子蟹肝胰臟中三酰甘油脂肪酶的mRNA表達水平隨著卵巢的發育也有所提高[24]。三酰甘油等中性脂肪是主要提供能量的脂質,這點可在大量表達的脂質代謝基因中得到證實。性腺發育期,在三疣梭子蟹肝胰腺轉錄組中發現與線粒體脂肪酸氧化供能有關的酶基因包括酰基輔酶A脫氫酶、四醇脫氫酶、線粒體樣D-β-羥基丁酸脫氫酶和短鏈脫氫酶等[24]。雄蟹精子能量的主要來源也被認為是脂質的氧化作用[35]。
2.2 脂質參與內分泌調控
生殖期,親蟹體內的部分脂質充當了性腺發育過程中激素合成的前體物質參與體液調節。比如,膽固醇是性激素、蛻皮激素、腎上腺皮質類固醇激素等生物合成的前體。因此,在卵巢發育階段,中華絨螯蟹的總膽固醇含量以及肝胰腺中膽固醇酯的含量呈現上升的趨勢[20]。再如,ARA通過環氧合酶、前列腺素合成酶轉化為前列腺素。前列腺素抑制脂肪的分解,使肝胰腺中脂肪的積累大于分解,并部分轉移至卵巢,促進親蟹卵巢成熟。隨著卵巢進一步發育和卵子的產出,抱卵蟹肝胰腺中ARA的含量降低[10],此時脂肪的分解加強,給受精卵提供其余必需的脂肪酸。另外,除了作為前體物質,過量的EPA和DHA還能與三疣梭子蟹前列腺素的中間產物結合[36],抑制其對卵巢發育的生物活性。青蟹卵巢和血淋巴中的固醇類激素含量在卵黃發生初期表現為類似的上升趨勢[37],這種上升促進了性激素的合成和性腺的增長與成熟。
此外,脂質本身對生殖內分泌也存在一定的調控作用。脂質可以影響中華絨螯蟹眼柄軟組織蛻皮抑制激素(MIH)的基因表達,但不同脂質對其表達的作用相反[25]。當中華絨螯蟹飼料脂肪含量由6%增加到15%時,MIHmRNA的表達明顯受到抑制,并且魚油(n-3 PUFA)也能明顯抑制眼柄中MIHmRNA的表達,利于性腺發育。反之,豬油(飽和脂肪酸)和磷脂促進中華絨螯蟹MIHmRNA的表達,磷脂可以提高雌二醇的含量[25]。非基因組的作用機制中,膽固醇能通過長時間孵育作用調控中華絨螯蟹眼柄神經分泌細胞的鈣電流,通過對鈣離子內流的抑制,影響動作電位的傳播,調制神經肽激素分泌[38],影響生殖發育。對于三疣梭子蟹來說,飼料中添加4%的大豆卵磷脂可以提高血清中類固醇激素(孕酮和雌二醇)的含量,并增大卵母細胞直徑[39],這可能是類固醇激素生物合成代謝途徑基因表達上調所致[40]。再者,磷脂的攝入能降低雌激素硫酸轉移酶的表達,提高雌激素活性[40]。
因此,不同類型的脂質可以通過轉錄水平、神經水平和體液水平等多種途徑來調節性激素的分泌,進而影響蟹類性腺的成熟。
3 脂質營養對蟹類性腺發育的影響
蟹類的天然餌料包括魚、蝦、貝、蜆、螺、沙蠶等,種類繁多。但品質不穩定,且受季節影響較大,經常缺乏必需脂肪酸、HUFA、磷脂等,導致親蟹質量降低。因此,脂質的合理攝入能夠改善親蟹的性腺發育水平。國內外研究表明,親蟹各組織尤其是肝胰腺、性腺和胚胎的脂肪酸組成與攝入飼料的脂肪酸組成呈正相關[41-43]。飼養試驗在揭示攝入脂質與性腺成熟關系的同時,也有助于了解親蟹對不同脂質的需求量、平衡比例、來源的選擇,以及脂質攝入過量和不平衡導致的發育不良、性早熟、肝胰腺白化癥等不良后果。一般來說,性腺指數(GSI)和肝胰腺指數(HI)能在一定程度上反映甲殼動物卵巢和肝胰腺中營養積累和發育的狀況,因此可以作為評價性腺發育水平的指標。
3.1 脂質攝入量對性腺發育的影響
親蟹在性腺發育中需要大量磷脂和PUFA,但從頭合成的能力有限[44],因此必須從外界攝取。飼養試驗揭示了不同雌蟹繁殖期性腺成熟對PUFA和磷脂的需求和適宜添加量(表4)。有關雄蟹繁殖期對PUFA和磷脂的需求量尚未有具體研究。一個有趣的現象是,雖然HUFA是性腺和胚胎正常發育的重要保證,但試驗表明中華絨螯蟹親蟹對DHA、EPA、ARA的需求量并不高,性腺發育只要滿足其最低需求量[45],這可能是因為性腺和胚胎中的脂肪酸組成具有較強的保守性。但也有研究認為適量添加DHA和EPA對中華絨螯蟹卵巢發育作用顯著[25,41],而且可以顯著提高初孵幼體的成活率、變態率和抗逆境能力[46]。
相反,過多攝入脂質,特別是固醇、飽和脂肪酸等,以及生產中投喂動物性餌料過多,則會導致養殖中華絨螯蟹的性早熟。膽固醇在中華絨螯蟹體內積累可直接促進性腺促進激素和法尼酸甲酯及17-β雌二醇、睪酮等甾醇類性激素的合成和分泌,造成性腺發育過快,成熟過早[47]。另外,高蛋白質攝入也可能導致中華絨螯蟹的膽固醇攝入增加[48],誘導肝胰腺和全身的脂質沉積,并且過多的營養轉移到性腺導致早熟。此外,含飽和脂肪酸較高的花生油能顯著提高中華絨螯蟹類固醇類激素含量,豬油次之,但過量添加也極可能造成過早發育[25]。飽和脂肪酸攝入量超過34%時中華絨螯蟹有早熟現象發生[49]。據此,控制飼料的脂質含量,并適當補充磷脂和HUFA對中華絨螯蟹性早熟存在一定的反作用,能夠在轉錄水平上促進脂肪氧化輸出,減輕高脂飼料造成的代謝負擔[50-51]。
因此,中華絨螯蟹性腺成熟需要DHA、EPA與ARA的適量添加,對于初始體重75 g以上的雌蟹,推薦的HUFA和磷脂添加量在2%左右。對于初始體重更小的雌蟹,可增加DHA、EPA和磷脂的添加量至6%以上。而飼料中飽和脂肪酸和固醇的添加量應予以控制,避免中華絨螯蟹的早熟和發育不良,影響親蟹的營養品質及繁殖性能。
與中華絨螯蟹相似,三疣梭子蟹對DHA、EPA、ARA的需求要也不高,僅需滿足其最低需求量[36,43,52],并且HUFA含量過高時對梭子蟹有一定的負面影響[52]。ARA的添加量只需在0.06%~0.24%的范圍內,過度添加易造成梭子蟹卵巢水分含量過高,阻礙卵膜卵柄發育[36]。
3.2 不同脂質攝入比例對性腺發育的影響
不同脂質與脂肪酸在親蟹體內的比例平衡十分重要。中華絨螯蟹飼料中添加卵磷脂后飽和脂肪酸含量下降,MUFA、PUFA、HUFA含量的上升有利于卵巢發育[54]。DHA/EPA、n-3/n-6、磷脂/HUFA的配比適當,對促進雌蟹性腺成熟,提高卵子受精、孵化率起著重要作用(表5)。中華絨鰲蟹生長所需DHA/EPA比例為2左右[55]。過高或者過低的DHA/EPA比例都會抑制營養物質的積累,甚至造成肝胰腺白化癥。飼料中n-3和n-6 PUFA的動態平衡對性成熟十分重要,單純追求高營養價值的n-3 PUFA的含量而忽略了n-6 PUFA的含量可能會適得其反。中華絨螯蟹卵巢發育飼料中n-3/n-6比例至少為2時有較好的性腺發育效果[56],磷脂/HUFA的比例處于1.6/2.5~2.3/2.0時也有助于維持中華絨螯蟹體內n-3和n-6 PUFA的平衡[45],促進卵巢發育。
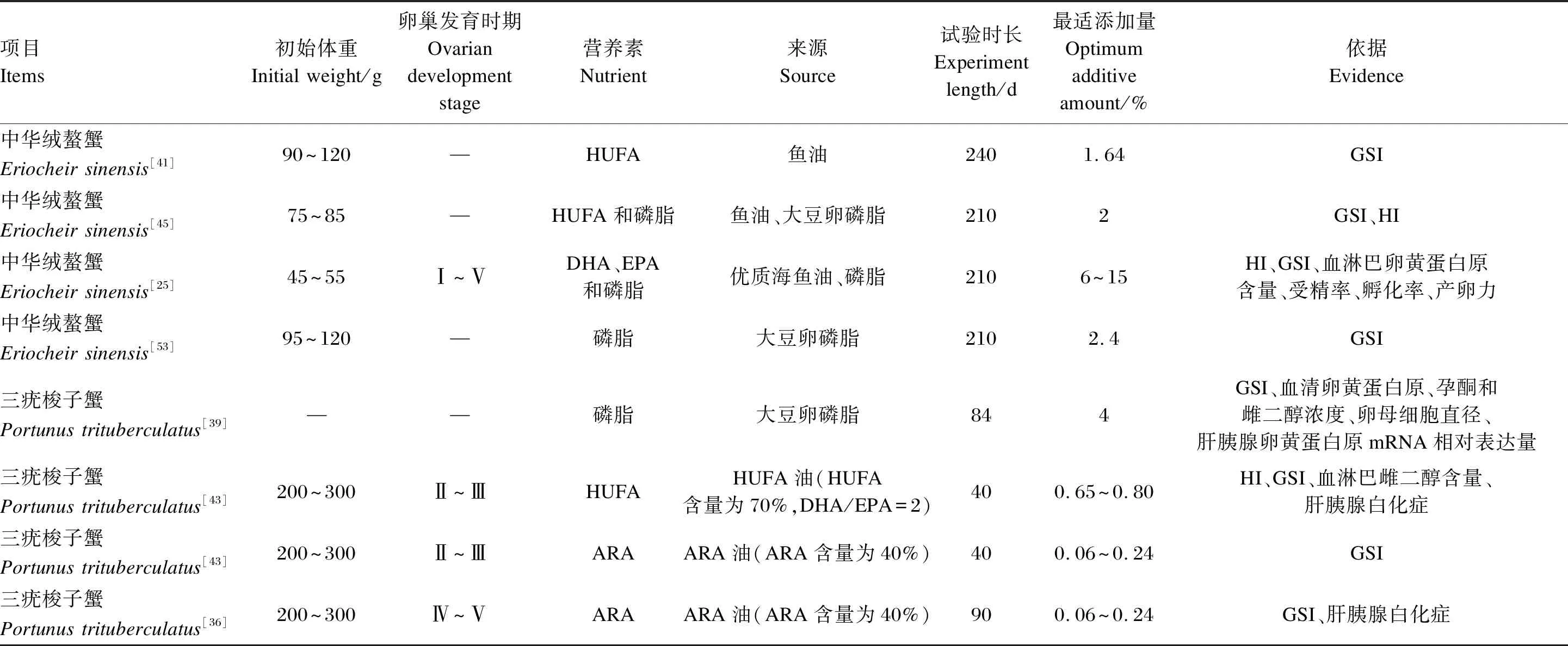
表4 雌蟹繁殖期多不飽和脂肪酸和磷脂添加量的飼養試驗結果
與中華絨螯蟹相似,配合飼料中較高的DHA/EPA比例也更有利于三疣梭子蟹肝胰腺營養物質的積累和卵黃物質合成,適宜的比例至少為2[43,52,57]。但HUFA含量過高,油酸、亞油酸和亞麻酸含量過低,則不利于色素在三疣梭子蟹肝胰腺中的沉積,或者是提供能量的飽和脂肪酸和MUFA過少,不能保證梭子蟹肝胰腺正常營養物質的合成及代謝,存在較大氧化壓力,甚至會破壞肝胰腺的正常功能[36,43,52]。再者,ARA可能會抑
制n-3 HUFA的合成,并且ARA也可與EPA和DHA競爭與前列腺素的轉化酶系統相結合,影響梭子蟹的脂質利用[36]。
由此可知,繁殖期應注意親蟹飼料DHA/EPA和n-3/n-6比例為2最佳,磷脂/HUFA比例為0.6~1.2,以及MUFA、PUFA、HUFA等的適量補充,利于蟹類性腺發育,避免肝胰腺白化癥等問題的發生。

表5 雌蟹繁殖期飼料脂質最適比例的飼養試驗結果
3.3 脂質來源對性腺發育的影響
混合植物油替代魚油可以平衡飼料中的脂肪酸組成,也能夠影響親蟹的性腺發育和繁殖性能。魚油被植物油替代能夠降低脂肪吸收和脂肪酸生物合成,增加β-氧化,并且增加Δ6脂肪酸脫氫酶的表達水平[58]。隨著植物油含量的增加,飼料中的亞油酸和亞麻酸含量顯著增加,這可能在一定程度上可提高中華絨螯蟹的生殖力[59]。添加魚油與豆油比例為2∶1和1∶1時,更有利于中華絨螯蟹卵巢發育,比例為1∶2時次之[60]。使用50%~75%植物油替代飼料中魚油喂養雌性中華絨螯蟹可以適當提高其產卵量[11,60]。因而,在33%~75%替代水平內,對雌蟹卵巢發育都是有益的。對于雄蟹,研究認為混合植物油(豆油∶菜籽油=1∶1)替代不同水平魚油對中華絨螯蟹雄蟹性腺發育無顯著影響[61],其適宜的替代水平為25%~50%。對三疣梭子蟹來說,50%的魚油替代水平可明顯提高卵巢和肌肉中蛋白質含量,而100%的魚油替代水平會顯著降低梭子蟹肝胰腺和卵巢中HUFA的含量[62]。
就磷脂源而言,磷蝦油比大豆卵磷脂和蛋黃卵磷脂更有效促進中華絨螯蟹肝胰腺和卵巢的脂質沉積,主要是極性脂質和n-3 PUFA的沉積[54]。此外,磷蝦油對中華絨螯蟹卵黃發生的促進作用比其他來源的磷蝦油更為明顯,其抗氧化效果也最好。
因此,建議選擇33%~75%植物油替代魚油,能夠提高親蟹的性腺發育水平,而磷蝦油的抗氧化效果使之成為最有利于不飽和脂肪酸在性腺積累的磷脂源。
4 小結與展望
對蟹類性腺發育各時期脂質組成的研究闡明了脂質在性腺發育期的轉移規律,這可能和脂質在性腺發育的氧化供能與內分泌調控方面的生理功能相關。根據脂質攝入量、不同脂質的攝入比例和來源等試驗數據,分析脂質攝入失衡的后果,認為滿足不同蟹類性腺對脂質的需求,都應適量補充DHA、EPA、ARA、固醇、磷脂等,并且注意配比合理。來源的選擇方面,部分植物油替代魚油是有利于性腺發育的。同時,配合補充維生素C、維生素E、硒、β-胡蘿卜素、蝦青素等天然抗氧化劑,并且采用抗氧化能力強的磷蝦油作為磷脂源,能有效保護性腺脂質的積累。
不過,目前對于蟹類脂質生理作用的理解不夠完善,飼料營養的配比還未到最優,因此應從如下方面進一步開展脂質營養調控的研究:1)繼續闡明蟹類的性腺發育過程中脂質代謝相關的功能基因,并且開展營養基因組學研究,分析在各種脂質缺乏、適宜和過剩條件下基因表達情況,以期從分子角度為脂質調控性腺發育提供參考依據;2)探索脂質生殖調控作用下可能存在的反饋機制,即固醇類激素是否促進了性腺脂肪的堆積,而積累到一定程度的性腺脂質是否又反過來調控激素的分泌等;3)在傳統的干預試驗中引入營養遺傳學的思路以豐富營養調控蟹類性腺發育的研究,比如不同遺傳背景、家系的親蟹是否對脂質營養有不同需求,應通過特異性的飼料配方有針對地進行飼喂,此外,營養學與遺傳學結合也有助于發現蟹類性腺發育中與脂質代謝相關的特殊遺傳變異;4)目前的研究仍存在偏重雌蟹的問題,以后應同時關注雄蟹性腺發育的營養生理與需求。
綜上所述,蟹類繁殖營養學的研究為高利用率親蟹營養強化飼料的開發奠定基礎,而脂質營養在性腺發育期的平衡攝入能夠提高親蟹的營養價值,優化其繁殖性能,推進經濟蟹類苗種產業的高效、健康、穩步發展。
