卵形鯧鲹 SST1基因的克隆與組織表達
2021-10-24 05:38:38馮鵬霏潘傳燕余艷玲杜雪松張永德羅洪林
西北農業學報 2021年8期
關鍵詞:分析
黃 姻,馮鵬霏,潘傳燕,余艷玲,武 霞,杜雪松,張永德,羅洪林
(1.廣西壯族自治區水產科學研究院 廣西水產遺傳育種與健康養殖重點實驗室,南寧 530021;2.廣西師范大學 生命科學學院,廣西桂林 541006)
生長激素(Growth hormone,GH)是脊椎動物主要的生長調節因子,受多種激素的調控,通過其受體GHR作用于靶組織,或通過刺激肝臟產生胰島素樣生長因子(Insulin like growth factor,IGF)間接發揮作用[1]。在控制GH釋放的眾多激素中,生長抑素(Somatostatin,SS)是最直接有效的抑制劑[2]。SS是一個多肽激素家族,不僅可以直接抑制GH和IGF-1的合成與分泌,還可以通過降低對GH與IGF-1結合的敏感性,并以垂體外的方式影響GH-IGF-1系統[3-4],在動物的生長、發育和代謝等許多方面發揮重要的作用[5-6]。SS主要通過與靶細胞表面的特異性受體(Somatostatin receptor,SST)結合而發揮抑制GH分泌的作用[7]。SST是一種跨膜受體,屬于G蛋白偶聯受體(G-protein-coupled receptors,GPCRs)超家族,在動物體內已經發現7個SST(SST1~SST7),但目前在硬骨魚中尚未鑒定到SST4[8-9]。SST分布在機體多個系統和組織器官中,包括中樞神經系統、腸道、垂體、腎臟、甲狀腺、肺、免疫細胞和各種癌細胞[10-11]。每個SST都具有亞型選擇性、組織特異性和種屬特異性的分布模式,參與細胞內不同信號傳導的激活過程。研究表明,SST1可影響體質量并導致生長遲緩[12],還可以影響形態性狀的發育,包括體長、體高和胸圍等[13]。
卵形鯧鲹(Trachinotusovatus)是中國名貴的海水養殖魚類,也是適合中國近、遠海集約化養殖的重要品種之一,它具有生長快、適應性強等優良的生物學特性,其人工養殖近年來在中國南部沿海地區得到迅速發展[14],已成為中國農業部推薦的9種海水養殖品種之一。然而,由于育種工作的滯后,導致卵形鯧鲹出現明顯的種質退化現象,種群遺傳多樣性面臨較高的丟失風險[15],因此,開展卵形鯧鲹生長發育相關功能基因的研究,對促進卵形鯧鲹種質資源的開發利用及其產業的健康持續發展意義重大。SST1基因的表達與動物生長發育和形體性狀顯著相關,可作為研究生長相關的候選基因之一。本研究克隆卵形鯧鲹的SST1基因,并對其所表達的受體蛋白進行生物信息學分析,預測其可能的生物學功能,采用熒光定量PCR的方法對SST1基因在卵形鯧鲹不同胚胎發育時期及其幼魚不同組織中的分布和表達進行定量分析,旨在為進一步研究卵形鯧鲹SST1及SS基因在其生長發育過程中的作用奠定基礎。
1 材料與方法
1.1 材 料
卵形鯧鲹取自深圳市南澳鎮大鵬灣海域人工養殖群體,對卵形鯧鲹進行人工繁育[16],采集其胚胎發育的受精卵期、4細胞期、16細胞期、多細胞期、高囊胚期、原腸早期、原腸中期、原腸末期、胚體形成期、眼囊期、耳囊期、心臟跳動期、晶體出現期、初孵仔期共14個不同時期組織樣品各50~100 mg,每個時期3個生物學重復。另取雌雄各3尾,體質量為523~586 g的卵形鯧鲹幼魚,經MS222麻醉致死后,分別剖取其背鰭、皮膚、肌肉、眼睛、腮、腦、心臟、肝臟、脾臟、頭腎、精巢、卵巢、胃、腸共14個組織樣品各50~100 mg,每個組織雌雄各3個生物學重復。樣品經液氮速凍后轉入-80 ℃超低溫冰箱保存,用于SST1組織表達研究。
1.2 方 法
1.2.1SST1基因的生物信息學分析 登錄BioProjec(https://www.ncbi.nlm.nih.gov/bioproject),從卵形鯧鲹的基因組(登錄號:PRJNA574895)中找到SST1基因序列(ID:EVM0001630),利用NCBI BLAST對卵形鯧鲹與其他物種的SST1基因序列(表1)進行同源比對分析,下載與卵形鯧鲹相近的18種魚類的SST1基因序列,采用Mega X軟件對卵形鯧鲹與這18種近緣魚類的SST1基因序列進行多重比對,計算遺傳距離,采用Poisson模型,對空位采用完全刪除(Complete deletion),并用“bootstrap”1 000次自舉檢測各分支的置信度,構建NJ系統進化樹,采用Evolview 3.0對進化樹進行美化。
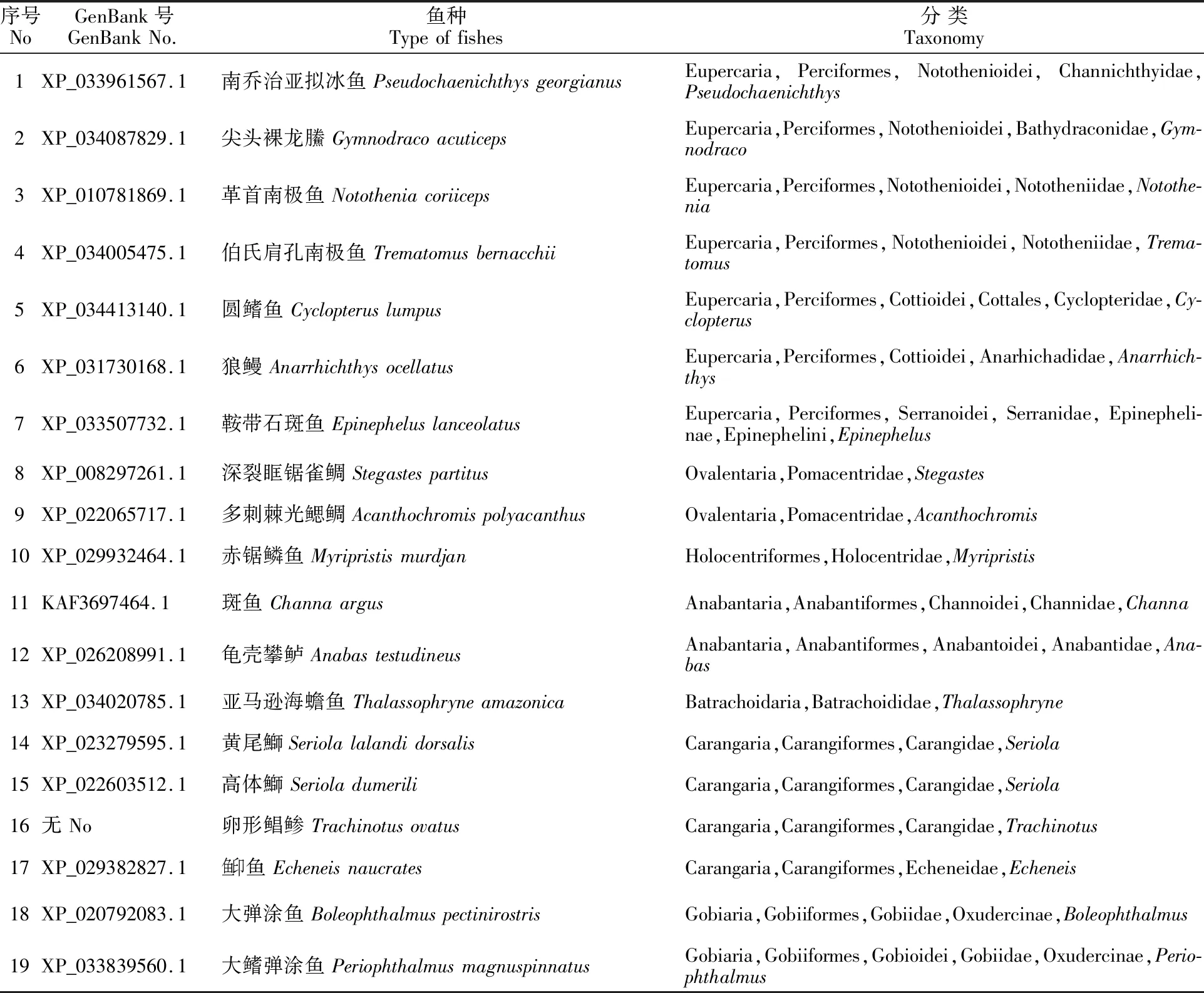
表1 19種魚類的 SST1基因信息Table 1 SST1 gene information of 19 fish species
參考有關文獻中的相關方法[17-18],利用NCBI的ORF finder對卵形鯧鲹的SST1基因序列進行開放閱讀框及其所編碼的氨基酸序列進行預測分析;采用Protparam分析基因所表達SST1蛋白的理化特性;DNASTAR 7.1分析SST1蛋白的親水性、抗原指數,NetSurfP預測蛋白的二級結構,Phyre2同源構建其三維結構模型,并用SAVES 5.0軟件對構建的模型進行評估;TMHMM分析其跨膜結構;采用SignalP分析其信號肽,采用NetPhos 2.0分析其磷酸化位點,PSORT預測其亞細胞定位。在線分析軟件的分析內容與網址信息見表2。

表2 SST1基因在線分析軟件信息Table 2 Information of online analysis software for SST1 gene
1.2.2 總RNA提取及反轉錄 按照RNA提取試劑盒[天根生物科技(北京)有限公司]的操作說明,分別提取卵形鯧鲹14個胚胎發育時期組織樣品和幼魚14個臟器組織樣品的總RNA,采用NanoDrop One檢測RNA質量濃度,通過20 g/L 瓊脂糖凝膠電泳檢測其完整度。再根據Hiscript Ⅲ反轉錄試劑盒操作說明進行反轉錄,合成 cDNA第一鏈。
1.2.3 卵形鯧鲹SST1基因的表達分析 以18S rRNA為內參基因[19],根據卵形鯧鲹SST1基因序列設計熒光定量PCR引物(SST1-F:5′-GGTCCCTGATGTGTCGGTTGG-3′;SST1-R:5′-ACAGTGGGTCTGCGGTAGCG-3′),以卵形鯧鲹14個不同胚胎發育時期和幼魚14個組織的cDNA為模板進行熒光定量PCR擴增。反應體系為15 μL,其中7.5 μL的ChamQ SYBR qPCR Mix,上下游引物(10 μmol/L)各0.3 μL,cDNA模板1.2 μL,5.7 μL的ddH2O,每個樣品3個重復。SST1基因的表達量采用相對Ct法(2-△△Ct)進行統計分析。
1.2.4 統計分析SST1基因在卵形鯧鲹14個不同胚胎發育時期和幼魚14個組織中的表達量結果用 “平均值±標準差”來表示,運用統計學軟件IBM SPSS Statistics 19.0對結果進行相對獨立的單因素方差分析(ANOVA),利用Duncan’s多重比較分析各組織間的差異顯著性,P<0.05為差異顯著,P<0.01為差異極顯著。
2 結果與分析
2.1 SST1基因的生物學信息
2.1.1SST1同源性及進化分析 將卵形鯧鲹SST1基因的氨基酸序列與同科近緣魚類黃尾鰤(XM_023423827)和高體鰤(XM_022747791.1)進行序列比對,序列相似度為92.38%和 91.87%,確定該序列為卵形鯧鲹的SST1基因序列。利用卵形鯧鲹SST1基因與其他18種魚類的SST1基因比對分析,構建ML系統進化樹(圖1)。所分析的魚類主要聚成3支,其中一支為鱸形目(Perciformes)魚類,包括南極魚亞目(Notothenioidei)、杜父魚亞目(Cottioidei)、鮨亞目(Serranoidei)魚類;第二支主要由雀鯛科(Pomacentridae)、金鱗魚科(Holocentridae)、攀鱸目(Anabantiformes)、蟾魚科(Batrachoididae)等魚類聚合而成;第三支主要由鲹形目(Carangiformes)、蝦虎魚目(Gobiiformes)等魚類聚合而成。其中卵形鯧鲹、黃尾鰤與高體鰤均屬于鲹科(Carangidae)魚類,具有較近的遺傳距離,而魚印魚(Echeneis naucrates)為魚印科(Echeneidae)魚類,遺傳距離則稍遠。
2.1.2SST1基因的序列基本結構及編碼蛋白的理化性質分析 卵形鯧鲹SST1基因的cDNA序列長度為1 155 bp(圖2),序列中A、C、G、T含量分別為20.78%(240個)、26.58%(307個)、25.89%(299個)和26.75%(309個),GC含量(52.48%)高于AT含量(47.53%),說明編碼區的DNA雙鏈較為穩定。卵形鯧鲹SST1基因序列的開放閱讀框為1 104 bp,編碼368個氨基酸,其包含7 TM_GPCR_Srx結構域。所編碼的蛋白分子質量為41.42 ku,理論等電點為8.47,為堿性蛋白。帶負電荷的殘基總數(Asp + Glu)為26,帶正電荷的殘基總數(Arg + Lys)為31,說明SST1所編碼的蛋白為帶正電的蛋白。蛋白不穩定性指數(Ⅱ)為39.82,說明該蛋白為較穩定的蛋白。
卵形鯧鲹SST1基因編碼的蛋白平均親水系數(GRAVY)為0.539,說明該蛋白為疏水性蛋白。序列中含有較多的疏水性氨基酸,疏水性區段主要集中于34~63、70~95、103~134、152~175、195~226、246~271、279~289、311~318位氨基酸(圖3)。疏水性氨基酸中色氨酸Trp、苯丙氨酸Phe、纈氨酸Val、亮氨酸Leu、異亮氨酸Ile、丙氨酸Ala和甲硫氨酸Met分別占編碼氨基酸的1.90%、5.71%、11.14%、10.60%、6.79%、8.15%和3.50%。跨膜區分析結果(圖4)顯示,SST1蛋白存在7個跨膜結構域,跨膜結構域的位置為36~58、71~93、108~130、151~173、198~220、247~269、279~301。
2.1.3SST1基因編碼蛋白的抗原性及亞細胞定位分析 采用Protean中的Jameson-Wolf方法預測卵形鯧鲹SST1蛋白的抗原指數,了解其可能的抗原表位位點。由圖5可看出,其抗原指數較高的區段為第22~35、64~68、141~150、176~185、234~245、272~278、301~310、318~328、333~339、341~351、354~362位氨基酸。SST1蛋白全長序列中潛在的抗原表位相對不是很豐富,且相對較集中在序列的尾端。對SST1亞細胞定位進行預測分析,發現其主要定位于細胞質膜,推測其在細胞膜發揮作用。
2.1.4SST1基因編碼蛋白的信號肽及磷酸化位點分析 采用Signal P對SST1蛋白進行預測分析,未發現信號肽位點(圖6)。磷酸化位點預測結果(圖7)顯示,絲氨酸磷酸化位點有4個,主要位于240、308、320、341位點;蘇氨酸磷酸化位點5個,主要位于149、339、362、366、367位點;酪氨酸磷酸化位點8個,主要位于9、24、27、28、58、133、329、361位點。這17個磷酸化位點的可信度均為0.70以上,說明其可信度較高。
2.1.5 SST1蛋白結構預測 采用NetSurfP對SST1蛋白的二級結構進行在線預測,結果如圖8所示,SST1蛋白二級結構主要包含α-螺旋(Alpha helix)、延伸鏈(Extended strand)及無規則卷曲(Random coil)3 種類型,其中α-螺旋氨基酸有150個,占總數的40.76%、延伸鏈氨基酸60個,占總數的16.13%、無規則卷曲氨基酸149個,占總數的40.49%。
利用Phyre 2對SST1蛋白進行三級結構預測,構建SST1三維結構模型,同源建模的模板序列與SST1序列的一致度為43%,模型的可信度為100%。SST1蛋白三維結構(圖9-a)顯示,其主體結構為由7段α-螺旋并列扭曲排列成一柱狀結構,該結構也是跨膜區的主要結構,柱狀結構的一端連接一個由4個α-螺旋排列成的小短柱狀結構,位于胞外(圖9-b)。采用SAVES在線程序對構建的SST1蛋白模型進行評估,結果顯示,77.70%的殘基平均3D-1D得分≥ 0.2,ERRAT質量評分為100,Procheck 9項評估中,通過4項,警告5項,無錯誤項,說明構建的模型可靠性較高。
2.2 SST1基因在卵形鯧鲹不同胚胎發育時期的表達分析
SST1基因在卵形鯧鲹胚胎發育的14個關鍵時期均有表達,結果見圖10。由圖可知,從受精卵期到晶體出現期SST1基因都是低表達;進入初孵仔期SST1基因表達量開始急劇增加,極顯著高于此前的各個胚胎發育時期(P<0.01)。
2.3 SST1基因在卵形鯧鲹幼魚不同組織中的表達分析
對卵形鯧鲹幼魚的14個組織進行SST1表達量檢測,結果見圖11,SST1基因在卵形鯧鲹腦組織中呈現高度的表達,其表達量極顯著高于其他13個組織(P<0.01),其次是在卵巢中也呈現高度的表達,其表達量極顯著高于另外12個組織(P<0.01),在精巢中的表達量低于卵巢,但顯著高于皮膚組織(P<0.05),極顯著高于其他10個組織(P<0.01),皮膚組織的表達量相對較低,但顯著高于其他10個組織(P<0.05)。肌肉、心、肝等10個組織的表達量極低,其組間差異不顯著(P>0.05)。
3 討 論
SST1作為SS其中的一個特異性受體,介導了SS在動物生長發育過程中所發揮的負調控作用[20]。目前,SST1基因在赤點石斑魚(Epinephelusakaara)[21]、斜帶石斑魚(Epinepheluscoioides)[22]、線翎電鰻(Apteronotusalbifrons)[23]、虹鱒(Oncorhynchusmykiss)[24]等魚類中已被鑒定或克隆。本研究結果表明,卵形鯧鲹SST1基因開放閱讀框1104 bp,編碼368個氨基酸,與赤點石斑魚[21]384個氨基酸大小接近,蛋白分子質量為41.42 ku,帶正電荷,理論等電點為8.47,屬于堿性蛋白,且蛋白較穩定。亞細胞定位與跨膜結構分析結果顯示,蛋白主要定位于細胞質膜,存在7個跨膜結構,三級結構顯示,其主體結構是由7段α-螺旋并列扭曲排列成一柱狀結構,位于跨膜區,這與SST1屬于GPCRs超家族,而GPCRs由7個跨膜受體結構域組成結果一致[25]。
磷酸化是蛋白質翻譯后修飾(Protein post-translational modifcation,PTM)中最常見的類型,在真核生物的幾乎所有細胞過程中都發揮重要的作用[26]。蛋白的磷酸化是在蛋白質上引入帶負電的磷酸基團,導致蛋白質構象和功能活性發生變化,大多數磷酸化發生在蛋白質的絲氨酸殘基上,蘇氨酸和酪氨酸殘基的磷酸化作用不如絲氨酸殘基的磷酸化[27]。蛋白的磷酸化調節細胞的多種生物過程,如基因表達、代謝、細胞周期調節、分化和凋亡等[28-29]。據估計,大約30%的人類蛋白質在其生命周期的某個時刻被磷酸化[28]。本研究發現,卵形鯧鲹SST1蛋白具有17個磷酸化位點,其中絲氨酸磷酸化位點有4個,主要位于蛋白的第3細胞內環和C-端,定位于細胞內。蘇氨酸磷酸化位點有5個,主要位于蛋白的第2細胞內環和C-端,定位于細胞內;酪氨酸磷酸化位點有8個,主要位于蛋白的N-端、第1、2細胞內環和C-端,定位于細胞外和細胞內,而對于SST來說,第3細胞內環和C-端潛在磷酸化位點對磷酸化、內化和脫敏非常重要[30-31]。預測結果顯示,卵形鯧鲹SST1蛋白存在較多的磷酸化位點,推測該蛋白存在磷酸化現象,但需通過質譜分析等方法進一步研究。
系統發育樹為研究物種起源及其分子進化,進而探索基因功能提供了依據。系統發育分析表明SST家族可聚為2個大分支和7個主要分支,其中一個大分支包含SST2、3、5和7,另一個包含SST1、4和6,SST1和SST4彼此密切相關,SST6型存在于許多魚類中,但在四足動物和鳥類中不存在[9],而在人類、其他四足動物和鳥類中存在的SST4型似乎在所有被調查的硬骨魚類基因組中都缺失[32-33]。本研究對卵形鯧鲹與其他18種魚類SST1基因的系統發育分析表明,系統發育樹有3個主要分支,其中一支由鱸形目(Perciformes)7種魚類組成,第二支主要由雀鯛科(Pomacentridae)、金鱗魚科(Holocentridae)、攀鱸目(Anabantiformes)與蟾魚科(Batrachoididae)等6種魚類聚合而成;第三支主要由鲹形目(Carangiformes)、蝦虎魚目(Gobiiformes)等6種魚類聚合而成,其中卵形鯧鲹、黃尾鰤與高體鰤均屬于鲹科(Carangidae)魚類,顯示了較近的遺傳距離。SST1基因的系統發育樹代表了這些物種SST1基因的進化關系,但與這些物種的系統分類學存在較高的一致性。
SS與SST廣泛存在于魚類發育的不同時期以及各個組織中。SST的表達直接影響著SS對GH分泌的雙向調節作用[34]。本研究發現SST1在卵形鯧鲹的初孵仔期之前的受精卵期、4細胞期、16細胞期、多細胞期、高囊胚期、原腸早期、原腸中期、原腸末期、胚體形成期、眼囊期、耳囊期、心臟跳動期和晶體出現期這13個胚胎發育時期都是微量的低表達,直到初孵仔期表達量迅速升高。這一結果與生長抑素前體(PreprosomatostatinⅠ,PPSⅠ)的mRNA在大西洋鱈魚(Gadus morhua)胚胎發育過程中的表達量變化趨勢一致[35]。說明SS高表達能抑制GH的分泌,低表達卻能促進GH的分泌促進細胞增殖和分化,而SS與SST基因的表達受胚胎發育過程以及動物的營養狀況有關。
雖然SST的基因分布存在重疊現象,但卻具有明顯的表達模式特異性,SST作用機制存在組織特異性應答[36]。本研究通過熒光定量PCR分析發現,SST1在卵形鯧鲹不同組織中的表達量差異極顯著,其中在腦部組織中顯著高度表達,其次是卵巢和精巢,而在其他10個組織中的表達量則較低。Slagter等[24]對虹鱒(Oncorhynchus mykiss)的研究發現,SST1在腦、胃、胰腺和腎臟中高表達,而Zhang等[21]發現赤點石斑魚SST1在20個組織中廣泛表達,并且在每個組織中的表達水平非常相似。上述研究說明SST1可能介導了卵形鯧鲹的多種生理功能,其表達分布可能存在組織特異性和種屬特異性,不同種屬的SST1基因表達模式的差異,表明其在調節不同的生理過程中可能存在功能差異。
大腦和垂體表達多種神經肽,如GH、促性腺激素釋放激素(GnRH)、卵泡刺激素(FSH)、黃體生成激素(LH)、催乳素(PRL)和睪酮(testosterone,Ts)等[37],它們共同調控動物的生長和繁殖,并且SST也參與了這些過程[4,38]。本研究結果顯示卵形鯧鲹腦部SST1高表達,說明SST1可能參與了神經內分泌活動。卵巢中SST1的高表達表明它可能參與生殖激素的調節,在雌性性腺發育中發揮重要作用。
4 結 論
總之,SST1在卵形鯧鲹發育的不同時期和不同組織中均有表達,其中,在初孵仔期呈現高表達以抑制GH的分泌,在其他時期則呈現低表達,更有利于GH分泌促進細胞的增殖分化與機體發育;在腦組織中表達量最高,其次是卵巢和精巢,表明SST1可能在神經內分泌、雌性性腺發育及精子發生過程中都發揮著重要作用。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
