中部地區(qū)家養(yǎng)山羊線粒體遺傳多樣性與母系起源研究
2021-10-21 02:02:54趙金艷王先寧尹慧茹哈斯通拉嘎王擁慶韓浩園
中國(guó)畜牧雜志 2021年10期
趙金艷,權(quán) 凱,王 俊 ,王先寧,尹慧茹,李 君,哈斯通拉嘎,王擁慶,韓浩園*
(1.河南牧業(yè)經(jīng)濟(jì)學(xué)院動(dòng)物科技學(xué)院,河南鄭州 450046;2.沈丘縣農(nóng)牧科技研發(fā)中心,河南周口 466300)
山羊?qū)儆隈Z化較早的牲畜之一。山羊群體具有多母系起源,遺傳多樣性豐富。家養(yǎng)山羊(Capra hircus)由野生山羊進(jìn)化而來(lái),捻角野山羊(C.aegagrus)和旋角野山羊(C.falconeri)被認(rèn)為是家養(yǎng)山羊的兩大野生祖先。線粒體DNA(Mitochondrial DNA,mtDNA)是動(dòng)物唯一的核外遺傳物質(zhì),遵循嚴(yán)格的母系遺傳[1]。其中,片段長(zhǎng)度為1 220 bp 的D-環(huán)控制區(qū)(D-loop)受自然選擇影響較大,存在巨大變異,所以廣泛應(yīng)用于哺乳動(dòng)物群體遺傳多樣性、物種親緣關(guān)系和種群間系統(tǒng)進(jìn)化等方面的研究[2]。Naderi 等[3]發(fā)現(xiàn)7 個(gè)高度分化的線粒體單倍型分支(A、B1、B2、C、D、F、G)。
我國(guó)擁有豐富的山羊品種資源,長(zhǎng)期的自然選擇和人工選擇可能是導(dǎo)致南北地區(qū)家養(yǎng)山羊遺傳多樣性和種質(zhì)分化的主要原因。中部地區(qū)地方山羊品種豐富,有山西省太行黑山羊、山東省濟(jì)寧青山羊、河南省伏牛白山羊、槐山羊和堯山白山羊等。其中,太行黑山羊是極其珍貴的食品資源,對(duì)飼養(yǎng)環(huán)境有較強(qiáng)的適應(yīng)性,放牧、圈養(yǎng)均能適應(yīng),尤其是肉質(zhì)美味可口,目前由于個(gè)體小、生長(zhǎng)慢、屠宰率低等原因群體數(shù)量急劇減少[4];濟(jì)寧青山羊是優(yōu)良的羔皮用山羊品種,具有性成熟早、多胎多羔、常年發(fā)情、抗病力強(qiáng)等優(yōu)良種質(zhì)特性,近年來(lái),濟(jì)寧青山羊由于受體型瘦小、生長(zhǎng)速度緩慢等缺點(diǎn)的限制,純種處于瀕危狀態(tài)[5-6];伏牛白山羊具有產(chǎn)肉性能好、屠宰率高、繁殖性能好、耐粗飼、抗病力強(qiáng)、適宜山地放牧等優(yōu)點(diǎn);槐山羊具有適應(yīng)性強(qiáng)、繁殖力強(qiáng)、抗病力強(qiáng)、肉質(zhì)鮮美、皮張質(zhì)量好等優(yōu)點(diǎn);堯山白山羊是肉皮兼用型山羊品種,具有個(gè)體大、抗病力強(qiáng)、四肢粗壯、屠宰率高等特性,尤其是肉質(zhì)膻味較輕而備受當(dāng)?shù)叵M(fèi)者青睞,近年來(lái)開(kāi)展肉羊雜交,純種堯山白山羊分布區(qū)域縮小、數(shù)量銳減[7]。由于市場(chǎng)競(jìng)爭(zhēng)、雜化等各種原因,這些品種數(shù)量近年來(lái)均有下降,為了保護(hù)山羊本地品種種質(zhì)資源的多樣性,有必要系統(tǒng)開(kāi)展中部地區(qū)山羊品種群體遺傳結(jié)構(gòu)和遺傳多態(tài)性的研究,為中部地區(qū)山羊群體種質(zhì)的穩(wěn)定、持續(xù)發(fā)展奠定理論研究基礎(chǔ)。
本研究對(duì)中部地區(qū)的伏牛白山羊、槐山羊、濟(jì)寧青山羊、太行黑山羊和堯山白山羊的線粒體D-loop 區(qū)遺傳多樣性、遺傳結(jié)構(gòu)及母系起源進(jìn)行研究,旨在為中國(guó)中部地區(qū)山羊地方品種資源的評(píng)估、保護(hù)、利用及山羊品種起源、進(jìn)化研究等提供科學(xué)依據(jù)。
1 材料與方法
1.1 樣本采集 本研究共采集145 個(gè)本地山羊品種外周血液樣本,包括20 個(gè)伏牛白山羊、74 個(gè)槐山羊、28 個(gè)濟(jì)寧青山羊、13 個(gè)太行黑山羊、10 個(gè)堯山白山羊,采集樣本的羊只年齡段為6 月齡至4 歲,隨機(jī)采集各品種保種場(chǎng)的雌、雄山羊個(gè)體,樣本采集后置于-20℃保存。
1.2 血液總DNA 提取 采用北京艾德萊生物科技有限公司動(dòng)物基因組DNA 提取試劑盒提取山羊外周血基因組DNA,利用Thermo Nanodrop 1000 檢測(cè)OD260/OD280與DNA 濃度,稀釋至20 ng/μL,4℃保存?zhèn)溆谩?/p>
1.3 山羊D-loop 區(qū)序列PCR 擴(kuò)增 山羊線粒體D-loop區(qū)上游引物(5'-AACCACTATTAACCACATCTA-3')和下游引物(5'-CACTTACCATGTAAAAGACCC-3')序列由生工生物工程(上海)股份有限公司合成。PCR 擴(kuò)增體系為25 μL:1 μL DNA 模板、1 μL 上/ 下游引物(10 μmol/L)、9.5 μL ddH2O、12.5 μL TaqPCR Master Mix(北京康為世紀(jì)生物科技有限公司)。PCR 擴(kuò)增程序?yàn)椋?5℃預(yù)變性4 min;94℃變性30 s,58 ℃退火30 s,72℃延伸60 s,36 個(gè)循環(huán);72℃延伸9 min,4℃保存。利用1%瓊脂糖凝膠電泳檢測(cè)PCR 產(chǎn)物,并由生工生物工程(上海)股份有限公司進(jìn)行雙向測(cè)序。
1.4 數(shù)據(jù)分析 利用Lasergene 軟件包SeqMan 軟件拼接雙向測(cè)序的序列,利用MEGA 6.0 軟件[8]進(jìn)行多重比對(duì)分析。下載9 條參考序列:旋角野山羊(AB044305)、捻角野山羊(AB004076)、Lineage A(AY155721)、Lineage B(DQ121578)、Lineage C(DQ188892)、Lineage D(AY155952)、Lineage D(DQ188893)、Lineage F(DQ241349)、Lineage G(EF618084),利用MEGA6.0 軟件構(gòu)建NJ 系統(tǒng)發(fā)育樹(shù),自舉檢驗(yàn)重復(fù)數(shù)為1 000,并基于Kimura 2-parameter(K2P)模型[9]計(jì)算濟(jì)寧青山羊、伏牛白山羊、太行黑山羊、槐山羊和堯山白山羊間遺傳距離。
利用DnaSP V5 軟件進(jìn)行5 個(gè)山羊品種線粒體D-loop 區(qū)遺傳多態(tài)性分析、單倍型分析、品種間遺傳分化、基因流分析和中性檢驗(yàn)[10],利用NETWORK 10.2.0.0 軟件(Fluxus Technology Ltd.,Kiel,Germany)構(gòu)建濟(jì)寧青山羊、伏牛白山羊、太行黑山羊、槐山羊和堯山白山羊單倍型網(wǎng)絡(luò)圖。
2 結(jié)果與分析
2.1 遺傳多態(tài)性分析 本研究對(duì)槐山羊、太行黑山羊、濟(jì)寧青山羊、伏牛白山羊和堯山白山羊線粒體D-loop遺傳多樣性進(jìn)行分析,總位點(diǎn)數(shù)為1 138 bp,缺失位點(diǎn)為156 個(gè),用于計(jì)算遺傳多樣度的位點(diǎn)為982 個(gè)(不包括缺失位點(diǎn)),其中,多態(tài)位點(diǎn)140 個(gè)(排除缺失位點(diǎn)),單態(tài)位點(diǎn)842 個(gè)。突變位點(diǎn)共定義63 個(gè)單倍型,其中,9 個(gè)(Hap 3、5、8、13、16、20、26、30、33)為 共享單倍型,品種特異單倍型為54 個(gè)。由表1 可知,5 個(gè)山羊品種總單倍型多樣度(Hd)為0.965±0.004,核苷酸多樣度(Pi)為0.013 88±0.000 66,研究發(fā)現(xiàn)伏牛白山羊的單倍型多樣度最低(0.831),槐山羊的單倍型多樣度最高(0.957),太行黑山羊的核苷酸多樣度最低(0.011 93),濟(jì)寧青山羊的核苷酸多樣度最高(0.014 34)。
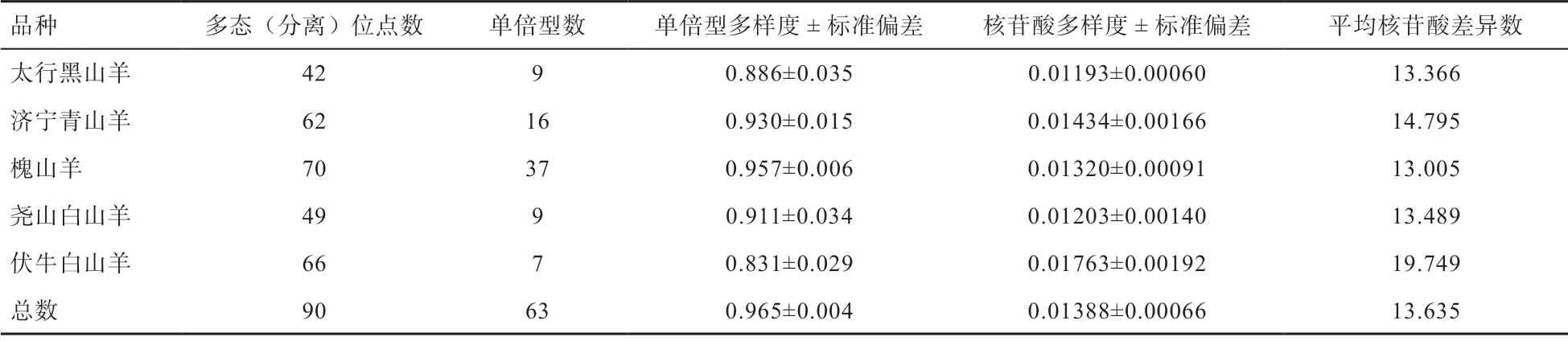
表1 5 個(gè)山羊品種線粒體D-loop 遺傳多態(tài)性分析
2.2 群體結(jié)構(gòu)分析 本研究將捻角野山羊、旋角野山羊和6 個(gè)D-loop 分支(Lineage A、B、C、D、F、G)作為參考序列,構(gòu)建濟(jì)寧青山羊、太行黑山羊、槐山羊、伏牛白山羊和堯山白山羊系統(tǒng)進(jìn)化樹(shù)(圖1、2),結(jié)果表明5 個(gè)山羊品種與捻角野山羊聚類,說(shuō)明本研究的5 個(gè)山羊品種母系起源為捻角野山羊;大部分山羊個(gè)體(130 個(gè))屬于支系A(chǔ)(Lineage A),極少數(shù)個(gè)體(15個(gè))屬于支系B(Lineage B);支系A(chǔ) 包括濟(jì)寧青山羊、太行黑山羊、槐山羊、伏牛白山羊和堯山白山羊,支系B 包括濟(jì)寧青山羊、槐山羊、伏牛白山羊和堯山白山羊。如表2 所示,5 個(gè)品種間遺傳距離為0.013 6~0.018 8,均未達(dá)到亞種間遺傳距離。伏牛白山羊與其他4 個(gè)品種的親緣關(guān)系相對(duì)較遠(yuǎn),與濟(jì)寧青山羊親緣關(guān)系最遠(yuǎn),堯山白山羊與其他4 個(gè)品種的親緣關(guān)系相對(duì)較近。
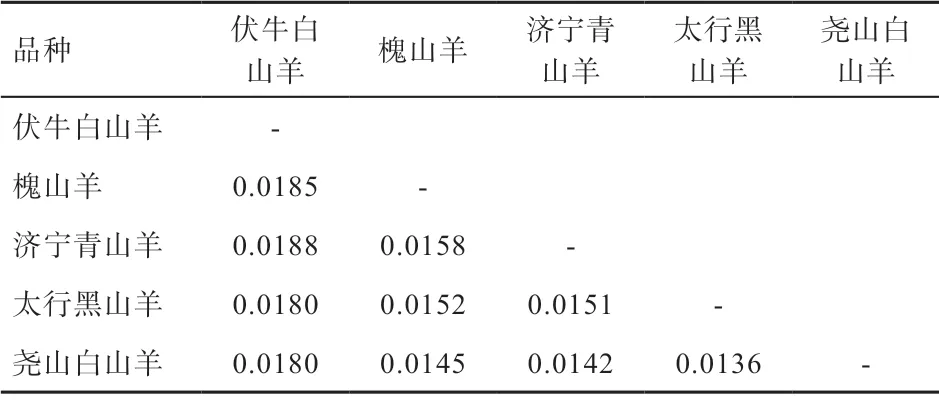
表2 5 個(gè)山羊品種間遺傳距離
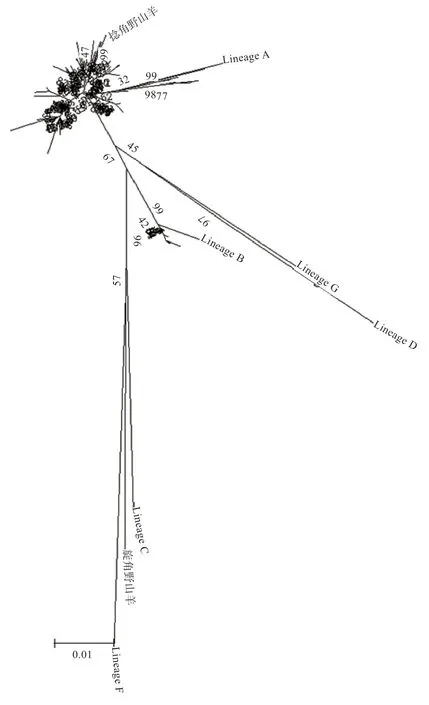
圖1 5 個(gè)山羊品種系統(tǒng)進(jìn)化樹(shù)原始樹(shù)
2.3 群體遺傳分化分析 遺傳分化結(jié)果如表3 所示,品種間核苷酸平均差異數(shù)(Kxy)為11.406 8~16.242 9,遺傳分化系數(shù)(Fst)為0.016 6~0.169 3,核苷酸歧異度(Dxy)為0.011 6~0.016 5。伏牛白山羊與濟(jì)寧青山羊間的Kxy和Dxy最大,說(shuō)明2 個(gè)品種間核苷酸差異最大,遺傳關(guān)系最遠(yuǎn),與遺傳距離結(jié)果一致。伏牛白山羊與堯山白山羊和太行黑山羊間Fst達(dá)到明顯分化程度,其他品種間均為中度分化或分化程度很弱,槐山羊與濟(jì)寧青山羊和堯山白山羊、濟(jì)寧青山羊與堯山白山羊之間均分化程度很弱,說(shuō)明品種間存在基因交流,與基因流(Nm)為3.92 的結(jié)果一致。

表3 品種間遺傳分化系數(shù)
2.4 群體擴(kuò)張分析 由表4 可知,中性檢驗(yàn)結(jié)果表明伏牛白山羊、濟(jì)寧青山羊和太行黑山羊Fu and Li's F 檢驗(yàn)結(jié)果為正值,且顯著偏離中性,表明伏牛白山羊、濟(jì)寧青山羊和太行黑山羊近期經(jīng)歷過(guò)瓶頸效應(yīng)和平衡選擇。槐山羊和堯山白山羊Fu and Li's F 檢驗(yàn)均為P>0.10,不顯著,符合動(dòng)態(tài)平衡,未出現(xiàn)群體大規(guī)模擴(kuò)張或瓶頸現(xiàn)象,群體大小保持相對(duì)穩(wěn)定。
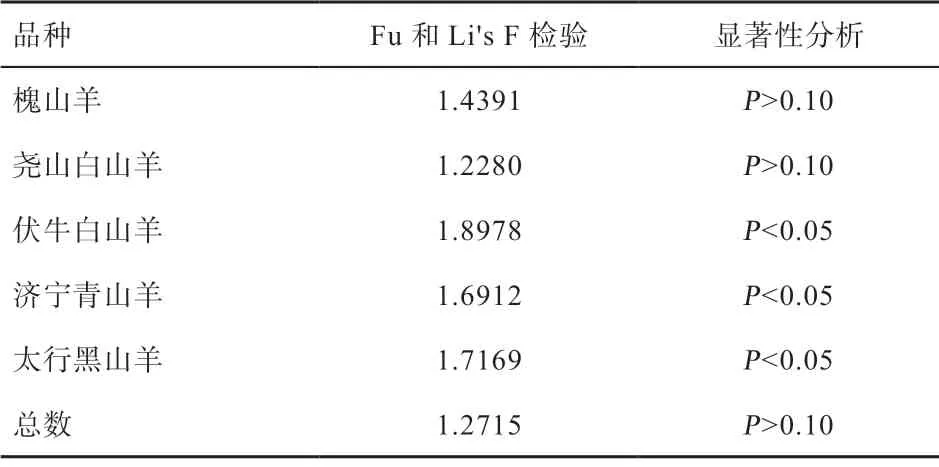
表4 5 個(gè)山羊品種群體擴(kuò)張中性檢驗(yàn)結(jié)果
2.5 群體單倍型及母系起源分析 如表1 所示,本研究共發(fā)現(xiàn)63 個(gè)單倍型,共有單倍型為9 個(gè),品種特異單倍型為54 個(gè),槐山羊單倍型最多,為37 個(gè),濟(jì)寧青山羊定義16 個(gè)單倍型,太行黑山羊與堯山白山羊均為9 個(gè)單倍型,伏牛白山羊單倍型最少為7 個(gè)。由圖3 可知,63 個(gè)單倍型可分為2 個(gè)單倍群(Haplogroup A 和Haplogroup B),分別對(duì)應(yīng)系統(tǒng)發(fā)育樹(shù)中的支系A(chǔ) 和支系B,說(shuō)明濟(jì)寧青山羊、槐山羊和伏牛白山羊存在2 個(gè)母系起源(支系A(chǔ) 和支系B),堯山白山羊和太行黑山羊?yàn)閱蝹€(gè)母系起源(支系A(chǔ))。且每個(gè)支系呈現(xiàn)“星狀”,分布清晰,沒(méi)有交叉。
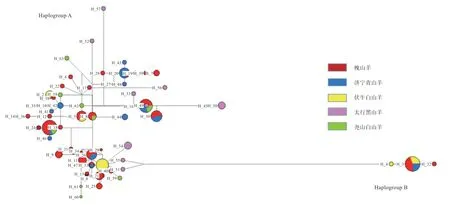
圖3 單倍型網(wǎng)絡(luò)圖
3 討 論
3.1 中部地區(qū)山羊遺傳多樣性Hd和Pi是評(píng)價(jià)群體遺傳多樣性和分化的重要指標(biāo)[11]。本研究發(fā)現(xiàn)5 個(gè)山羊品 種 總Hd為0.965±0.004,Pi為0.013 88±0.000 66。其中,太行黑山羊Hd為0.886,Pi為0.01193。閆益波等[4]研究發(fā)現(xiàn)太行黑山羊Hd值為0.998;武建亮等[12]研究結(jié)果為0.800 3。本研究結(jié)果位于這2 個(gè)數(shù)值之間,說(shuō)明山西太行黑山羊群體多態(tài)性較高,近年來(lái)群體遺傳多樣度較為豐富,并維持穩(wěn)定,具有一定的選擇潛力。本研究中伏牛白山羊Hd為0.831,Pi為0.017 63,與李婉濤等[13]人發(fā)現(xiàn)的伏牛白山羊群體線粒體D-loop 區(qū)存在著豐富的多態(tài)性結(jié)果一致。濟(jì)寧青山羊在5 個(gè)品種中遺傳多樣度較高,Hd為0.930,Pi為0.013 20,遺傳多樣性信息豐富,提供的遺傳信息量大,這可能與品種選育過(guò)程中育種群體導(dǎo)入了外血、群體選育時(shí)間短等因素有關(guān)[14]。
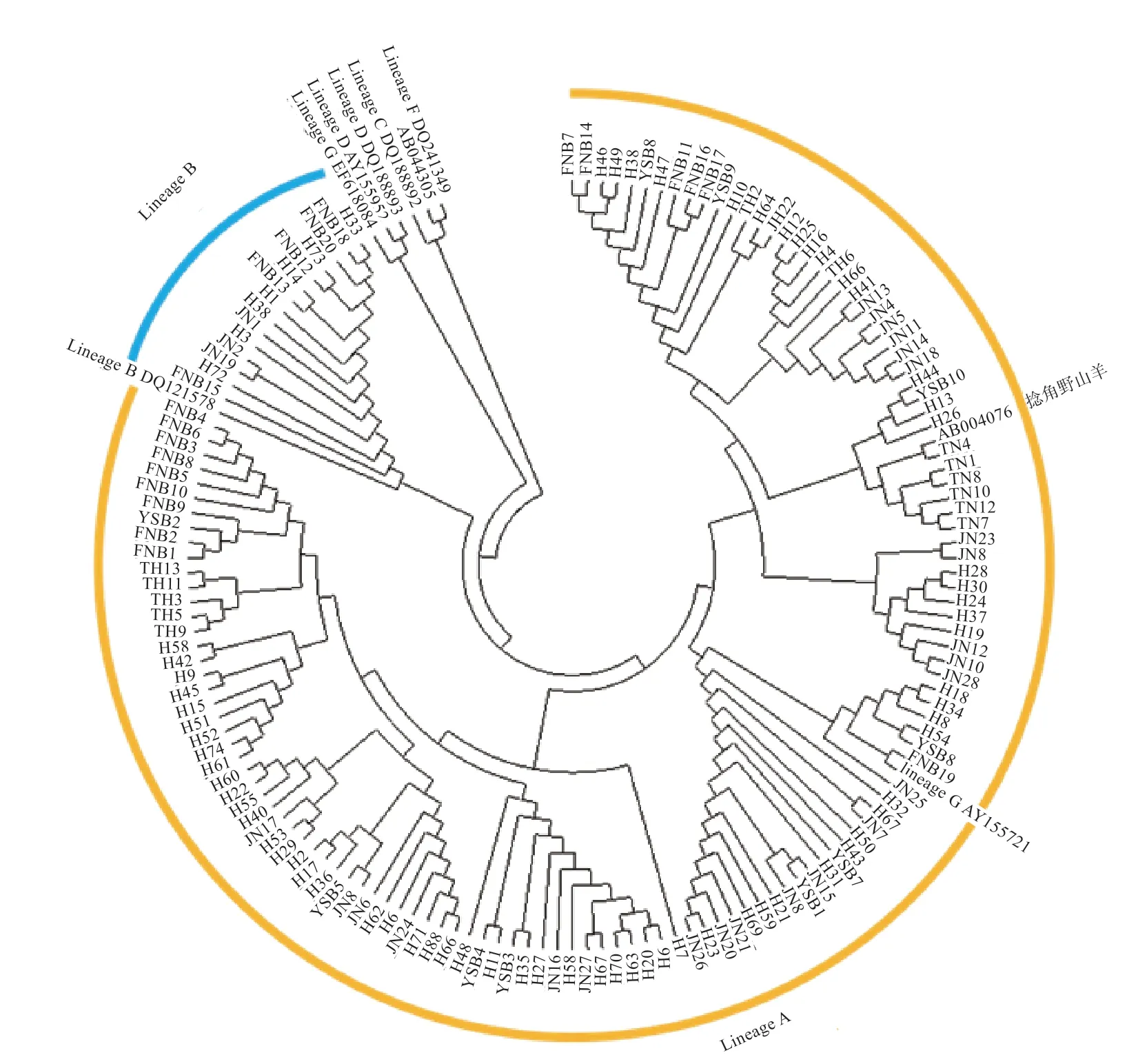
圖2 5 個(gè)山羊品種系統(tǒng)進(jìn)化樹(shù)自展一致樹(shù)
郝榮超等[15]曾對(duì)中國(guó)南部地區(qū)家養(yǎng)山羊遺傳多樣性進(jìn)行研究,發(fā)現(xiàn)南部地區(qū)家養(yǎng)山羊平均Hd和平均Pi分別為0.954 和0.019 57,西北地區(qū)山羊分別為0.919和0.019 08,與本研究中原地區(qū)山羊品種遺傳多樣性相近。有研究針對(duì)河南5 個(gè)地方山羊品種的遺傳多樣性進(jìn)行研究,發(fā)現(xiàn)河南省地方山羊的變異主要存在于品種內(nèi),品種間的變異相對(duì)較小,有足夠的遺傳變異適應(yīng)環(huán)境的變化和人們的選擇需要[16]。
3.2 群體結(jié)構(gòu)分析 本研究遺傳距離結(jié)果表明5 個(gè)品種間遺傳距離為0.013 6~0.018 8,郝榮超等[15]發(fā)現(xiàn)西北地區(qū)山羊品種間遺傳距離為0.016~0.019,南部地區(qū)品種間遺傳距離為0.017~0.025,說(shuō)明中原地區(qū)山羊品種間遺傳距離較近,品種間交流較南方和西北地區(qū)更頻繁。本研究結(jié)果顯示太行黑山羊與其他4 個(gè)品種間的遺傳距離范圍為0.014 2~0.018 0,均未達(dá)到亞種間遺傳距離。閆益波等[4]研究表明山西太行黑山羊與槐山羊的遺傳距離為0.014,山西太行黑山羊和槐山羊分布的東北和華東地區(qū)冬季寒冷,對(duì)毛皮性狀具有較大的選種要求,這2 個(gè)品種都以產(chǎn)絨性狀和板皮性狀為優(yōu),因而群體間起源較近。
Fst可以反映群體近交或群體間遺傳分化程度。本研究中除伏牛白山羊與太行黑山羊、堯山白山羊之間達(dá)到明顯分化,其他品種間Fst為0.016 6~0.122 0,說(shuō)明群體間的遺傳分化水平低,而且基因流大于1,均質(zhì)化作用強(qiáng),能夠抵制遺傳漂變作用,防止群體間遺傳分化發(fā)生。王可等[6]也發(fā)現(xiàn)濟(jì)寧青山羊的遺傳多樣性得到了保留,群體間的基因流較大,遺傳分化水平較低。閆益波等[4]發(fā)現(xiàn)山西太行黑山羊與槐山羊的群體間分化程度較低,均與本研究結(jié)果一致。有研究對(duì)中國(guó)其他地區(qū)山羊品種遺傳分化進(jìn)行研究,F(xiàn)st與本研究結(jié)果范圍一致[16-17]。
3.3 母系起源分析 李祥龍等[18]發(fā)現(xiàn),家養(yǎng)山羊中既有彎角也有旋角,因此認(rèn)為捻角野山羊和旋角野山羊都是家養(yǎng)山羊的野生祖先。實(shí)際上由于捻角野山羊的地理分布與山羊最初家養(yǎng)的地方有高度的相關(guān)性,所以捻角野山羊是最可能的候選者[19]。本研究發(fā)現(xiàn)槐山羊、濟(jì)寧青山羊、伏牛白山羊、太行黑山羊和堯山白山羊均與捻角野山羊聚為一類,旋角野山羊單獨(dú)聚類。本研究結(jié)果與其他中國(guó)家養(yǎng)山羊母系起源進(jìn)化研究結(jié)果一致[15,20],均認(rèn)為捻角野山羊是中國(guó)家養(yǎng)山羊的野生祖先。
中國(guó)山羊品種存在A、B、C 和D 4 個(gè)分支[21],A和B 是在我國(guó)山羊品種中廣泛存在的類型,且經(jīng)歷過(guò)群體擴(kuò)張,而類型C 和D 則以低頻率存在于我國(guó)部分山羊個(gè)體中[15,22],本研究印證了這些觀點(diǎn)。本研究聚類發(fā)現(xiàn),中原地區(qū)山羊共分為兩大支系,支系A(chǔ) 和支系B,且槐山羊、濟(jì)寧青山羊、伏牛白山羊和堯山白山羊在A、B 支系均有分布。而太行黑山羊僅來(lái)源于支系A(chǔ)。當(dāng)然,要澄清山羊的起源進(jìn)化問(wèn)題,可能僅有mtDNA D-loop標(biāo)記是不夠的,必須借助其他遺傳標(biāo)記研究。
4 結(jié) 論
中原地區(qū)5 個(gè)山羊品種線粒體D-loop 區(qū)單倍型多樣度與核苷酸多樣度較高,品種間均未達(dá)到亞種間遺傳距離,伏牛白山羊、濟(jì)寧青山羊和太行黑山羊近期經(jīng)歷瓶頸效應(yīng),槐山羊和堯山白山羊符合動(dòng)態(tài)平衡。系統(tǒng)進(jìn)化樹(shù)結(jié)果表明,中原地區(qū)山羊祖先為捻角野山羊,有支系A(chǔ) 和支系B 2 個(gè)母系起源。
