低植酸作物遺傳改良途徑與磷資源高效利用
2021-10-24 09:40:00李家慶郭競選袁力行
植物營養與肥料學報 2021年9期
關鍵詞:途徑
蔣 鵬,李家慶,郭競選,趙 政,袁力行
(中國農業大學資源與環境學院,國家農業綠色發展研究院/植物-土壤相互作用教育部重點實驗室,北京 100193)
磷 (phosphorus,P) 是一種有限的、不可再生的礦質元素,在生命遺傳繁衍 (DNA和RNA)、能量轉運 (ATP) 等過程中發揮著重要作用[1]。磷是作物生長所必需的大量元素之一。作物根系主要吸收無機磷酸鹽 (phosphate,Pi)。由于土壤磷具有低溶解度、低遷移率和高固定率的特征,土壤溶液中的Pi濃度僅為1~10 μmol/L[2-3],而植物組織內的Pi濃度一般達到5~20 mmol/L,因此在農業生產中需要施用磷肥來維持作物生長。磷礦資源是磷肥生產的原料,全球探明儲量可達6.9 × 1010t,其大部分集中在摩洛哥(USGS,2020)[4]。我國磷礦資源儲量約有 3.2 × 109t[4],主要分布于云、貴、川、湘、鄂5省[5],磷礦石P2O5的平均品位為16.85%[5-6],存在資源集中、富礦及易選礦少等問題[7]。然而,農業磷資源的過量投入卻導致了全球農田磷盈余,從1961年的2.3 × 106t增長到2013年的1.08 × 107t,僅有22%的磷投入最終被人類消費[8]。大部分盈余磷進入環境引起了水體富營養化[9-10]、土壤重金屬污染等問題[11-12]。為此,磷資源的可持續利用及其對糧食安全和環境安全的影響已經引起世界各國的重視[13]。
為了解決磷的資源與環境問題,必須從整個農業系統磷流鏈條的角度創新磷資源可持續性管理[14]。其中,提高作物磷效率 (P efficiency,PE) 尤為重要,包括作物從土壤中獲取磷的效率 (PupE,P uptake efficiency) 和體內磷的利用效率 (P utilization efficiency,PutE)[14-15]。植物根系進化出多種形態和生理學策略來高效獲取土壤磷。磷高效吸收的理想根構型 (root system architecture,RSA) 應具有較淺的根基部生長角度、密集的側根分支和較長的根毛,從而通過增加淺層土壤的根表面積來提高PupE[16-17]。植物根系還可以通過增加質子、有機酸陰離子和酸性磷酸酶分泌來提高土壤中難溶磷的利用率[18]。為了提高PutE,植物可將衰老葉片組織中豐富的線粒體與葉綠體中的核酸 (DNA和RNA) 分解進行循環利用[19],活化與再利用液泡中存儲的磷,以及減少磷向籽粒的分配等[20-21]。此外,植物磷的吸收利用通常伴隨著與氮、鈣等元素的相互作用[22-24],調控植物對其它元素的吸收利用也能間接提高磷效率。
作物體內60%~85%的磷儲存在成熟的籽粒中[25-26],每年作物收獲帶走的磷約占全球農田磷肥施用量的85%[27],而動物或人類消費的磷僅有約10%能被再循環利用[28]。植酸 (肌醇-1,2,3,4,5,6-己磷酸,InsP6) 是磷在籽粒中的主要儲存形式,占籽粒總磷的65%~80%[29-30]。植酸能夠螯合Fe2+、Zn2+等元素形成植酸鹽[31]。籽粒萌發過程中內源植酸酶被激活,降解植酸鹽來釋放肌醇、磷和其他礦質陽離子用于支撐幼苗生長[32]。然而,包括人類在內的非反芻動物均無法消化植酸鹽來獲取其螯合的Fe2+、Zn2+等有益微量元素[33-34]。這些植酸鹽隨動物排泄物排出體外,進入水體后加劇富營養化而造成環境污染[35]。畜牧業中通常向動物飼料中添加磷補充劑[36]或外源植酸酶[35]來滿足動物營養需求,但磷補充劑的添加無法從根本上解決高磷排泄物對環境的危害。添加植酸酶雖然能夠降解籽粒中富含的植酸,但費用高昂。
農業系統的磷流動鏈條包括磷礦開采加工、磷肥生產、土壤磷過程、作物生產、畜禽養殖、人類消費,及最終向環境的磷輸入等多個過程,將礦石中的磷逐步轉化為土壤磷、植物磷和動物磷,以滿足人類的消費需求。開發籽粒低植酸 (low grain phytate,LGP) 作物,可以通過降低籽粒植酸含量來解決植酸磷引起的營養問題,降低作物磷需求,減少磷向環境的排放,從而提升磷在農業系統中的循環利用效率,顯示出廣闊的應用前景和商業價值[37]。籽粒低植酸作物品種有兩種類型:一類品種的籽粒總磷基本不變,但植酸含量低,因此,大部分籽粒磷可以被動物有效利用而導致“廢物磷”排放減少,并且籽粒中有益元素的生物有效性也較高,營養品質好,降低了動物飼料中磷補充劑等資源的額外投入;另一類品種的籽粒植酸和總磷含量均低,作物吸收的磷主要積累在秸稈中,這類品種既具有籽粒低植酸的優點,又減少了籽粒收獲帶走的磷量,積累在秸稈中的磷還可通過秸稈還田來實現磷在農田中的循環利用,降低了作物生產的磷資源需求 (圖1)。本文系統綜述作物籽粒中磷的來源、籽粒低植酸品種的遺傳改良途徑,剖析現有研究存在的不足,并對未來籽粒低植酸品種培育與應用進行展望。
1 植物籽粒磷的來源
植物體內磷以無機磷酸鹽 (Pi) 和有機磷酸酯兩種形式存在。籽粒中Pi在細胞質基質或內質網中合成植酸,并最終儲存在蛋白質儲存液泡 (protein storage vacuole) 中[38]。盡管籽粒植酸含量在不同作物品種中存在差異,但它與籽粒總磷之間的相關性通常 ≥ 95%[39]。磷在籽粒中的積累主要通過3個步驟完成:Pi從源器官運輸到籽粒、籽粒利用Pi合成植酸以及合成植酸的轉運與儲存。
Pi運輸到籽粒有兩個途徑,包括從葉片等營養器官中活化的Pi通過韌皮部運輸到籽粒,和從根系吸收的Pi通過木質部運輸到籽粒。因此,土壤Pi的供應和植物組織的Pi濃度能夠顯著影響Pi從源器官到籽粒的運輸過程[40]。目前在模式植物擬南芥中鑒定到一些關鍵基因,如AtPHT1;5[41]基因可能參與Pi向籽粒的韌皮部運輸,AtSPDT[42]基因則通過介導Pi從木質部向韌皮部的橫向轉運來影響Pi向籽粒的分配;在水稻中也鑒定到參與木質部Pi在莖節的橫向轉運基因SPDT[21]、OsPHO1;1和OsPHO1;2[43];另外,OsPHT1;8[44]基因可能參與穗軸中Pi向籽粒的運輸。在籽粒內部,OsPHT1;4[45]和OsPHT1;8基因參與了Pi向胚的轉運,OsSULTR3;3[46]基因則可能在亞細胞水平介導細胞質中Pi向內質網的轉運以合成植酸 (表1和圖2)。
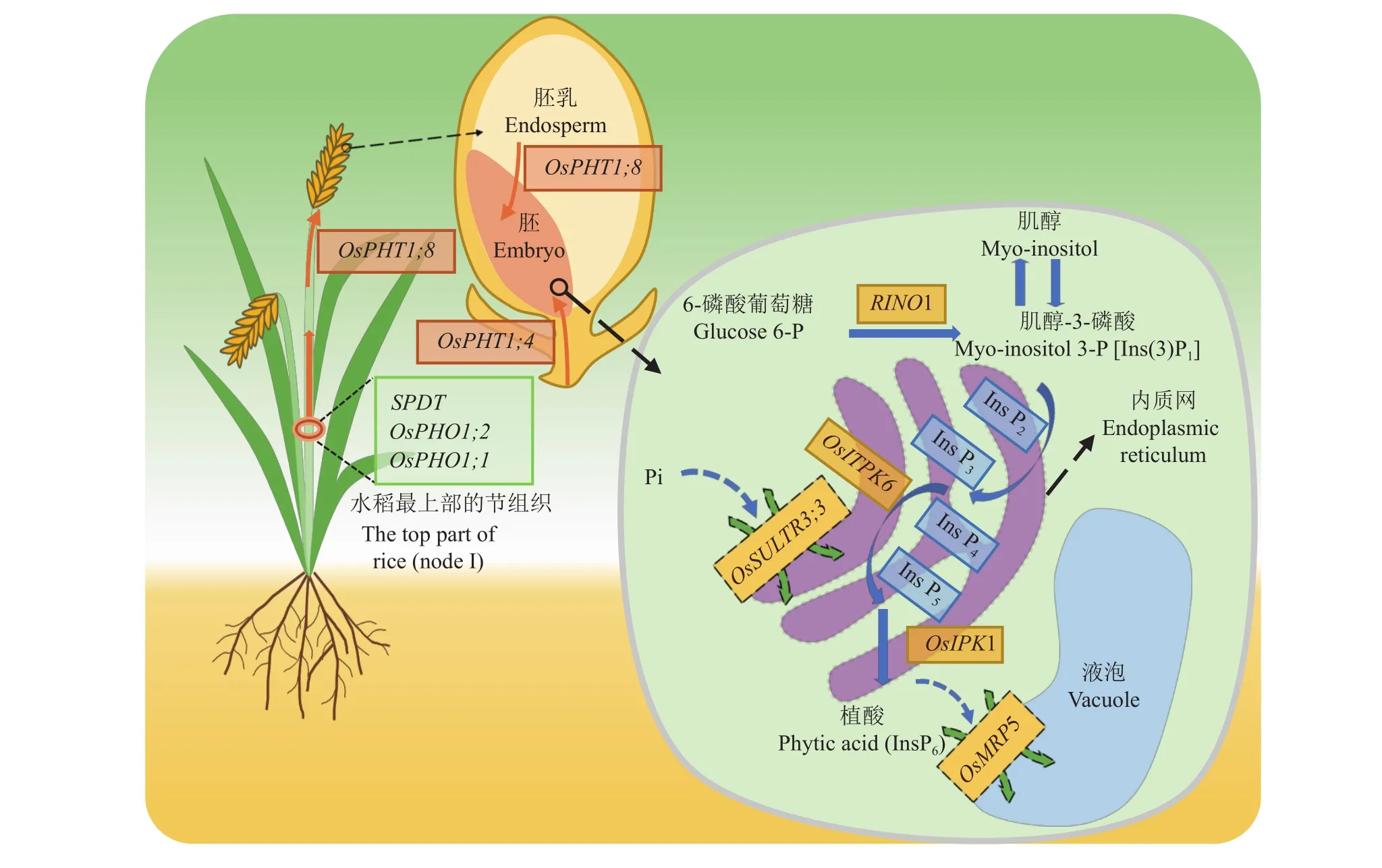
圖2 影響籽粒植酸累積的關鍵基因 (以水稻為例)Fig. 2 Key genes involved in grain phytate accumulation (take rice as an example)
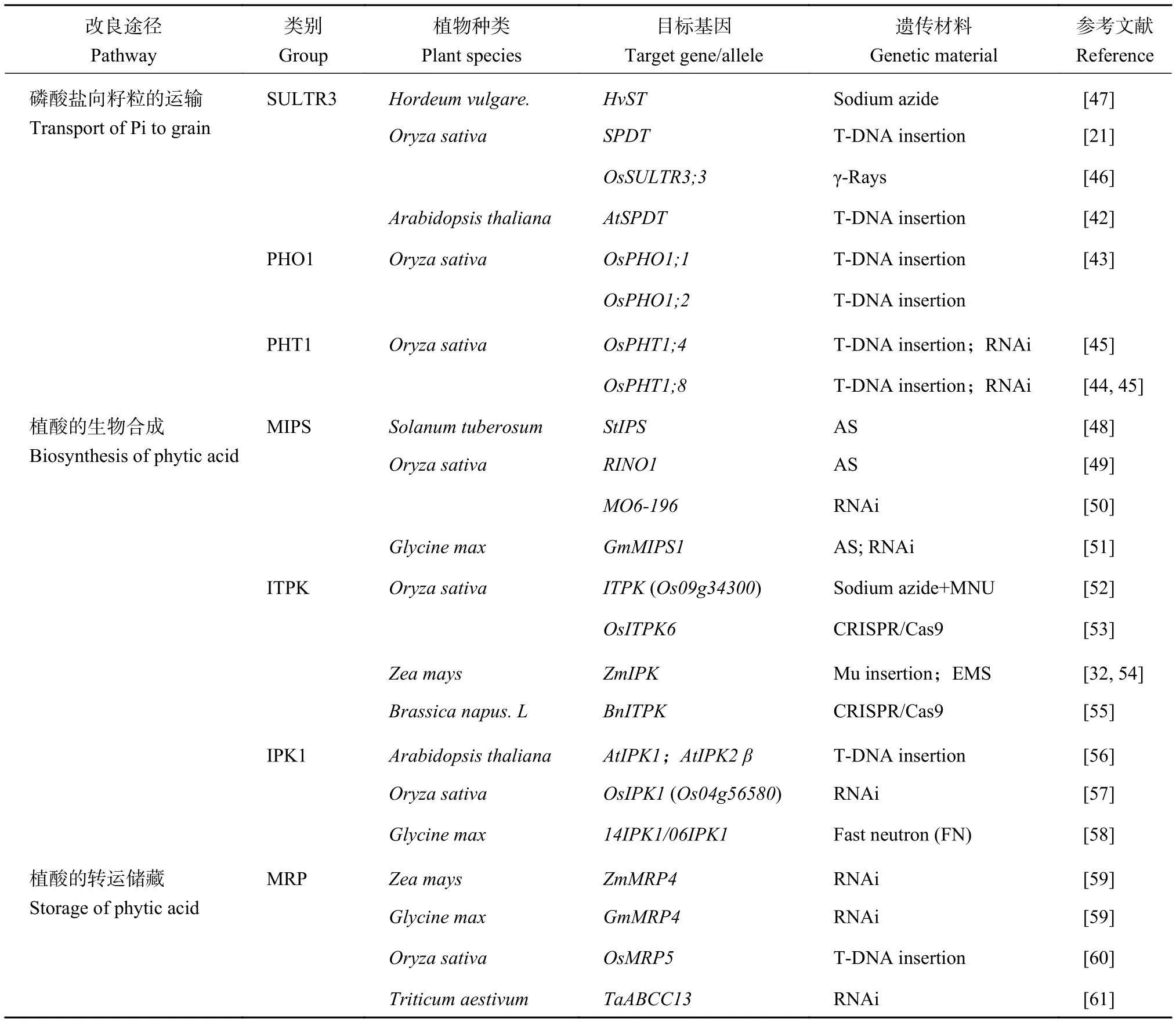
表1 籽粒低植酸品種的遺傳改良途徑Table 1 Pathways of genetic improvement for low grain phytate varieties
Pi運輸到籽粒后,在細胞質基質或內質網中合成植酸,包括非脂質依賴性途徑和脂質依賴性途徑兩種不同的途徑,均以肌醇作為反應底物。6-磷酸葡萄糖 (glucose 6-phosphate) 首先在肌醇-3-磷酸合酶(myo-inositol-3-phosphate synthase,MIPS) 作用下轉化成肌醇-3-磷酸 [Ins (3) P1],肌醇-3-磷酸水解酶[Ins(3) P1- monophosphatase,IMP] 進一步催化肌醇-3-磷酸生成肌醇。植物組織中常見的脂質依賴性途徑是以磷脂酰肌醇 (phosphatidylinositol,PtdIns) 為前體,由肌醇在磷脂酰肌醇合酶 (phosphatidylinositol synthase) 的催化下合成,磷脂酰肌醇在磷脂酶 C(phospholipase C,PLC) 水解作用下生成肌醇-1,4,5-三磷酸[myo-inositol-1,4,5-trisphosphate,Ins (1,4,5)P3],經過多步磷酸化生成植酸。而非脂質依賴性途徑在植物籽粒植酸合成中占主導地位,該途徑中肌醇在肌醇激酶 (myo-inositol kinase,MIK) 的催化下生成肌醇-3-磷酸,然后再經過多個激酶的順序磷酸化形成植酸,涉及MIPS、ITPK、IPK1等關鍵酶。植酸生物合成途徑以及相關酶的作用機制非常復雜,還需要深入研究[62]。
合成后的植酸在籽粒中進一步轉運并儲存。不同谷物中的植酸含量和分布不同,如玉米中植酸主要儲存于胚和盾片中,小麥、大麥和水稻中80%的植酸儲存于糊粉層和麩皮中,只有小部分植酸儲存在胚中[63];豆科作物中95%以上的籽粒植酸在子葉中積累[64];擬南芥中植酸則主要儲存在胚中[65]。在水稻中,籽粒發育過程中胚的植酸含量始終較低,但調控植酸合成最后一步的OsIPK1基因在胚和糊粉層中表達最高,因此推測胚中合成的植酸立即被運輸到糊粉層中,或者植酸只在糊粉層和亞糊粉層中被合成[66]。目前鑒定到ABC轉運蛋白 (ATP-binding cassette transporter) 家族成員中的MRP (multidrugresistance-associated protein) 蛋白可能參與了籽粒中植酸的轉運與儲存過程[67]。
2 籽粒低植酸品種的遺傳改良途徑
降低Pi從源器官向籽粒的分配,阻斷植酸的生物合成或轉運儲藏,都能顯著降低籽粒的植酸含量,是籽粒低植酸品種遺傳改良的主要途徑。低植酸突變體材料是籽粒低植酸品種遺傳改良的種質基礎。研究早期,利用物理、化學誘變或插入突變等方法構建作物突變體庫,科學家直接篩選獲得一些低植酸突變體材料,并利用這些遺傳資源分離克隆到控制籽粒植酸累積的關鍵基因。另外,隨著籽粒植酸磷積累途徑分子機制的解析,通過反義、RNAi干擾等技術下調關鍵基因表達,或采用T-DNA插入、CRISPR/Cas9靶向敲除基因,也能創制相應的低植酸突變體。利用上述正向或反向遺傳學手段,在許多作物種類中都取得了很好研究進展,具體匯總在表1中。
2.1 阻斷Pi轉運途徑—減少無機磷向籽粒的運輸
減少Pi從源器官向籽粒的運輸與分配是降低籽粒植酸含量最直接的途徑,傾向于產生籽粒植酸低且總磷也低的品種。目前,已經在SULTR3、PHO1、PHT1家族中克隆出參與Pi向籽粒運輸的關鍵基因成員。它們在植物體內的時空表達模式并不相同,相關低植酸突變體的農藝性狀表現也有很大差異。
2.1.1 SULTR 3 (硫酸鹽轉運蛋白,sulfate transporter) 近年來,在水稻和擬南芥中均鑒定到SULTR3;4基因參與Pi由木質部向籽粒的運輸過程。SULTR3;4基因編碼的SPDT (SULTR-like phosphorus distribution transporter) 蛋白定位于質膜上,介導H+/Pi的同向轉運,而不參與轉運硫。Yamaji等[21]研究發現,水稻籽粒灌漿期間,SPDT基因在旗葉與穗部相連的莖節(node I) 處大量表達,受低磷脅迫誘導。它介導了根系吸收的Pi從莖節木質部向韌皮部轉移,尤其是Pi從擴大維管束 (enlarged vascular bundles,EVBs)向與穗部相連的分散維管束 (diffuse vascular bundles,DVBs) 轉移,從而將Pi優先分配到籽粒。Ding等[42]在擬南芥中也鑒定到類似功能的AtSPDT基因,介導了Pi從木質部經維管形成層向韌皮部的橫向轉運,進而實現Pi向新生葉及籽粒的優先分配。由于擬南芥的維管系統不同于水稻,根系吸收的Pi通過蒸騰作用向上運輸到木質部后,是由位于不同器官分枝點的木質部薄壁細胞中的AtSPDT轉運蛋白將其從木質部卸載,再經由維管形成層和伴胞/韌皮部薄壁細胞中的AtSPDT進一步向韌皮部轉運,最后運輸到籽粒。
在野生型水稻中,地上部總磷的64.5%向籽粒分配,而在spdt突變體中則降低到42.5%~44.0%。由于整株總磷含量沒有顯著改變,突變體秸稈的磷含量由35%增至57%。因此,spdt籽粒磷含量下降主要是抑制SPDT介導Pi向籽粒分配所造成的。籽粒總磷降低約20%,但并不影響突變體的農藝性狀及產量,而秸稈中增加的磷累積可通過秸稈還田來循環利用,因此能節約20%的磷肥投入。突變體的籽粒植酸濃度比野生型降低25%~32%,但不影響籽粒發芽率及其早期萌發,Fe2+、Zn2+的生物有效性得到增強,還降低了植酸磷向環境排放的風險。通過抑制SPDT基因介導的Pi向籽粒分配的過程,可顯著降低籽粒植酸磷含量。不影響作物產量,減少了作物磷需求,提高了籽粒品質,是遺傳改良的最好靶標。
水稻中還有另外一個SULTR3家族成員OsSULTR3;3,其突變體ossultr3;3的籽粒植酸濃度顯著降低35.2%~43.9%,總磷濃度相應下降18.9%~27.5%[46]。OsSULTR3;3基因在發育籽粒的維管束中表達,其編碼蛋白定位于內質網上。異源表達體系沒有檢測到OsSULTR3;3蛋白轉運肌醇等植酸前體、磷酸鹽或硫酸鹽的活性,但不排除在植物中具有相應的轉運功能。OsSULTR3;3基因只對植酸合成后期步驟有影響,基因突變會導致編碼IPK1、ITPK激酶的基因表達水平改變。鑒于InsP3到InsP6的轉化發生在內質網上[65],OsSULTR3;3基因可能參與Pi向內質網的轉運,從而為植酸合成提供原料。此外,突變體總硫、硫酸鹽以及糖醇、生物胺和游離脂肪酸等營養相關代謝物的濃度上升,說明該基因還可能參與植物組織內部磷酸鹽和硫酸鹽穩態調節及信號傳導過程。Zhou等[68]發現ossultr3;3突變體與商業栽培種的雜交后代,可以穩定觀察到籽粒植酸降低和代謝物增加的表型。因此,遺傳操作OsSULTR3;3既能降低籽粒植酸,還能增強籽粒營養,也是有效的改良途徑之一。
大麥lpa1-1突變體作為最早進行商業化育種的籽粒低總磷類型的LGP品種[69-70],籽粒植酸含量降低約50%,總磷降低15%~20%[71],并且能夠特異性降低胚乳30%的總磷[72],具有良好的田間表現[73]。有意思的是,導致lpa1-1突變體表型的大麥HvST也編碼了一個SULTR3家族成員,有可能參與植酸合成原料Pi的轉運,但具體機制還有待研究[47,74]。
2.1.2 PHO1 (磷轉運蛋白,phosphate transporter)研究發現水稻OsPHO1;1和OsPHO1;2基因也參與Pi向籽粒的分配過程。它們編碼的轉運蛋白同屬于PHO1家族,介導Pi的跨質膜流入過程[43]。在成熟期,OsPHO1;1和OsPHO1;2基因在莖節 (node I) 處有較高的表達,其中OsPHO1;1基因主要在DVBs的韌皮部表達,而OsPHO1;2基因主要在EVBs的木質部表達。OsPHO1;2負責將Pi從EVBs的木質部中卸載,而OsPHO1;1則將Pi裝載到DVBs的韌皮部后再運輸到籽粒。敲除OsPHO1;2基因會減少Pi向籽粒的分配,與野生型相比籽粒磷濃度降低約44%~50%,但并不影響其他礦質元素的分配。與敲除OsPHO1;2基因相比,敲除OsPHO1;1基因對水稻Pi向籽粒分配的影響較小。此外,OsPHO1;1基因在穎果的胚珠維管束和內珠皮的外層,OsPHO1;2基因在穎果的胚珠維管束和珠心表皮也有表達,推測它們可能也參與了Pi從母體組織向籽粒發育的子代組織的傳遞。這一假設也在擬南芥的PHO1同源基因研究中得到驗證[75]。然而,ospho1;1和ospho1;2突變體與野生型的株高和穗數相比均有下降,籽粒明顯變小且萌發延遲,ospho1;2突變體的千粒重僅為野生型的52%~58%。因此,對PHO1基因的遺傳操作會導致作物產生不良的農藝表現,用于培育低植酸品種的價值不大。
2.1.3 PHT1 (磷轉運蛋白,phosphate transporter)研究發現磷轉運蛋白PHT1家族成員參與了衰老組織中活化出的Pi向籽粒的再轉移過程。AtPHT1;5基因在擬南芥衰老葉片的維管束韌皮部中高度表達,負責Pi從衰老葉片到庫器官的轉移。過表達AtPHT1;5可導致轉基因株系的籽粒Pi濃度提高兩倍[41]。OsPHT1;4[45]和OsPHT1;8[44]基因在水稻籽粒發育過程中也有高表達,但二者功能并不冗余。OsPHT1;4基因在胚中的表達量是胚乳和穗軸的10倍[45],可能介導Pi從其他組織向籽粒胚的轉運[76]。OsPHT1;4基因功能缺失會導致籽粒中22%~32%的植酸濃度和胚中21%~24%的總磷含量下降,胚乳總磷含量不受影響。參與植酸合成的MIPS合酶和IPK1激酶基因的表達也顯著降低,并造成胚的縮小。過表達OsPHT1;4基因則導致胚和胚乳中總磷和植酸濃度增加,植酸合成相關酶的基因表達也被上調。OsPHT1;4基因的功能缺失會導致穗莖稈較弱,結實率比野生型降低50%~58%,產量和千粒重也相應減少,籽粒萌發和根系伸長均受到抑制[45]。OsPHT1;8基因可能參與Pi從穗軸向籽粒的運輸,以及從胚乳向胚的運輸。缺失OsPHT1;8基因導致突變體穗軸中總磷含量增加30%,灌漿期籽粒中總磷降低30%,結實率降低[44]。OsPHT1;4和OsPHT1;8基因在水稻體內有著復雜的生物學功能,基因缺失突變體有不良農藝表現。然而,選擇性沉默胚乳中的OsPHT1;8基因則導致胚中總磷含量下降40%~50%,但不影響胚乳中的磷含量,對千粒重和結實率影響也較小[77],成為一種可能的籽粒低植酸性狀的改良途徑。
2.2 阻斷植酸合成途徑—控制Pi合成植酸
作物籽粒中的植酸主要是通過非脂質依賴性途徑合成,涉及MIPS、ITPK、IPK1等關鍵酶。降低這些關鍵酶的表達及活性,傾向于產生籽粒低植酸、總磷不變的作物品種。
2.2.1 MIPS (肌醇-3-磷酸合酶,myo-inositol-3-phosphate synthase) 植酸合成的首要步驟是MIPS催化6-磷酸葡萄糖生成Ins (3) P1,該反應也是生物體內肌醇的唯一從頭來源[78]。不同植物基因組中的MIPS基因數量不同,如水稻有兩個MIPS基因,大豆有4個MIPS基因,它們在籽粒發育階段各有一個MIPS基因特異性高表達[47,79-82]。MIPS基因最早由Dean-Johnson等[83]通過遺傳互補從釀酒酵母中克隆。Kuwano等[84]通過反義RNA來降低編碼MIPS合酶的RINO1基因表達,獲得籽粒植酸含量下降、但總磷含量沒有降低的水稻突變體。雖然敲除MIPS基因可以顯著降低籽粒植酸含量,但通常會對籽粒萌發、幼苗生長和產量造成不良影響。Keller等[48]利用反義RNA抑制馬鈴薯中StIPS基因的表達水平至野生型水平的20%以下,籽粒肌醇含量明顯降低,但會導致葉片形態改變、早熟衰老及整體塊莖產量下降。
有研究發現對籽粒MIPS基因進行特異性沉默似乎可以避免敲除基因帶來的相關缺陷。Kuwano等[49]采用籽粒特異性啟動子oleosin18,反義下調了水稻RINO1基因表達,籽粒植酸含量減少了68%,但對籽粒干重、發芽或植株生長沒有產生負面影響,Ali等[50]利用RNAi技術沉默水稻RINO1基因表達也取得了類似的效果。在大豆中使用籽粒特異性啟動子vicilin下調GmMIPS1基因的表達,也不會影響其他組織的肌醇代謝[85],Kumar等[51]利用vicilin靶向破壞大豆籽粒中GmMIPS1基因的表達,突變株系的籽粒中分別觀察到38.75%和41.34%植酸含量的降低,礦質元素Fe2+、Ca2+的含量顯著增加,表明特異性沉默MIPS基因來實現籽粒低植酸性狀遺傳改良切實可行。
2.2.2 ITPK (肌醇三/四磷酸激酶,inositol tris/tetrakisphosphate kinase) ITPK激酶屬于ATPGRASP折疊蛋白家族 (ATP-grasp fold protein)。在肌醇形成與代謝中間過程中,ITPK激酶催化的肌醇磷酸鹽的持續磷酸化過程研究最多。通過可逆賴氨酸乙酰化可以調控ITPK1激酶,進而抑制肌醇的更高磷酸化,降低細胞中的植酸含量[86]。目前在擬南芥[87-88]和大豆[89]中分別鑒定到4種不同的ITPK激酶,其中大豆GmITPK3在籽粒發育早期有較高表達。在水稻中鑒定到6個ITPK激酶[62],其中OsITP5/6K-4和OsITP5/6K-6基因在籽粒胚中特異性表達。盡管水稻OsITP5/6K-6基因的突變可以有效抑制水稻籽粒的植酸合成,卻會顯著影響植株的生長和繁殖[53]。Kim等[52]篩選水稻TILLING (targeting of induced local lesions IN genomes) 庫鑒定到4個低植酸突變體,其中兩個目標基因屬于ITPK基因家族,其突變導致籽粒中植酸含量分別減少46%和68%,其中減少46%植酸含量的突變株系的農藝性狀無顯著變化。目前在玉米中只鑒定到一個編碼ITPK激酶的基因ZmIPK,且在胚中特異性表達,該基因的突變可以降低玉米籽粒30%~50%的植酸含量且總磷不變,籽粒干重有部分下降但不會影響功能[32,54]。最近,Sashidhar等[55]在多倍體油菜中尋找到15個ITPK同源基因,通過CRISPR/Cas9技術獲得了兩個ITPK基因三重突變體bnitpk1。其籽粒植酸含量顯著降低27.2%~35.3%,同時Pi含量增加,總磷基本保持不變,并沒有表現出產量和種子活力等農藝性狀的缺陷。因此,在不同作物中敲除關鍵酶ITPK的編碼基因可以降低籽粒植酸含量,但對農藝性狀的影響在不同作物種類中存在較大差異,該途徑的應用還需更深入的研究。
2.2.3 IPK1 (肌醇-五磷酸二激酶,inositolpentakisphosphate 2-kinase) 植酸生物合成的最終步驟是IPK1激酶催化Ins (1,3,4,5,6) P5轉化為InsP6。Sun等[90]在玉米中鑒定到ZmIPK1A和ZmIPK1B為兩個相似的旁系同源基因,ZmIPK1B基因在根中表達,而ZmIPK1A基因在幼穗、籽粒的胚乳和胚中等部位均有表達。水稻的IPK1基因在籽粒發育早期的糊粉層中表達最高[62]。擬南芥中至少有兩個IPK1基因表達,其中AtIPK1基因在發育階段的花蕾、角果等部位表達[91]。AtIPK1和AtIPK2β基因雙突變體的籽粒植酸含量降低83%,Pi含量增加10倍,且籽粒產量不受影響[56]。
擬南芥中的AtIPK1基因的突變雖然可以降低籽粒植酸含量,但會導致植物體內磷酸鹽穩態的紊亂,導致磷酸鹽的過度積累[92]。植酸參與細胞核的mRNA輸出過程,AtIPK1基因突變會導致mRNA輸出不足,Lee等[93]發現Gle1蛋白是調控植酸在植株生長和繁殖中功能的重要因子,gle1基因突變的擬南芥表現出對植酸的敏感性增強,可以彌補AtIPK1基因突變的缺陷,顯著提高突變體的營養生長和籽粒產量,為從遺傳操作植酸信號轉導途徑培育低植酸作物品種提供了可能。Ali等[57]使用啟動子oleosin18特異性沉默籽粒IPK1基因的表達,突變體水稻的籽粒植酸含量顯著降低,Pi含量增加,胚乳的鐵含量為野生型的1.8倍,并且對籽粒萌發等農藝性狀沒有產生負面影響。Vincent等[58]通過挖掘作物優良的等位變異,對大豆6號染色體與14號染色體的IPK1基因進行基因型和籽粒磷表型間的關聯分析,培育出兩個位點上均有突變的籽粒低植酸品種,籽粒萌發和田間出苗率等性狀均無顯著降低,并表現出極低的植酸水平與適宜的Pi水平。圍繞植酸信號轉導過程,特異性沉默籽粒IPK1基因及挖掘優良等位變異等為遺傳改良籽粒低植酸品種提供了可行途徑。
2.3 阻斷植酸轉運途徑—控制籽粒的植酸含量
植酸在細胞質基質或內質網中通過肌醇的順序磷酸化合成后,在由ABC型的MRP蛋白介導下跨膜運輸與區室化,并最終儲存在液泡中。也有研究認為,MRP蛋白是通過調控植酸合成原料Pi的供應來參與調節植酸水平[67,94-95]。對植酸轉運途徑進行阻斷,植酸將無法轉運到液泡內存儲,因而在細胞質被磷酸酶降解,從而使植酸含量減少、Pi含量增加,傾向于產生籽粒低植酸、總磷不變的品種。Shi等[59]利用籽粒特異性啟動子globoulin-1靶向沉默玉米胚中編碼MRP4蛋白的基因,突變體籽粒的植酸含量下降32%~75%,總磷基本保持不變,籽粒發育未受影響。在大豆中對同源基因進行沉默也得到相似效果。
然而,T-DNA插入缺失水稻OsMRP5基因,雖然顯著降低了籽粒的植酸含量,但是子代出現純合致死現象[66]。大豆中MRP蛋白的功能缺失也可能會對作物生長發育造成不良影響。Neelam等[96]對低植酸大豆品系 (攜帶一個MIPS和兩個MRP基因突變)與正常品系的RNA測序數據進行了差異表達和功能富集分析,發現突變體籽粒的細胞壁合成相關基因提前表達,而光合作用相關基因在發育后期表達降低,可能會對籽粒的產量和發育造成負面影響。Bhati等[61]利用RNAi技術沉默小麥TaABCC13基因的表達,籽粒的植酸含量雖然降低22%~34%,但小麥的穗發育不良、籽粒灌漿減少,并且TaABCC13基因參與小麥的側根發生和重金屬解毒。MRP基因在植物體內有著復雜的生物學功能,還不能簡單通過敲除或沉默該基因的表達來進行籽粒低植酸性狀的遺傳改良。
3 展望
培育籽粒低植酸品種是解決農業磷資源、環境及動物/人類營養問題的關鍵。較外源植酸酶添加等手段[97],它仍是解決籽粒植酸磷問題的優選方案。本文綜述提出的調控植酸磷積累關鍵基因可以作為未來育種的潛在靶標。目前亟待解決的問題是,籽粒低植酸表現會伴隨產量下降、種子萌發率低等不良農藝性狀,尤其是遺傳操作植酸合成途徑相關基因。未來可從3個方向著手,突破制約籽粒低植酸品種培育及應用的瓶頸。
3.1 特異性修飾籽粒中關鍵基因的表達
植酸和其前體肌醇、肌醇磷酸鹽及衍生物肌醇焦磷酸鹽 (InsP7,InsP8) 行使著重要功能,如信號轉導、生物和非生物脅迫的響應、膜運輸及光形態建成等[98],針對肌醇代謝途徑的擾動都可能會影響籽粒和植株的發育[99]。未來的研究應該深入關鍵基因對植物生長發育具體的調控功能,通過對籽粒中關鍵基因的時空表達進行定向修飾,而不影響植物其他組織中的肌醇代謝,尋找低植酸性狀與植物正常發育的平衡點。如前文所述,利用水稻和大豆的籽粒特異性啟動子oleosin18[49]和vicilin[51]沉默植酸合成途徑關鍵酶基因的表達,既能調控植酸的合成又不造成籽粒的發育缺陷。
3.2 發掘優良等位變異,結合新型基因編輯技術予以應用
利用遺傳多樣性種質資源,高通量分析籽粒植酸磷累積表型,找到能夠平衡低植酸和產量等重要農藝性狀的遺傳材料。隨后利用全基因組關聯或候選基因關聯等策略,挖掘作物基因組內優良的等位變異,這在低植酸大豆育種中已經取得良好進展[64]。通過傳統育種如雜交,可將低植酸性狀與籽粒高萌發率等農藝性狀相結合[100],能顯著提高田間出苗率[101],但是存在后代性狀分離的缺陷。Rosso等[102]利用分子標記輔助育種開發了針對大豆GmMIPS1基因的良好遺傳標記,利用該技術可以將優良等位變異導入到高產品種中予以應用。近年來,以CRISPR/Cas9為代表的新型基因編輯技術發展迅猛,能夠實現對特定DNA片段的敲除、插入和替換,具有育種周期短、編輯效率高等優勢[103]。在新型基因編輯技術的幫助下,可以精準引入關鍵基因及優良等位變異,大大加快籽粒低植酸品種的育種。
3.3 針對籽粒低植酸品種的磷營養管理
一般認為,維持作物籽粒的正常萌發和幼苗的早期生長只需要籽粒磷濃度高于1 mg/g即可,剩余的大部分籽粒磷并不是種子萌發和幼苗發育所必需[104]。但相較于野生型,籽粒低植酸品種對外源磷營養的供應更為敏感[105],是造成現存的低植酸品種籽粒的萌發和活力比野生型低的重要原因。因此,可以通過田間磷營養管理來提高低植酸品種籽粒的萌發率,如播種時增施種子啟動磷肥或種子包膜,為籽粒低植酸品種進行針對性的田間農藝管理,這也是未來的研究重點。
猜你喜歡
語數外學習·高中版中旬(2023年2期)2023-05-10 13:26:53
語數外學習·高中版中旬(2022年5期)2022-07-13 20:47:51
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中學生百科·大語文(2017年10期)2017-11-04 06:56:38
中國衛生(2016年3期)2016-11-12 13:23:26
公民與法治(2016年22期)2016-05-17 04:20:13
中國衛生(2014年12期)2014-11-12 13:12:52
癌變·畸變·突變(2014年6期)2014-02-27 06:15:03
