新疆野蘋果幼苗葉片響應NaCl脅迫的miRNAs及靶基因篩選
2021-10-30 01:26:00何晨晨劉俐君魯曉燕
新疆農業科學 2021年4期
何晨晨,劉俐君,魯曉燕
(石河子大學農學院/特色果蔬栽培生理與種質資源利用兵團重點實驗室,新疆石河子 832000)
0 引 言
【研究意義】鹽脅迫是植物生長發育過程中的重要限制因子,MicroRNA(miRNA)在植物應答非生物脅迫的過程中具有重要的調控作用。miRNA是位于基因組非編碼區的內源性的小分子單鏈RNA,長度多為20~24 bp[1],可特異識別靶mRNA并與之結合,降解靶mRNA、抑制翻譯、甲基化,負向調控基因表達[2],miRNA可以在轉錄水平、轉錄后水平和翻譯水平參與植物體內的不同生物學過程[3],植物的生長發育、信號轉導以及對脅迫的應答等[4]。在植物抵御鹽脅迫過程中,miRNA在調控植物種子萌發、花芽分化、形態建成等發揮重要作用[5]。新疆野蘋果(Malussieversii(Ledeb.)Rome.)屬薔薇科(Rosaceae),蘋果屬(MalusMill.)植物[6],新疆野蘋果主要分布在中亞地區的天山山脈,以及我國新疆西部伊犁和塔城地區,具有很強的生態適應性[7];新疆野蘋果作為我國西部地區的蘋果生產的一個重要砧木,能耐極端最低溫-30℃左右,對溫度上限不敏感,耐旱力較強,耐瘠薄以及抗病蟲害等多種優良的抗逆性狀[8]。尹蓉[9]研究表明,對15種主要蘋果屬植物的抗逆性相關生長指標及抗逆系數分析表明,新疆野蘋果屬耐鹽性中等蘋果屬植物資源。于瑋瑋等[10]研究表明,新疆野蘋果幼苗在NaCl脅迫下可通過積累脯氨酸和可溶性糖,提高SOD和POD的活性,以減緩NaCl脅迫對幼苗造成的傷害且具有一定的耐鹽性。【前人研究進展】Xu等[11]利用Illumina測序法對紅肉甜橙研究顯示,51個已知的miRNAs在突變型(MT)和野生型(WT)之間有顯著的表達差異。Pantaleo等[12]在葡萄中鑒定出24個保守的miRNA家族,26個已知但不保守的miRNA。王永江等[13]對甜橙miRNAs進行了鑒定,預測并篩選出其調控的靶基因與植物與病原物互作相關。Niu等[14]研究發現,梨樹與休眠相關的MADS-box基因和miRNAs共同控制著梨花芽休眠的轉變。Sun等[15]研究表明,柑橘miRNAs及其靶基因與植物的發育、轉錄、蛋白質降解等多種細胞過程有關。【本研究切入點】呂新民等[16]預測miRNA及其靶基因與酸棗生長發育、轉錄調節等相關,其中一些基因可能參與酸棗響應NaCl脅迫過程。研究借助先進的Illumina測序平臺對新疆野蘋果NaCl脅迫應答的Small RNA進行測序,快速、高效地讀取高質量的測序數據。【擬解決的關鍵問題】對150 mmol/L NaCl處理6和48 h時的新疆野蘋果幼苗葉片進行small RNA測序。篩選差異表達的miRNA及其靶基因,為分析新疆野蘋果的耐鹽機制提供參考。
1 材料與方法
1.1 材 料
供試材料為新疆野蘋果種子,購買于新疆伊犁哈薩克自治州霍城縣61團。
新疆野蘋果種子經4℃低溫層積處理90 d左右。發芽后播于進口泥炭∶蛭石=3∶1混勻的基質中,放置于人工氣候箱(RXZ智能型,寧波江南儀器廠)中培養,培養條件為:溫度(25±1)℃,相對濕度65%~80%,光照強度12 000 lx,發芽期黑暗狀態,出苗期光照白天16 h,黑夜8 h。等種子發芽長出6~8片真葉時選取生長健壯的幼苗作為試材,剪取約2~3 cm的莖端幼嫩組織接入培養基中進行誘導分化培養,誘導培養基為:MS培養基+0.5 mg/L IBA+2 mg/L 6-BA;繼代增殖培養基為:MS培養基+0.5 mg/L IBA+2 mg/L 6-BA,每30 d繼代1次;連續幾代增殖培養后選取生長健壯的莖段,接入生根培養基中培養,生根培養基為:1/2MS培養基+0.5 mg/L IBA;所有培養基中均加蔗糖30 g/L、瓊脂粉7 g/L,用1 mol/L的NaOH調節pH值到5.8~6.0。組培苗生根后挑選根系生長較好,長勢健壯且整齊一致的幼苗轉入有1/2日本園式營養液水培盒中,培養1周后轉入完全營養液中。將水培盒放入人工氣候箱內進行培養,在培養箱外的一側放置供氧泵,持續通氧。
1.2 方 法
1.2.1 試驗設計
水培苗第6~8片葉完全展開時,處理:(1)對照(CK),日本園式營養液;(2)NaCl處理,日本園式營養液加150 mmol/L NaCl。NaCl濃度以每天50 mmol/L的梯度逐步遞增,全部處理于同1 d達到目標濃度,設此時為NaCl處理0 h。分別于處理6和48 h取新疆野蘋果幼苗葉片,樣品用清水沖洗表面雜物,再用去離子水沖洗干凈,用吸水紙吸干表面水分,稱取0.1 g,裝入1.5~2 mL的凍存管中,對照的葉記作LCK6h、LCK48h;NaCl處理的葉記作LNa6h、LNa48h,迅速投入液氮中冷凍,于-80℃冰箱保存備用。每個處理重復3次。
1.2.2 新疆野蘋果sRNA文庫構建
提取樣品的總RNA,small RNA測序由南京派森諾基因科技有限公司完成,質檢合格后進行文庫構建。將TotalRNA使用PAGE電泳切膠分離成18~30 nt的RNA,利用Blast將去接頭、去低質量、去污染的Clean Reads與Rfam13數據庫比對,統計分析數據庫中小RNA的種類、數量和類型,并對其注釋。
1.2.3 NaCl脅迫下miRNAs差異表達及靶基因GO和KEGG顯著性富集
對新疆野蘋果葉片差異表達的靶基因按照分子功能(MolecularFunction)、生物過程(Biological Process)和細胞組分(CellularComponent)進行GO分類,挑選每個GO分類中挑選p-value最小即富集最顯著的前10個GOterm條目進行展示。
采用DESeq(version1.18.0,Anders S和Huber W,2010)分析4個處理中表達差異miRNA,按照表達量倍數差異|foldchange|>2和表達差異顯著性p-value<0.05篩選出差異的保守miRNA。miRNA主要通過互補配對結合到靶位點。以該物種的mRNA的3‘UTR’序列為目標序列,對差異表達的miRNA序列,使用psRobot_tar進行靶基因預測,并對靶基因進行GO和KEGG顯著性富集分析。
1.2.4 NaCl脅迫下新疆野蘋果葉片差異miRNA與mRNA的反轉錄和實時熒光定量PCR
miRNA反轉錄采用miRcute Plus miRNA First-Strand cDNA kit試劑盒(TIANGE,北京)合成cDNA,qRT-PCR使用miRcute Plus miRNA qPCR Kit試劑盒(TIANGE,北京),采用20 μL反應體系:cDNA模板2 μL,2×miRcute Plus miRNA PreMix 10 μL,上下游引物(10-M)各0.4l,超純水7.2l。qRT-PCR在CFX96 Real-Time PCR儀(Bio-Rad,美國)上進行,反應程序為:95℃預變性15 min,94℃ 20 s,60℃ 34 s,45個循環。
mRNA反轉錄采用5X All-In-One RT MasterMix試劑盒(abm,加拿大)合成cDNA,qRT-PCR使用Green Real time PCR Master MIX試劑盒(TOYOBO,日本),采用20 μL反應體系:cDNA模板2 μL,2×SYBR Green Mix 10 μL,上下游引物(10 μM)各0.5 μL,超純水7 μL。qRT-PCR在CFX96 Real-Time PCR儀(Bio-Rad,美國)上進行,反應程序為:95℃預變性1 min,95℃ 10 s,55℃ 30 s,72℃ 30 s,40個循環。利用Primer 6軟件設計qRT-PCR引物,設計好的引物由生物工程(上海)股份有限公司合成。miRNA內參基因為5srRNA[17],mRNA內參基因為UBQ[18]。基因的表達量采用相對定量的方法,即2-△△CT法[19]。表1
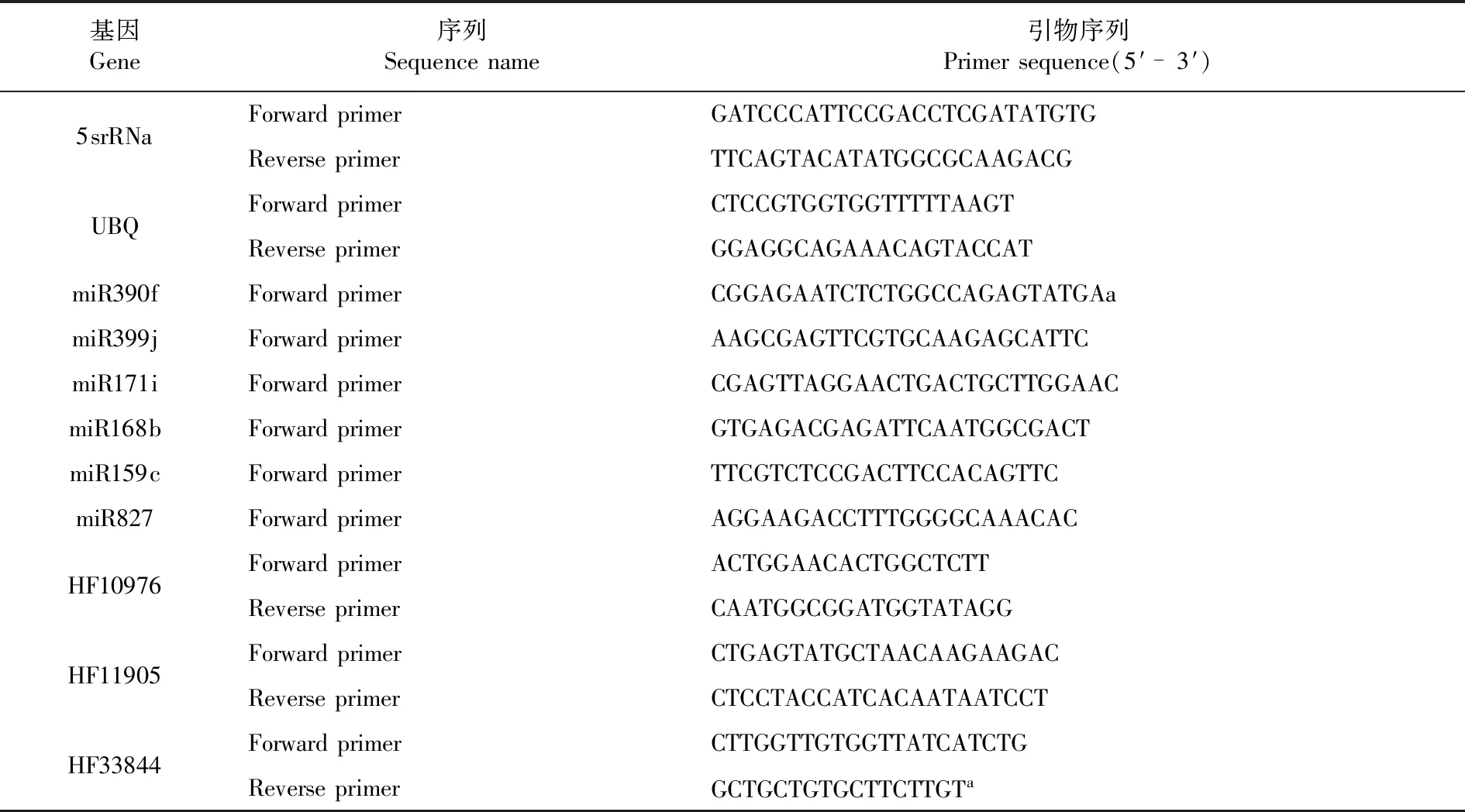
表1 qRT-PCR引物序列Table 1 The sequences of the primers for qRT-PCR
2 結果與分析
2.1 新疆野蘋果葉片不同處理RNA質量檢測
研究表明,每個樣品的RNA完整值(RNA integrity number,RIN)均大于7.5,OD260/280均大于2.0,OD260/230均大于1.6,28S/18S均大于1.5,符合建庫標準。表2
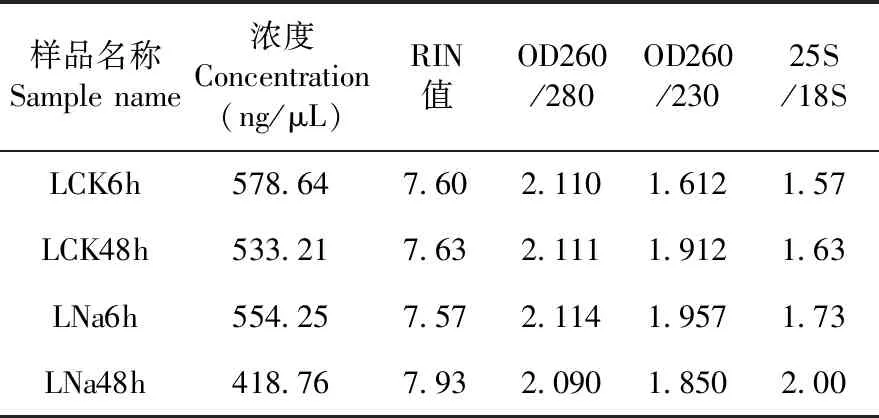
表2 不同處理RNA質量檢測Table 2 Quality detection of RNA in different treatments
2.2 新疆野蘋果葉片不同處理small RNA分類注釋
研究表明,測序分別從LCK6h、LCK48h、LNa6h、LNa48h 4個文庫中獲得35 350 971、34 458 835、43 963 615、35 699 217條Raw Reads,去除污染序列,去除沒有插入片段的reads,去掉包含polyA的序列和小于18 nt的小片段,分別得到29 610 812、28 904 431、41 122 379、33 401 117條Clean Reads,主要包含rRNA、tRNA、snRNA、snoRNA、known miRNA、novelmiRNA、以及unknown片段等。新疆野蘋果葉片LCK6h、LCK48h、LNa6h、LNa48h 4個處理中Unique Known miRNA數量分別為6 331、5 561、5 825、6 074,Unique Novel miRNA數量分別為35、35、37、42。表3
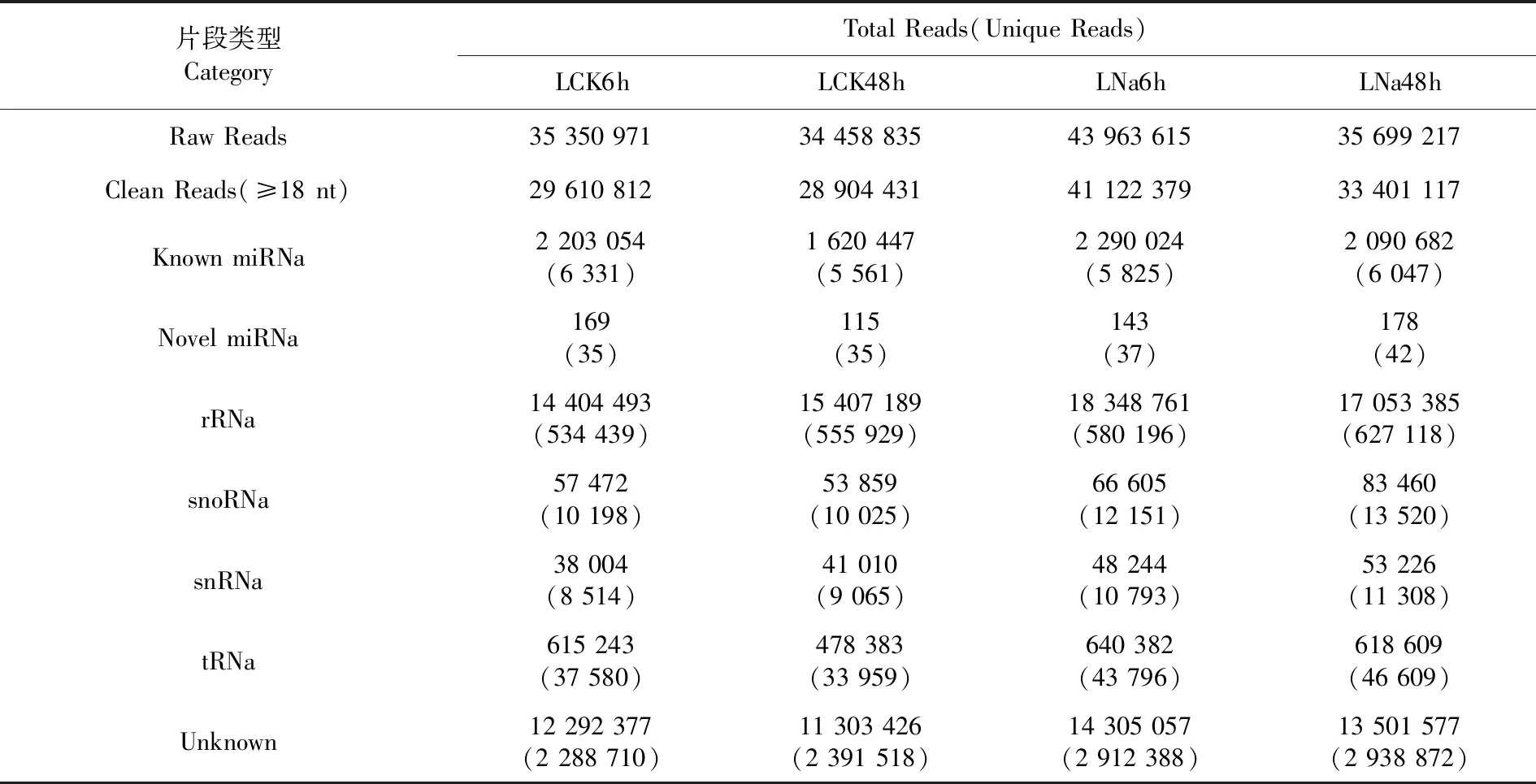
表3 新疆野蘋果數據過濾及small RNA分類注釋Table 3 Data filtering and small RNA classification annotation statistics of Malus sieversii
2.3 新疆野蘋果Clean Reads長度分布
研究表明,在LCK6h、LCK48h、LNa6h、LNa48h 4個處理中Total Reads的長度主要分布在20~24 nt,長度為24 nt的序列所占比例最多,其次是長度為21和20 nt的序列。Unique Reads的長度分布最多的均為24 nt。圖1
2.4 NaCl脅迫下差異miRNA及靶基因預測
研究表明,在新疆野蘋果已知的miRNA中,與對照相比,NaCl處理6 h時的差異miRNA數量為3個,其中上調表達的為2個,下調表達的為1個,靶基因數目為64個。NaCl處理48 h時的差異miRNA數量最多為13個,其中上調表達的為4個,下調表達的為9個,靶基因數目為108個。與NaCl處理6 h相比,NaCl處理48 h時差異miRNA數量為5個,其中上調表達的為3個,下調表達的為2個,靶基因數目為39個。NaCl處理6 h和48 h共有的差異miRNA有2個。表4
2.5 NaCl脅迫下miRNA靶基因GO顯著性富集
研究表明,NaCl處理6 h時,GO主要富集的分子功能MF主要包括:肌醇磷酸2激酶活性、寡糖基轉移酶活性、寡糖基轉移酶活性等。細胞組分CC主要包括:網格蛋白復合物、網格蛋白包被的坑等。生物過程BP主要包括:蛋白絲氨酸/蘇氨酸磷酸酶抑制劑活性、木聚糖代謝過程等。NaCl處理48 h時,GO主要富集的分子功能MF主要包括:氧氧化還原酶活性、DNA結合等。細胞組分CC主要包括:質外體、胞外區等。生物過程BP主要包括:木質素代謝過程、苯丙烷分解代謝過程、次生代謝過程等。圖2
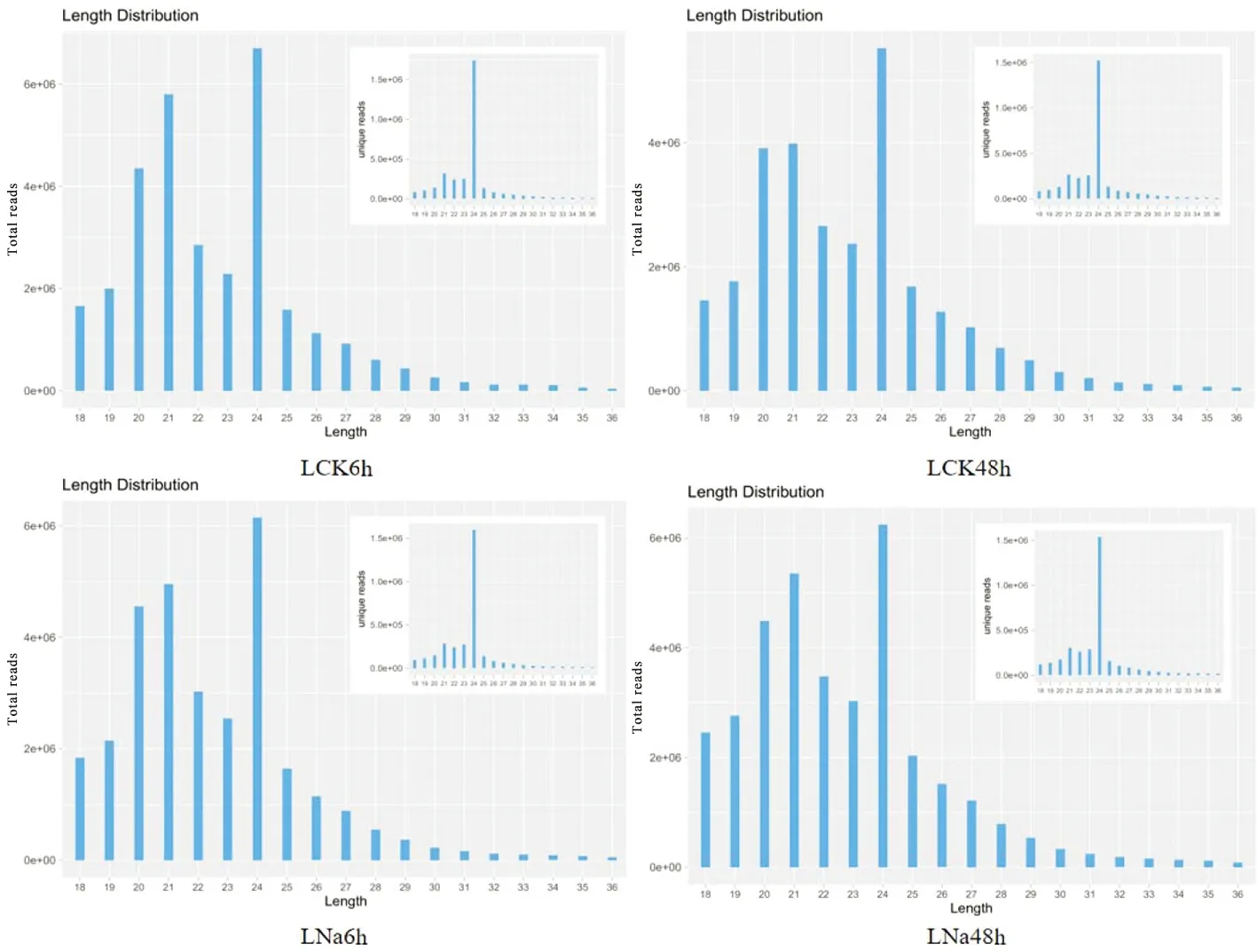
注:橫軸為不同的序列長度,縱軸為對應長度的序列豐度(*10 000)。大圖為去重前不同長度的Total Reads分布,右上角小圖為去重后不同長度的Unique Reads分布Note:The horizontal axis is different sequence length,and the vertical axis is the sequence abundance of corresponding length(*10,000).Large image shows total reads of different lengths before deweighting Distribution:The small figure in the upper right corner shows the distribution of Unique Reads with different lengths after deduplication圖1 新疆野蘋果miRNA 長度分布Fig.1 Distribution of miRNA length in Malus sieversii

表4 NaCl脅迫下新疆野蘋果差異表達miRNA及靶基因Table 4 differential expression of miRNA and target genes in Malus sieversii under NaCl stress
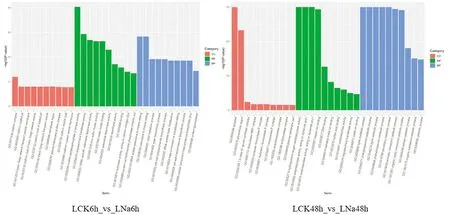
圖2 新疆野蘋果葉片miRNA的差異靶基因Go功能分類Fig.2 Go functional classification of differential target genes of miRNA in Malus sieversii
2.6 NaCl脅迫下miRNA的靶基因KEGG顯著性富集
研究表明,NaCl處理6 h時,主要富集在N-glycan生物合成、N-磷脂酰肌醇信號系統、色氨酸代謝、苯丙氨酸代謝、光合生物中的碳固定等。NaCl處理48 h時,主要富集在N-glycan生物合成、其他聚糖降解、煙酸酯和煙酰胺代謝、丙酸酯代謝、真核生物的核糖體生物發生等通路。NaCl處理6和48 h時共同富集的是N-glycan生物合成通路。圖3
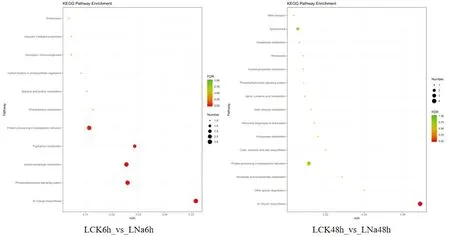
圖3 新疆野蘋果葉片miRNA的靶基因KEGG通路注釋Fig.3 KEGG pathway annotation of miRNA target gene in Malus sieversii
2.7 qRT-PCR驗證差異表達的miRNA
研究表明,在新疆野蘋果葉片差異顯著的miRNA中選取6個miRNA和3個mRNA,mdm-miR168b、mdm-miR159c、mdm-miR827、mdm-miR390f及其靶基因HF10976、mdm-miR171i及其靶基因HF33844、mdm-miR399j及其靶基因HF11095。利用qRT-PCR檢測基因的表達量,與測序數據趨勢基本一致,且miRNA與其靶基因mRNA的表達量呈負相關。圖4

圖 4 qRT-PCR驗證結果Fig.4 qRT-PCR validation data
3 討 論
miRNA最早于發現線蟲中[20],大量miRNA在擬南芥[21],玉米[22],番茄[23],水稻[24]等植物中也相繼被研究發現。并且各類果樹作物miRNA的研究也日趨成熟,已在蘋果[25-26]、柑橘[27-28]、草莓[29]、葡萄[30-31]、桃[32]等果樹中發現上百種不同的miRNA。Phillips研究表明,植物miRNA廣泛參與生物脅迫和非生物脅迫等各類逆境脅迫的響應[33],逆境脅迫會使得植物改變某些 miRNA 的表達模式,發生上調或下調表達,完成對其靶基因表達模式的調控[34]。
葉片中對照和NaCl處理的Clean reads長度主要分布在20~24 nt,長度為24 nt的序列所占比例最多,這與擬南芥[35]、番茄[36]、藏紅花[37]等研究結果一樣。對差異靶基因進行分類注釋,NaCl處理6和48 h時,KEGG共同富集的通路有N-glycan生物合成,天冬酰胺連接的糖鏈(N-linked glycan,N-glycan)是內質網蛋白質量控制系統中一個重要的調節信號,劉傳發[38]對擬南芥的分析發現,高爾基體糖原剪切酶突變體MNS1/2催化的N-glycan剪切的缺失調控了底物蛋白N-糖基化蛋白RSW2的豐度從而影響了纖維素的合成,產生了鹽敏感的表型,MNS1/2介導的N-glycan的剪切可能調節底物蛋白對鹽脅迫的響應。
馮克偉[39]對耐鹽性野生二粒小麥研究表明,在150 mM鹽脅迫下,7個miRNAs在3和6 h時表達量降低,12和24 h表達量顯著上升,而在250 mM鹽處理下miRNA在多個時間點均受鹽脅迫抑制了表達。鹽脅迫下山楊(Populustremula)miR398在3~4 h內其表達受到誘導,在48 h后開始下降,至72 h又開始積累[40],Chen等[41]研究發現鹽脅迫下擬南芥中miR159、miR171等表達上調,而miR398的表達下調。Liu等[42]在擬南芥中鑒定到在鹽脅迫下miR396、mi R168和miR169等表達量升高,過表達miR397的擬南芥植株抗鹽性提高。Patade等[43]發現甘蔗受到短暫的鹽脅迫后miR159含量顯著提高,miR159在甘蔗抗鹽脅迫過程中發揮重要作用。Ding等[44]發現,玉米鹽處理后mi R159大量增加。Wu等[45]發現鹽脅迫下桑樹(Morusalba)與對照相比miR827和miR2111的表達則顯著下降。Sun等[46]研究大豆根尖分生組織發現miR390e、miR399a、參與了大豆根系耐鹽性的調控。miR171是最早發現的 miRNAs 家族成員之一。Fan等[47]研究表明,水稻miR171通過剪切GRAS家族基因OsHAM促進營養生長向生殖生長過渡以及根尖分生組織穩態形成;擬南芥miR171通過靶向GRAS家族基因Scarecrow-Like(SCL)調控擬南芥的葉綠素生物合成[48]。Curaba等[49]發現大麥過表達的miR171能夠激活miR156對其下游靶基因表達抑制,導致植物營養生長向生殖生長轉變。實驗通過microRNA測序及qRT-PCR驗證發現,mdm-miR168b、mdm-miR159c、mdm-miR827、mdm-miR390f、mdm-miR171i、mdm-miR399j基因在NaCl處理6和48 h時表達量存在差異,其中mdm-miR390f及其靶基因HF10976、mdm-miR171i及其靶基因HF33844、mdm-miR399j及其靶基因HF11095表達量呈負相關,這3個miRNA參與了新疆野蘋果對NaCl脅迫的響應過程,但是新疆野蘋果不同時間點miRNA差異表達的調控原因及miRNA響應NaCl脅迫作用方式還需要進一步的試驗驗證。
4 結 論
NaCl處理6 h時3個miRNA差異表達,其中2個上調表達,1個下調表達,miRNA對應的靶基因數目為64個。NaCl處理48 h時13個miRNA差異表達,其中4個上調表達,9個下調表達,miRNA對應的靶基因數目為108個。mdm-miR390f及其靶基因HF10976、mdm-miR171i及其靶基因HF33844、mdm-miR399j及其靶基因HF11095表達量呈負相關,3個miRNA參與了新疆野蘋果對NaCl脅迫的響應過程。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
四川文學(2021年4期)2021-07-22 07:11:54
科技傳播(2019年22期)2020-01-14 03:06:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
絲綢之路(2014年9期)2015-01-22 04:24:46
