巴西橡膠樹轉錄中介體HbMed25基因克隆與表達分析
2021-10-16 14:29:00張世鑫吳紹華楊署光晁金泉李招娣田維敏
廣西植物 2021年9期
張世鑫 吳紹華 楊署光 晁金泉 李招娣 田維敏


摘 要:轉錄中介體(mediator,Med)中的Med25基因在植物的組織發育、器官發生、脅迫響應以及調控茉莉酸(JA)信號途徑等方面起重要調控作用。該文在巴西橡膠樹基因組序列中找到了橡膠樹Med25基因的序列,通過RT-qPCR和RACE技術克隆HbMed25基因的全長序列并進行生物信息學預測分析結果顯示,HbMed25基因包含2 655 bp的開放閱讀框,編碼884個氨基酸,蛋白分子量為95 kD,理論等電點為8.68,疏水性蛋白,亞細胞定位最可能為細胞核。Real-time PCR分析結果顯示,HbMed25基因在樹皮中表達量最高,在植物激素JA處理的形成層和膠乳材料中HbMed25基因在處理早期(1 h和2~4 h)上調表達,受割膠處理HbMed25基因在處理早期上調表達且對高產的橡膠樹品系(CATAS 8-79和CATAS 7-33-97)的割膠處理響應更明顯。綜上結果認為,HbMed25基因極大可能參與了JA信號途徑的調控,對闡明JA誘導橡膠樹次生乳管分化的分子調控機制具有促進作用。
關鍵詞:巴西橡膠樹,HbMed25,表達分析,茉莉酸信號途徑,乳管分化,橡膠合成
中圖分類號:Q943
文獻標識碼:A
文章編號:1000-3142(2021)09-1548-13
Abstract:The Med25 gene in the transcription mediator (Med)plays an important regulatory role in many processes,such as tissue development,organogenesis,stress response,and regulation of jasmonate acid(JA)signaling pathway in plants. The Med25 gene sequence was found in the genome of Hevea brasiliensis,and the full-length sequence of HbMed25 gene was cloned by RT-qPCR and RACE technology,and the biological information was predicted and analyzed. The results were as follows:The HbMed25 gene included an open reading frame for 2 655 bp,encoded 884 amino acids,the molecular weight was 95 kD and theoretical isoelectric point was 8.68,hydrophobic protein,and most likely localized in the nucleus in subcellular localization analysis. Real-time PCR analysis results showed that HbMed25 was the most highly expressed in the bark; HbMed25 was up-regulated at the early stage of treatment (1 h and 2-4 h)in the cambium and latex treated by the plant hormone of JA. After tapping,HbMed25 had the highest expression at the early stage of tapping treatment,and up-regulates expression in high-yield rubber clones(CATAS 8-79 and CATAS 7-33-97). All the results indicate that HbMed25 is highly likely to participate in the regulation of the JA signal pathway,and it plays a role in promoting the molecular regulation mechanism of JA-induced secondary laticifer differentiation.
Key words:Hevea brasiliensis,HbMed25,expression analysis,jasmonate signaling pathway,laticifer differentiation,rubber synthesis
中介體復合物(mediator,Med)在植物不同組織發育、花期、花的結構、植物抗逆性等方面具有重要作用。例如:Med8、Med17、Med18、Med20A可以調控植物的花期(Kim et al.,2011);Med18、Med21參與植物響應生物逆境脅迫(Dhawan et al.,2011);Med16參與植物對非生物的逆境響應(Wathugala et al.,2011)。在眾多的Med成員中,Med25的研究相對較多且對基因功能的了解更詳細。AtMed25可調控擬南芥中光敏色素信號的PFTI (Wollenberg et al.,2007; Backstrom et al.,2011),在植物響應逆境脅迫過程中具有顯著作用(Elfving et al.,2011)。AtMed25的缺失突變體植株細胞體積變大、細胞數目增加,導致植物的器官體積增大;過量表達AtMed25的植株細胞體積變小、細胞數目變少,導致植物器官體積減小。可見,AtMed25可以調控植物細胞的增殖速度和體積大小,是植物器官體積大小的負調控因子(Xu & Li,2011)。Chen et al.(2012)發現Med25與抗逆相關的茉莉酸(jasmonate acid,JA)信號途徑有密切聯系,AtMed25通過與AtMYC2直接相互作用使RNA聚合酶Ⅱ招募到AtMYC2 的靶標啟動子區域來調控茉莉酸響應基因表達,之后又發現Med25參與調控茉莉酸信號途徑的關鍵環節COI1-JAZ-MYC2。在植物處于自然狀態下,Med25與JAZ蛋白是競爭關系,競爭結合MYC2的結合位點,Med25與MYC2間的相互作用相對較弱;Med25還與JA受體COI1直接發生相互作用,將COI1招募到茉莉酸途徑的核心轉錄因子MYC2的靶標基因啟動子區域。植物發育或應對逆境脅迫時,產生活性茉莉酸(JA-Ile),Med25促進依賴于活性形式茉莉酸的“COI1-JAZ”受體復合物的形成,引起JAZ蛋白的降解,使Med25與MYC2的相互作用增強,MYC2由轉錄抑制轉變為轉錄激活,招募RNA聚合酶Ⅱ等轉錄機器,激活茉莉酸下游的響應基因的表達(Chen et al.,2012; Zhai & Li,2019; Wu et al.,2019)。
天然橡膠(natural rubber)是四種重要的工業原料和戰略物資之一,世界上99%的天然橡膠來自巴西橡膠樹(Hevea brasiliensis)(祁棟靈等,2012)。巴西橡膠樹的乳管是天然橡膠合成和貯存的地方,與天然橡膠產量直接相關的是樹干樹皮中的次生乳管(郝秉中和吳繼林,1982;Hao & Wu,2000)。乳管由維管形成層細胞分化而來,是一種特化的分泌組織。維管形成層細胞分化成次生乳管的過程,稱為次生乳管分化。我們前期研究發現割膠、機械傷害和茉莉酸類物質都可以誘導橡膠樹的次生乳管分化(Hao & Wu,2000;劉惠芳等,2001;Wu et al.,2002;Zhang et al.,2015;Tian et al.,2015),并建立了一套穩定的體外誘導橡膠樹萌條次生乳管分化的實驗系統。我們前期還發現橡膠樹的乳管細胞中存在特定的茉莉酸信號通路HbCOI1-HbJAZ3-HbMYC2,其通過上調法尼基焦磷酸合酶(FPS)基因和小橡膠粒子蛋白(SRPP)基因的表達增強天然橡膠生物合成(Deng et al.,2018)。因此,我們提出調節橡膠樹次生乳管分化和橡膠生物合成的茉莉酸信號途徑可能是乳管細胞的一條主要信號途徑,并且推測植物中介體Med25在JA信號途徑中起重要的調控作用。
模式植物中Med25的研究較多,其功能被越來越多地揭示出來。然而,橡膠樹中Med25的研究相對滯后。本文克隆了巴西橡膠樹的Med25基因,通過Real-time PCR技術分析其在組織特異性、茉莉酸處理的形成層和膠乳樣品以及不同植物激素處理的膠乳樣品中的基因表達,為揭示Med25參與茉莉酸信號途徑調控橡膠樹次生乳管分化和橡膠生物合成的分子調控機理提供理論依據。
1 材料與方法
1.1 HbMed25基因的克隆
HbMed25基因的開放閱讀框(ORF)信息來自巴西橡膠樹無性系CATAS 7-33-97基因組測序結果(Tang et al.,2016),得到的基因序列信息經過Blast比對后,設計ORF引物擴增出基因開放閱讀框并測序確認。
引物序列HbMed25-OF為5′-ATGGCGGAGAAGCAGCTGGT-3′,HbMed25-OR為5′-TTAACTCATAAATCCACCTCCAGGC-3′。使用橡膠樹無性系CATAS 7-33-97的膠乳cDNA作為模板,進行PCR擴增HbMed25基因的全長cDNA。PCR程序:95 ℃預變性3 min;95 ℃變性30 s,56 ℃退火30 s,72 ℃延伸2 min,循環35 次;72 ℃延伸20 min;16 ℃終止反應。PCR產物通過1%的瓊脂糖凝膠電泳檢測,回收并純化目的條帶,連接轉化到pEASY-Blunt Simple載體上,挑取陽性單克隆,使用M13引物進行菌落PCR篩選,選取插入DNA片段正確的陽性單克隆,送到廣州天一測序公司進行測序,以確定克隆基因cDNA序列的準確性。
1.2 同源性分析和系統進化分析
通過NCBI ORF finder工具對獲得的HbMed25基因序列進行ORF預測和確定,使用DNAMAN 7.0軟件將ORF序列翻譯為相應的氨基酸序列,并進行序列同源性比對;使用NCBI Conserved Domains工具和SMART工具獲取HbMed25蛋白的保守結構域信息;使用ExPASy網站的ProtParam、ProtScale 和TMpred Server等工具進行HbMed25蛋白的理化性質、親疏水性以及跨膜結構的推測;使用The predicted NLS進行氨基酸序列的核定位信號的預測;使用SWISS-MODEL工具對HbMed25蛋白的三級結構進行預測。
通過JGI網站Phytozome工具檢索到來源于陸生植物和藻類的65個Med25的蛋白序列。本文將橡膠樹HbMed25蛋白序列和其他陸生植物和藻類的65個Med25的蛋白序列進行系統進化分析,使用MEGA 6軟件繪制Neighbor joining系統進化樹,并進行1 000 bootstrap的統計學檢測。
1.3 組織特異性表達
所用材料為巴西橡膠樹無性系CATAS 7-33-97的開割成齡樹和一年生萌條,均種植在中國熱帶農業科學院試驗場三隊基地。同一時間采集同一棵橡膠樹的側根、幼莖、穩定期葉片、雌花、種仁(外胚乳)、樹皮、萌條的形成層區和膠乳等樣品進行組織特異性表達,每個樣品均來自3棵樹樣品的混合。采集膠乳樣品時,棄去前1 min流出的膠乳,取割膠后2~5 min流出的膠乳,膠乳直接在1.5 mL RNase free離心管中與裂解液SL(TIANGEN RNAprep Pure PlantKit,DP441)以膠乳與裂解液的體積比為1∶5進行混合,置于冰上,立即提取總RNA 或-80 ℃保存備用;其他組織樣品采集后,液氮速凍,液氮研磨后提取總RNA 或-80 ℃保存備用。
1.4 割膠處理
一天時間中橡膠樹的韌皮部組織膨壓在凌晨時段最高,割膠后的排膠動力最大,隨后在一天中逐步降低,到了晚上逐步恢復(Buttery & Boatman,1966)。凌晨時環境溫度低,能夠降低黃色體破裂導致的膠乳凝集,增加排膠的時間(Shi et al.,2019)。此外,橡膠生物合成關鍵酶基因表達還具有明顯的晝夜節律性(楊鵬等,2019)。為避免割膠時間、環境溫度和節律性等因素對本實驗的干擾,割膠組和對照組需在同一時間進行割膠和采集膠乳。選取同一林段且割齡一致的巴西橡膠樹無性系CATAS 7-33-97的開割樹,分為割膠和對照兩組,割膠組的橡膠樹35株,在同一時間進行割膠處理后,分別在割膠后的0.5、2、6、12、24、48、72 h等時間點進行第二次割膠,采集第二次割膠的膠乳作為不同時間割膠處理的樣品。對照組的35株橡膠樹不進行同一時間割膠處理,僅在割膠組第二次割膠時進行割膠,采集的膠乳樣品作為不同時間割膠處理的對照。每個時間點的開割樹膠乳樣品來自5棵樹膠乳樣品的混合,并采集3個生物學重復。
1.5 植物激素等化學因子處理
用單面刀片刮去面積為2 cm × 4 cm的萌條莖表皮及部分皮層,用滅菌的脫脂棉包裹處理部位,分別施加含有4 500 μg·mL-1 6-BA(對照組為滅菌水)、50 000 μg·mL-1 GA(對照組為滅菌水)、5 000 μg·mL-1 ET(對照組為滅菌水)、0.032 μg·mL-1曲古抑菌素A(trichostatin A,TSA)(對照組為5% DMSO溶液)、6.39 μg·mL-1冠菌素(coronatine,COR)(對照組為滅菌水)的溶液,利用塑料封口膜包裹,分別處理1、2、8、24、48、72、120 h,拆去脫脂棉和塑料封口膜,收集處理部位的膠乳。每個時間點的膠乳樣品來自5根萌條的膠乳混合,并采集3個生物學重復。
按照同樣的方法,使用6.39 μg·mL-1 COR(對照組為滅菌水)的溶液處理,分別處理0.5、1、2、8、24、48、72 h,切取處理部位的萌條樹皮,通過冰凍切片和激光顯微切割的方法,分離形成層區樣品。每個時間點的形成層樣品來自15根萌條樣品的混合,并采集3個生物學重復。
1.6 Real-time PCR 分析
使用總量為1 μg的總RNA樣品,參照Thermo Scientific RevertAid First Strand cDNA Synthesis Kit (K1622)的說明書,反轉錄合成cDNA的第一鏈。反轉錄得到的cDNA模板,稀釋10倍以后,進行Real-time PCR分析。通過Bio-Rad CFX Manager 3.0的分析系統(Bio-Rad Labratories Inc,Ca,USA)進行Real-time PCR分析,以巴西橡膠樹的肌動蛋白基因HbACTIN(Gene bank:JF270598)(Zhao et al.,2011)為內參基因,分析HbMed25基因在橡膠樹各個組織和各種處理樣品中的相對表達量。
Real-time PCR的引物序列HbMed25-QF為5′-GCAGCACCATGATTCCAACTC-3′,HbMed25-QR為5′- AGCCGAAGCACTCCTGTAAC-3′。參照TransStart Tip Green qPCR SuperMix (AQ141-04)的使用說明配制Real-time PCR的反應體系,使用Bio-rad CFX Manager 3.0配套的384孔板進行Real-time PCR的反應,每個反應配制10 μL的反應體系,成分如下:1 μL cDNA模板,0.6 μL的正向引物和反向引物,5 μL TransStart Tip Green qPCR SuperMix,用ddH2O補齊至10 μL。基因的相對表達量,使用Bio-Rad CFX Manager 3.0的分析系統自帶的 2-ΔΔCT 方法進行統計。每個樣品進行3次實驗技術重復。
2 結果與分析
2.1 HbMed25基因的克隆和生物信息學分析
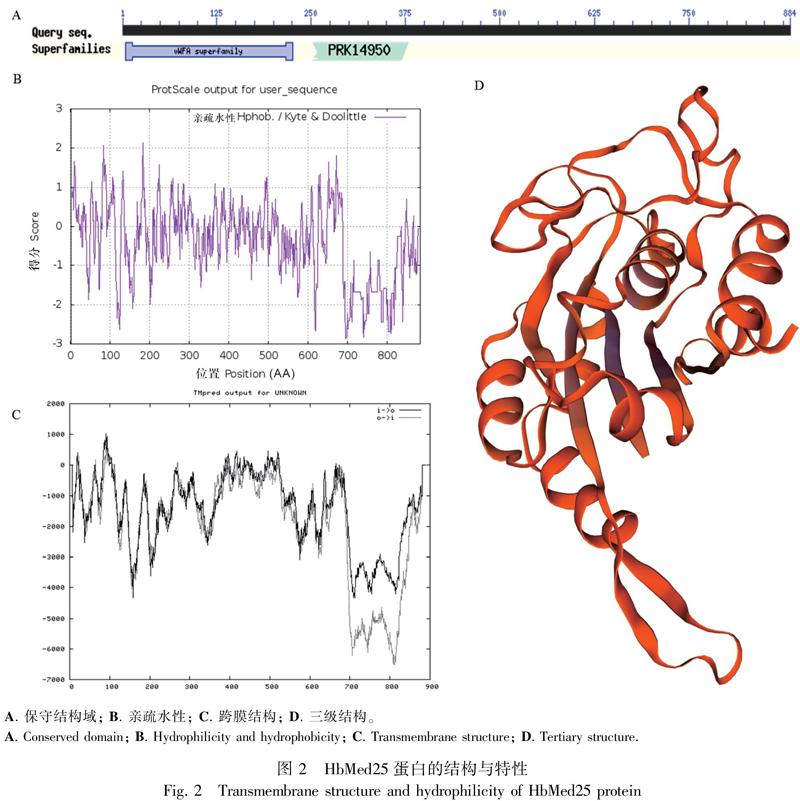
通過基因克隆及測序驗證,從巴西橡膠樹膠乳中擴增獲得HbMed25基因的全長ORF-cDNA序列2 655 bp(圖1)。使用ExPASy ProtParam在線分析工具分析HbMed25推導氨基酸序列的蛋白質理化性質,發現HbMed25氨基酸數量為884(圖1),并存在Med25蛋白特有的MED25_VWA保守結構域(圖2:A),相對分子量為95 kD,理論等電點8.68,HbMed25蛋白為偏堿性蛋白。HbMed25蛋白的脂肪系數為76.45,HbMed25蛋白的不穩定系數為34.83,為穩定蛋白。使用ExPASy-TMpred Server在線分析工具進行HbMed25蛋白的跨膜結構分析,發現HbMed25蛋白為親水性蛋白(圖2:B),HbMed25蛋白的總平均疏水指數均為-0.401,HbMed25蛋白沒有跨膜結構(圖2:C)。使用SignalP4.1 Server的神經網絡算法預測蛋白質的信號肽,結果表明HbMed25蛋白可能不存在信號肽。使用WoLF PSORT在線分析工具預測HbMed25蛋白的亞細胞定位,結果表明HbMed25蛋白能夠定位到細胞核且得分較高。通過The predicted NLS工具進行核定位信號預測,發現HbMed25含有較強的核定位信號,預測的核定位信號得分高達0.86分(分數范圍為0~1),核定位信號序列為EKKLCA,如圖3的紅點所示。利用SWISS-MODEL全自動蛋白質結構同源性建模的方法,建立HbMed25的蛋白三級結構,如圖2:D所示。
2.2 HbMed25基因的系統發育和同源性分析
將HbMed25推導的氨基酸序列,與其他模式植物的Med25基因所編碼的氨基酸序列進行比對,結果顯示HbMed25蛋白序列與擬南芥AtMed25(AT1G25540.2)、木薯MeMed25 (LOC110600417)、蓖麻RcMed25(LOC8260402)和楊樹PtMed25 (PF11265)蛋白,如圖4的五角星所示的序列相似性分別為61.34%、87.01%、51.91%和73.63%,且均具有Med25_VWA結構域(圖3)。HbMed25蛋白與不同種類植物Med25蛋白序列一致性較高,說明該基因在植物進化過程中較保守。
使用MEGA 6軟件將橡膠樹HbMed25氨基酸序列和其他陸生植物和藻類的65個Med25的氨基酸序列進行系統進化分析,由圖4可知,65個Med25共分為6個組別(通過不同顏色區分),這些組別主要是按照植物的進化程度進行分類,如HbMed25與擬南芥(Arabidopsis thaliana)、楊樹(Populus trichocarpa)、蓖麻(Ricinus communis)、木薯(Manihot esculenta)等高等植物聚類在一起,分在第1組(Group 1)中(分別用紅色和藍色五角星形標示);黃瓜(Cucumis sativus)和胡蘿卜(Daucus carota)兩種植物分在第2組(Group 2);磯山耬斗菜(Aquilegia coerulea)單獨在第3組(Group 3);二穗短柄草(Brachypodium distachyon)和團藻(Volvox carteri)等12種植物單分在第4組(Group 4);紫萍(Spirodela polyrhiza)單獨分在第5組(Group 5);鳳梨(Ananas comosus)和卷柏(Selaginella moellendorffii)等8種植物分在第6組(Group 6)。
2.3 HbMed25基因的組織特異性表達
對橡膠樹無性系CATAS 7-33-97的側根、幼莖、葉片、雌花、 種仁、開割樹樹皮、 開割樹膠乳以及一年生萌條的形成層和初生膠乳等不同組織樣品中的HbMed25基因表達進行Real-time PCR分析,結果表明HbMed25基因在開割樹皮中表達豐度最高,在側根、雌花、開割膠乳中表達豐度較高,在幼莖、葉片和胚乳中表達豐度較低(圖5:A)。
在自然狀態下的橡膠樹一年生萌條的形成層和初生膠乳樣品中的HbMed25基因表達豐度較低,差異不大(圖5:B)。
2.4 割膠對HbMed25基因表達的影響
對橡膠樹無性系CATAS 7-33-97的未開割樹和開割成齡樹的膠乳進行比較,分析割膠對HbMed25基因表達的影響。通過Real-time PCR 結果發現,HbMed25基因割膠處理的膠乳中的表達量比未開割樹的雖高一些,但沒有達到顯著差異(圖6:A)。
為進一步驗證割膠對HbMed25基因的影響,通過統一時間割膠后,在不同時間內第二次割膠,收集第二次割膠的膠乳來研究橡膠樹的膠乳生物合成,以消除割膠過程中機械傷害對膠乳生物合成過程中的影響。Real-time PCR結果表明,割膠處理的膠乳中HbMed25基因受割膠處理上調表達,割膠后2 h顯著上調表達,在6 h時達最大值,在12 h時雖有所恢復,但仍有極顯著的上調表達,直到24 h時才逐漸恢復至對照水平,整個割膠過程中對HbMed25的基因表達模式呈正向拋物線型(圖6:B)。
為進一步了解割膠在橡膠樹不同品系中HbMed25基因表達的變化,本文對橡膠樹PPIM 600、PR 107、CATAS 7-33-97和CATAS 8-79等4個品系的未開割樹進行割膠處理,每72 h割1刀,共割5刀(第0、第1、第2、第3和第4刀),采集不同刀次的膠乳,分析割膠在橡膠樹不同品系中HbMed25基因表達的變化。Real-time PCR結果表明,除RRIM 600在第0刀的表達量最高,隨后幾刀的表達量都低于第0刀,整體呈現下調模式以外,在PR 107、CATAS 7-33-97和CATAS 8-79等3個品系的膠乳中,割膠處理的膠乳中HbMed25基因均表現出上調表達的趨勢,且大致在第1刀的表達量最高,呈現上調后下調的正向拋物線型變化(圖6:C-F)。
2.5 茉莉酸對HbMed25基因表達的影響
利用茉莉酸活性形式結構的類似物—冠菌素(COR)體外誘導橡膠樹萌條次生乳管分化實驗系統,使用COR處理橡膠樹無性系CATAS 7-33-97的一年生萌條并采集處理部位的形成層區樣品和初生膠乳樣品。Real-time PCR結果表明,HbMed25基因在COR處理后的形成層區和初生膠乳樣品中均上調表達。根據基因在形成層區和膠乳中的表達豐度、上調表達的時間點、上調表達持續時間和上調表達倍數幾個指標分析,發現以下一些規律。在形成層區和初生膠乳中的表達豐度比較,HbMed25基因在形成層區中的表達豐度低于初生膠乳(圖7)。HbMed25基因在COR處理的形成層樣品中主要在早期時間段(1~4 h)有顯著的上調表達,且在形成層材料中在1 h達最高峰值,約為對照水平的5倍,呈正向拋物線型變化(圖7:A);在COR處理的初生膠乳中HbMed25在各時間段持續高水平的上調表達,且在2 h和4 h達最高峰值,為對照水平的10~20倍,也呈現正向拋物線型變化,說明COR處理可以明顯上調HbMed25基因在橡膠樹中的表達(圖7:B)。
2.6 植物激素對HbMed25基因表達的影響
使用COR體外誘導巴西橡膠樹萌條次生乳管分化類似的實驗系統分析乙烯(ET)、赤霉素(GA)、生長素(IAA)、細胞分裂素(6-BA)等植物激素處理的初生膠乳樣品中HbMed25的基因表達。Real-time PCR結果表明,ET處理的初生膠乳樣品中,HbMed25基因最開始是下調表達,在2~8 h有明顯的上調表達,且在8 h達最高峰值,隨后又下調表達(圖8:A)。IAA處理的初生膠乳中,HbMed25基因處于下調模式,在1、8、72 h都是極顯著的下調表達,表達模式相對比較混亂,沒有明顯的規律(圖8:B)。在GA和6-BA處理的初生膠乳中,HbMed25基因的表達模式大致相似,在整個處理時間段內都是處于下調表達,且在最早期(1 h)和中期(8~72 h)都是顯著或極顯著的下調表達(圖8:C,D)。
根據基因在初生膠乳中的表達豐度、表達的時間點、上調表達持續時間和上調表達倍數幾個指標分析,發現以下的一些表達規律。在四種植物激素處理初生膠乳中的表達豐度與COR處理相比較要低很多,與未處理的未開割樹和開割樹膠乳的豐度差不多;與HbMed25基因在COR處理的初生膠乳樣品中整體的顯著上調表達且呈正向拋物線型變化,在其他四種植物激素處理的初生膠乳中,除ET處理的初生膠乳中在前期(2~8 h)有明顯上調表達外,其他植物激素處理都是下調表達,表達模式相反;與COR處理的初生膠乳中HbMed25在2 h和4 h達最高峰值,為對照水平的10~20倍,其他植物激素處理的初生膠乳中,僅ET處理的前期(2~8 h)雖有上調表達,但也未達到對照水平的2倍。可見,其他四種植物激素處理對HbMed25基因表達沒有直接作用。
3 討論與結論
本文克隆得到巴西橡膠樹HbMed25基因,通過生物信息學分析發現橡膠樹HbMed25與木薯、楊樹等高等植物的Med25相似性較高,可能編碼親水性蛋白,亞細胞定位于細胞核,這些信息與中介體復合物作用于RNA聚合酶Ⅱ的功能是吻合的。通過組織特異性表達的結果,發現HbMed25在橡膠樹的樹皮、側根和雌花中的表達豐度較高,與擬南芥(Elfving et al.,2011)、紅掌(任保蘭等,2019)的組織特異性研究的結果類似。這些結果是與Med25在根中可以調節活性氧分布、生長素的濃度、細胞壁基因表達等,以及調控根毛的發育過程(Xu & Li,2011);在花中可以調節花期、花的生成和花器官發育(Wollenerg et al.,2011);并調節獼猴桃的花期和果實的大小(郭洋濤等,2014)等方面發揮特殊功能是密切相關的。
天然橡膠是在巴西橡膠樹樹干樹皮韌皮部中的特化組織——次生乳管中合成的(Hao & Wu,2000),這些次生乳管的數量與天然橡膠的產量密切相關,是由維管形成層的紡錘狀原始細胞分化而來(Hao & Wu,2000)。我們前期研究的證據表明,茉莉酸(JA)(Hao & Wu,2000;劉惠芳等,2001)、機械傷害(Wu et al.,2002;Tian et al.,2015)和茉莉酸活性形式的類似物——冠菌素(COR)都可以誘導橡膠樹次生乳管分化,而且COR誘導橡膠樹乳管分化的效果比茉莉酸和茉莉酸甲酯(MeJA)要更強(Zhang et al.,2015)。在COR處理形成層區中,HbMed25在處理的早期表現出極顯著的上調表達,且表達模式與茉莉酸處理的樹皮中JAZ成員、MYC成員的相似(Zhao et al.,2011; Deng et al.,2018)。在模式植物擬南芥的JA信號轉導途徑中,MYC2是茉莉酸信號轉導途徑中的核心轉錄因子,調控JA反應的多個方面,包括根生長發育、機械損傷反應、抗病抗蟲等逆境脅迫反應(Kim et al.,2011; Dhawan et al.,2011;Wathugala et al.,2011)。最近研究證明,AtMed25通過與AtMYC2直接相互作用,使RNA聚合酶Ⅱ招募到AtMYC2的靶標啟動子區域來調控茉莉酸響應基因表達(Chen et al.,2012; Zhai & Li,2019; Wu et al.,2019)。后來又發現,Med25參與調解茉莉酸信號途徑的關鍵環節COI1-JAZ-MYC2,在植物處于自然狀態下,體內沒有活性形式的茉莉酸(JA-Ile)時,JA受體COI1與抑制子蛋白JAZ不能發生互作,JAZ蛋白相對穩定,并與MYC2相互作用以抑制MYC2的轉錄活性。在此過程中,JAZ蛋白與Med25處于競爭關系,競爭結合MYC2的結合位點,此時的Med25與MYC2間的相互作用相對較弱;此外,Med25還可以與JA受體COI1直接發生相互作用,將COI1招募到JA的核心轉錄因子MYC2的靶標基因啟動子區域。可見,轉錄中介體Med25在茉莉酸信號轉導途徑中起重要調控作用。據此推測Med25在JA橡膠樹次生乳管分化的進程中也起重要的調控作用,而HbMed25在COR處理的早期就表現出極顯著的上調表達,可以印證HbMed25參與了橡膠樹體內的JA信號傳導途徑調控。
外施MeJA上調橡膠合成相關基因及增強橡膠合成的作用是相似的(Deng et al.,2018),并能夠增加橡膠產量(楊署光等,2019)。乙烯利作為最有效的排膠刺激劑,能顯著延長排膠時間,增加膠乳產量(史敏晶等,2015),但與橡膠生物合成沒有直接關系(Deng et al.,2018)。割膠和JA促進橡膠樹次生乳管分化和膠乳再生(郝秉中和吳繼林,1982),割膠可以引起兩方面的反應,即機械傷害反應和排膠導致膨壓的變化。機械傷害會誘導乙烯生物合成關鍵酶基因ACS的表達,合成大量內源乙烯(Argueso et al.,2007),機械傷害和膨壓的變化會產生內源JA(Howe & Schilmiler,2002)。目前,割膠引起膠乳產量增加是ET還是JA導致的還不甚明了。ET是一種強刺激劑,ET處理會引起諸多反應,如ET促進HEVB7、44KD蛋白等膠乳凝集因子的增加,排膠時間延長(Shi et al.,2016);ET提高水通道蛋白基因HbPIP的表達,增強水分向乳管細胞的運輸,增加膠乳的水稀釋效應,增加膠乳產量等(An et al.,2015)。HbMed25是轉錄中介體基因,可以在組織發育、器官形成、植物抗逆性等方面起重要調控作用,HbMed25很有可能參與膠乳凝集反應、水分調節等進程,但目前尚無相關研究驗證。
從本文的HbMed25基因表達的模式發現,HbMed25在割膠處理膠乳中上調表達,且在產量相對較高的橡膠樹品系(CATAS 8-79和CATAS 7-33-97)的膠乳中也呈現上調表達模式,因此可以看出割膠是促進HbMed25表達的。在JA活性形式結構類似物——COR處理的初生膠乳中,HbMed25在早期就表現出極顯著的上調表達模式,與茉莉酸處理的膠乳樣品中HbCOI1和HbJAZ1成員上調表達模式相似(Deng et al.,2018; Howe et al.,2019),而在ET處理的膠乳中HbMed25基因基本處于下調表達。從基因表達的豐度方面分析,HbMed25在割膠、COR處理膠乳中的表達豐度也遠遠高于ET處理。最近,我們發現割膠樹的乳管細胞茉莉酸含量顯著高于未開割樹,割膠促進天然橡膠生物合成與激活乳管細胞中的茉莉酸信號途徑密切相關(Deng et al.,2018;楊署光等,2019)。因此,我們推測割膠促進橡膠生物合成的效應大部分來自于機械傷害誘導的內源JA的功效,而非內源ET的作用。
由此可見,HbMed25是極有可能參與了橡膠樹茉莉酸信號轉導途徑調控橡膠樹的次生乳管分化和橡膠生物合成反應。下一步,我們將對Med25與JA信號途徑的關鍵環節成員COI1-JAZ-MYC2進行蛋白互作分析,為闡明巴西橡膠樹次生乳管分化和橡膠生物合成的分子調控機制打下良好基礎。
參考文獻:
AN F,ZOU Z,CAI XQ,et al.,2015. Regulation of HbPIP2;3,a latex-abundant water transporter,is associated with latex dilution and yield in the rubber tree (Hevea brasiliensis Muell. Arg.)[J]. PLoS ONE,10(4):e0125595.
ARGUESO CT,HANSEN M,KIEBER JJ,2007. Regulation of ethylene biosynthesis[J]. J Plant Growth Regul,26(2):92-105.
BACKSTROM S,ELFVING N,NILSSON R,et al.,2007. Purification of a plant mediator from Arabidopsis thaliana identifies PFT1 as the Med25 subunit[J]. Mol Cell,26(5): 717-729.
BUTTERY BR,BOATMAN SG,1966. Manometric measurement of turgor pressures in laticiferous phloem tissues[J]. J Exp Bot,17(2):283-296.
CHEN R,JIANG HL,LI L,et al.,2012. The Arabidopsis mediator subunit MED25 differentially regulates jasmonate and abscisic acid signaling through interacting with the MYC2 and ABI5 transcription factors[J]. Plant Cell,24(7): 2898-2916.
DENG XM,GUO D,YANG SG,et al.,2018. Jasmonate signalling in the regulation of rubber biosynthesis in laticifer cells of rubber tree,Hevea brasiliensis [J]. J Exp Bot,69(15): 3559-3571.
DHAWAN R,LUO H,FOERSTER AM,et al.,2009. HISTONE MONOUBIQUITINATION1 interacts with a subunit of the mediator complex and regulates defense against necrotrophic fungal pathogens in Arabidopsis [J]. Plant Cell,21(3):1000-1019.
ELFVING N,DAVOINE C,BENLLOCH R,et al.,2011. The Arabidopsis thaliana Med25 mediator subunit integrates environmental cues to control plant development[J]. Proc Natl Acad Sci USA,108(20):8245-8250.
GUO YT,LIN MM,QI XJ,et al.,2014. Cloning and expression analysis of AcMED25 and its relationship with fruit growth and size in kiwifruit[J]. J Fruit Sci,31(4):558-565.[郭洋濤,林苗苗,齊秀娟,等,2014. 獼猴桃 AcMED25 克隆、表達分析及其與果實生長和大小的關系[J]. 果樹學報,31(4):558-565.]
HAO BZ,WU JL,2000. Laticifer differentiation in Hevea brasiliensis:induction by exogenous jasmonic acid and linolenicacid[J]. Ann Bot London,85(1):37-43.
HOWE GA,SCHILMILER AL,2002. Oxylipin metabolism in response to stress[J]. Curr opin plant boil,5(3):230-236.
KIM YJ,ZHENG B,YU Y,et al.,2011. The role of mediator in small and long noncoding RNA production in Arabidopsis thaliana[J]. EMBO J,30(5):814-822.
LIU HF,WU JL,HAO BZ,2001. Effect of jsamonic acid and other plant growth regulators on laticifer differentiation in Hevea brasiliensis[J]. Chin J Trop Crop,22(3):6-16.[劉惠芳,吳繼林,郝秉中,2001. 茉莉酸和其它激素對巴西橡膠樹乳管分化的協同作用[J]. 熱帶作物學報,22(3):6-16.]
QI DL,WANG XQ,ZHANG ZY,et al.,2013. Current situation of chinese natural rubber industry and development suggestions[J]. Chin J Trop Agric,33(2):79-87.[祁棟靈,王秀全,張志揚,等,2013. 中國天然橡膠產業現狀及其發展建議[J]. 熱帶農業科學,33(2):79-87.]
REN BL,NIU JH,ZHANG L,et al.,2016. Cloning and expression analysis of AaPFT1/MED25 gene from Anthurium andraeanum[J]. Mol Plant Breed,14(3):554-561.[任保蘭,牛俊海,張凌,等,2016. 紅掌基因PFT1/MED25類基因AaPFT1的克隆與表達分析[J]. 分子植物育種,14(3):554-561.]
SHI MJ,CHENG C,TIAN WM,2015. Effects of ethrel on physiological parameters of latex flow in rubber tree clones RY8-79 and PR 107[J]. Chin J Trop Crop,36(5):926-932.[史敏晶,程成,田維敏,2015. 乙烯利刺激對橡膠樹無性系RY8-79和PR 107排膠生理參數的影響[J]. 熱帶作物學報,36(5):926-932.]
SHI MJ,CAI FG,TIAN WM,2016. Ethrel-stimulated prolongation of latex flow in the rubber tree (Hevea brasiliensis Muell. Arg.):an Hevb7-like protein acts as a universal antagonist of rubber particle aggregating factors from lutoids and C-serum[J]. J Biochem,159(2):209-216.
SHI MJ,LI Y,DENG SM,et al.,2019. The formation and accumulation of protein-networks by physical interactions in the rapid occlusion of laticifer cells in rubber tree undergoing successive mechanical wounding[J]. BMC Plant Biol,19(1):8.
TANG CR,YANG M,FANG YJ,et al.,2016. The rubber tree genomereveals new insights into rubber production and species adaptation[J]. Nat Plants,2(6):16073.
TIAN WM,YANG SG,SHI MJ,et al.,2015. Mechanical wounding-induced laticifer differentiation in rubber tree:An indicative role of dehydration,hydrogen peroxide,and jasmonates[J]. J Plant Physiol,182:95-103.
WATHUGALA DL,RICHARDS SA,KNIGHT H,et al.,2011. OsSFR6 is a functional rice orthologue of sensitive to freezing-6 and can act as a regulator of COR gene expression,osmotic stress and freezing tolerance in Arabidopsis[J]. New Phytol,191(4):984-995.
WOLLENBERG AC,STRASSRE B,CERDAN PD,et al.,2008. Acceleration of flowering during shade avoidance in Arabidopsis alters the balance between flowering locus C-mediated repression and photoperiodic induction of flowering[J]. Plant Physiol,148(3):1681-1694.
WU FM,DENG L,ZHAI QZ,et al.,2019. Mediator subunit MED25 couples alternative splicing of JAZ genes with fine-tuning of jasmonate signaling[J]. Plant Cell,32:429-448.
WU JL,HAO BZ,TAN HY,2002.Wound-induced differentiation in Hevea brasiliensis shoots mediated by jasmonic acid[J]. J Rubber Res. 5(5):53-63.
XU R,LI YH,2011. Control of final organ size by mediator complex subunit 25 in Arabidopsis thaliana[J]. Development,138(20):4545-4554.
YANG P,ZHANG SX,CHEN HP,et al.,2015. Rhythmic expression of genes encoding enzymes for rubber biosynthesis in rubber trees[J]. Chin J Trop Crop,36(9):1587-1592.[楊鵬,張世鑫,陳惠萍,等,2015. 巴西橡膠樹乳管細胞中5個橡膠合成關鍵酶基因節律性表達研究[J]. 熱帶作物學報,36(9):1587-1592.]
YANG SG,ZHAO Y,CHEN YY,et al.,2019. Correlation between the expression level of genes related to jasmonate signaling and rubber yield[J]. Guihaia,39(5):641-649.[楊署光,趙悅,陳月異,等,2019. 橡膠樹茉莉酸信號途徑相關基因表達與橡膠產量的相關性[J]. 廣西植物,39(5):641-649.]
YOU YR,ZHAI QZ,AN CP,et al.,2019. LEUNIG_HOMOLOG mediates MYC2-dependent transcriptional activation in cooperation with the coactivators HAC1 and MED25[J]. Plant Cell,31:2187-2205.
ZHAI QZ,LI CY,2019. The plant mediator complex and its role in jasmonate signaling[J]. J Exp Bot,70(13):3415-3424.
ZHANG SX,TIAN WM,2015. Cross talk between cytokinin and jasmonates in regulating the secondary laticifer differentiation in rubber tree[J]. J Rubber Res,18(1):38-48.
ZHAO Y,ZHOU LM,CHEN YY,et al.,2011. MYC genes with differential responses to tapping,mechanical wounding,ethrel and methyl jasmonate in laticifers of rubber tree (Hevea brasiliensis Muell. Arg.)[J]. J Plant Physiol,168(14): 1649-1658.
(責任編輯 蔣巧媛)
