遼東櫟幼苗生長(zhǎng)對(duì)種子大小和子葉去除處理的響應(yīng)
2021-10-16 14:15:05朱雅婷王建禮閆興富張金峰
廣西植物 2021年9期
朱雅婷 王建禮 閆興富 張金峰
摘 要:該文在溫室內(nèi)盆播條件下,設(shè)置不去除子葉(對(duì)照)、去除1/4子葉(輕度子葉去除)、1/2子葉(中度子葉去除)和3/4子葉(重度子葉去除)4個(gè)處理,研究了種子大小和子葉去除處理對(duì)遼東櫟幼苗生長(zhǎng)及其形態(tài)學(xué)可塑性的影響。結(jié)果表明:(1)大種子幼苗株高、基莖、葉片數(shù)、單株葉面積、總干質(zhì)量和根冠比在所有處理均大于小種子幼苗,除株高外,上述其他參數(shù)在輕度子葉去除處理的不同大小種子幼苗間均差異顯著(P<0.05),前者總干質(zhì)量在中度子葉去除處理與后者差異也顯著(P<0.05)。大種子幼苗比葉面積和比根長(zhǎng)在各處理均大于小種子幼苗,比枝長(zhǎng)則相反。(2)隨著子葉去除強(qiáng)度增大,大、小種子幼苗株高、基莖、葉片數(shù)、單株葉面積和總干質(zhì)量等均呈減小趨勢(shì),株高、基徑、葉片數(shù)和單株葉面積在重度子葉去除處理均顯著小于對(duì)照和輕度子葉去除處理;小種子幼苗對(duì)子葉損失反應(yīng)更為敏感,其株高、葉片數(shù)、單株葉面積和總干質(zhì)量在中度子葉去除處理也顯著小于對(duì)照(P<0.05);小種子幼苗根冠比隨子葉去除強(qiáng)度增大而增大,在中度和重度子葉去除處理顯著大于對(duì)照和輕度子葉去除處理(P<0.05)。(3)隨著子葉去除強(qiáng)度增大,大小種子幼苗的比葉面積和比根長(zhǎng)均逐漸增大,但大種子幼苗比枝長(zhǎng)持續(xù)減小,輕度子葉去除處理的小種子幼苗比枝長(zhǎng)最大。
關(guān)鍵詞:遼東櫟,種子大小,子葉去除,幼苗生長(zhǎng),形態(tài)學(xué)可塑性
中圖分類(lèi)號(hào):Q948.1
文獻(xiàn)標(biāo)識(shí)碼:A
文章編號(hào):1000-3142(2021)09-1524-10
Abstract:Four treatments of cotyledon excisions (no cotyledon excision,excisions of 1/4,1/2 and 3/4 cotyledon,defined as control,light,medium,and high cotyledon excisions,respectively)were conducted to investigate the effects of seed size and different cotyledon excisions on the growth and morphological plasticity of Quercus wutaishanica seedlings planted in greenhouse pots. The results were as follows:(1)The shoot height (SH),basal stem diameter (BSD),leaf number (LN),leaf area per plant (LAPP),total dry mass (TDM),and root-shoot ratio (RSR)of Q. wutaishanica seedlings germinated from large seeds were higher than those of seedlings from small seeds in all treatments of cotyledon excision. There were significant differences in all above parameters,with the exception of SH,among seedlings from large and small seeds,on which cotyledons were lightly excised (P<0.05). Moreover,TDM of the former was significantly higher than that of the latter for the medium cotyledon excision (P<0.05). Also,higher specific leaf area (SLA)and specific root length (SRL)of seedlings from large seeds than those from small seeds were observed in all treatments of cotyledon excisions,while the specific shoot length (SSL)was detected in reverse. (2)With the intensity of cotyledon excision increasing,SH,BSD,LN,LAPP and TDM of seedlings from both large and small seeds appeared a trend of decline. SH,BSD,LN and LAPP of seedlings emerged from high cotyledon excision seeds were significantly lower than those of seedlings from control and light cotyledon excision seeds (P<0.05). Higher sensitivity to cotyledon excision was found in seedlings from small seeds than from large seeds,and SH,LN,LAPP and TDM of seedlings originated from small seeds,of which cotyledons were mediumly excised were also lower than those of seedlings from control seeds (P<0.05). RSR increased with the intensity of cotyledon excision increasing,and they were markedly higher in seedlings from seeds,regardless of whether they were large or small sized,mediumly and highly excised than those in seedlings from control and light cotyledon excision seeds (P<0.05). (3)With the intensity of cotyledon excision increasing,SLA and SRL gradually increased,while SSL of seedlings emerged from large seeds continuously decreased and the seedlings emerged from small seeds whose cotyledons were lightly excised had the maximum of SSL.
Key words:Quercus wutaishanica,seed size,cotyledon excision,seedling growth,morphological plasticity
富含營(yíng)養(yǎng)的櫟屬(Quercus)植物種子是脊椎動(dòng)物的重要食物資源,種子成熟散落后遭動(dòng)物大量取食(孫書(shū)存和陳靈芝,2001; 高賢明等,2003; 閆興富等,2011),即使在幼苗建立期間,捕食動(dòng)物也可能以幼莖為搜尋線索(Pyare & Longland,2000)而大量取食其子葉(高賢明等,2003; 閆興富等,2011,2012),從而增大其幼苗死亡的風(fēng)險(xiǎn)。植物從種子到幼苗的轉(zhuǎn)變是其生活史中對(duì)環(huán)境因子反應(yīng)最為敏感的階段,在從種子萌發(fā)開(kāi)始的幼苗建植過(guò)程中,種子貯藏的營(yíng)養(yǎng)物質(zhì)為種子萌發(fā)、幼苗生長(zhǎng)和莖葉損傷后再生所需營(yíng)養(yǎng)提供了保障(閆興富等,2012)。有研究認(rèn)為種子遭受取食危害后嚴(yán)重影響其萌發(fā)及其幼苗生長(zhǎng)(Lombardo & McCarthy,2009; 劉艷等,2013; 劉影等,2017; 姜曉旭等,2018),特別是高強(qiáng)度子葉損失可減少幼苗根莖生物量積累(Shi et al.,2017),甚至導(dǎo)致幼苗建植失敗(Mancilla-Leytón et al.,2013),但輕度子葉損失不會(huì)影響種子萌發(fā)率(Mancilla-Leytón et al.,2012),且可通過(guò)加快滲透(Finch-Savage & Clay,1994)或促進(jìn)IAA等生長(zhǎng)調(diào)節(jié)物質(zhì)合成(Prewein et al.,2006)而促進(jìn)種子快速萌發(fā)和同步出苗(Hou et al.,2010; Giertych & Suszka,2011; Perea et al.,2011; Shi et al.,2017)。子葉部分損失對(duì)幼苗生長(zhǎng)發(fā)育無(wú)顯著影響(Giertych & Suszka,2011),因?yàn)樽尤~中的營(yíng)養(yǎng)物質(zhì)在幼苗建立過(guò)程中已大量轉(zhuǎn)移并貯藏于主根(Giertych & Suszka,2011; 閆興富等,2012; 姜曉旭等,2018),而這些營(yíng)養(yǎng)物質(zhì)有助于幼苗在子葉或莖葉受損后的生長(zhǎng)恢復(fù)和新芽萌生,可增大幼苗在林下成功建植的機(jī)會(huì)(Perea et al.,2011; 閆興富等,2012)。
種子子葉損失對(duì)幼苗生長(zhǎng)的影響可能與種子大小(Yi et al.,2015; Lebrija-Trejos et al.,2016)、子葉受損程度(Giertych & Suszka,2011; 姜曉旭等,2018)和損失部位(Hou et al.,2010)等因素有關(guān),尤其是種群內(nèi)和種群間甚至同一植株內(nèi)普遍存在的種子大小差異(朱金雷和劉志民,2012)可能對(duì)子葉損失后幼苗的生長(zhǎng)發(fā)育具有重要影響。貯藏更多營(yíng)養(yǎng)物質(zhì)的大種子萌發(fā)建立幼苗的機(jī)會(huì)更大(黃雙全等,2007),且具有更強(qiáng)的耐受惡劣環(huán)境能力和對(duì)光照和養(yǎng)分等資源的競(jìng)爭(zhēng)能力(Lebrija-Trejos et al.,2016)。因?yàn)榉N子營(yíng)養(yǎng)物質(zhì)向幼苗的轉(zhuǎn)移在其早期發(fā)育過(guò)程中具有重要作用,所以,大種子萌發(fā)幼苗壽命更長(zhǎng),生物量積累更多(Jrgensen et al.,2019),而且大種子貯藏的營(yíng)養(yǎng)物質(zhì)遠(yuǎn)遠(yuǎn)多于種子萌發(fā)和幼苗生長(zhǎng)的營(yíng)養(yǎng)需要(Yi et al.,2015)。有研究認(rèn)為,較大種子的更多營(yíng)養(yǎng)物質(zhì)不僅作為種子萌發(fā)和幼苗建立的營(yíng)養(yǎng)來(lái)源(Yi et al.,2015),而且在大多情況下,動(dòng)物是從距胚根更遠(yuǎn)的子葉末端開(kāi)始消耗營(yíng)養(yǎng)物質(zhì),這些被消耗的營(yíng)養(yǎng)物質(zhì)可能是植物對(duì)捕食動(dòng)物的飽和作用或?yàn)閿U(kuò)散動(dòng)物提供食物“回報(bào)”而額外儲(chǔ)備的冗余營(yíng)養(yǎng)(Perea et al.,2011; Yi et al.,2015),動(dòng)物對(duì)種子的這一取食行為還與Prewein et al. (2006)報(bào)道的“櫟屬植物種子子葉近胚軸端單寧含量相對(duì)較高”和Hou et al. (2010)報(bào)道的“子葉頂端的營(yíng)養(yǎng)物質(zhì)對(duì)維持種子活力和幼苗建立更為重要”的種子結(jié)構(gòu)相關(guān)特征高度吻合。因此,相同的子葉損失量對(duì)較大種子萌發(fā)幼苗生長(zhǎng)發(fā)育的負(fù)面影響可能相對(duì)較小(Yi et al.,2015)。
遼東櫟(Quercus wutaishanica)是我國(guó)暖溫帶落葉林的重要優(yōu)勢(shì)樹(shù)種之一,其種子在秋季成熟散落后立即萌發(fā)并將子葉的大部分營(yíng)養(yǎng)物質(zhì)快速轉(zhuǎn)移至主根,胚芽在翌年春季伸長(zhǎng)形成幼苗(孫書(shū)存和陳靈芝,2001; 高賢明等,2003),這一特殊的萌發(fā)和幼苗建立方式不僅降低了種子脫水死亡和被動(dòng)物捕食的風(fēng)險(xiǎn),而且即使在幼苗子葉遭動(dòng)物捕食的情況下仍能正常存活與生長(zhǎng)(閆興富等,2011,2012)。已有研究結(jié)果顯示,即使在結(jié)實(shí)大年遼東櫟種子也會(huì)因捕食動(dòng)物取食而大量損失(孫書(shū)存和陳靈芝,2001; 閆興富等,2011),但其種子結(jié)實(shí)量在不同年份間的明顯波動(dòng)(孫書(shū)存和陳靈芝,2001; 程積民等,2009)和種子大小變異(程積民等,2009; 閆興富等,2011)在種群水平(Wang et al.,2000)和種子水平上(Yi & Yang,2010)對(duì)捕食動(dòng)物的飽和效應(yīng)可使動(dòng)物捕食危害得以緩解。然而,種子萌發(fā)后遼東櫟幼苗的子葉仍面臨較高動(dòng)物捕食風(fēng)險(xiǎn)(閆興富等,2011,2012),已有研究采用人工切除子葉的方式模擬研究不同程度動(dòng)物取食子葉對(duì)種子萌發(fā)和幼苗早期生長(zhǎng)的影響(閆興富等,2014),但其使用的種子大小較為均一,且幼苗生長(zhǎng)時(shí)間相對(duì)較短,而大小各異的種子在去除子葉后對(duì)幼苗生長(zhǎng)的影響仍值得進(jìn)一步探索。本研究以不同大小的遼東櫟種子為材料,在盆播條件下研究不同程度人工去除子葉對(duì)幼苗生長(zhǎng)的影響,研究結(jié)果有助于進(jìn)一步深入了解捕食動(dòng)物取食種子危害對(duì)遼東櫟幼苗生長(zhǎng)和種群實(shí)生更新的影響機(jī)制,并為森林培育和退化植被恢復(fù)實(shí)踐中的種苗繁育提供參考。
1 材料與方法
1.1 種子的采集和處理
實(shí)驗(yàn)用遼東櫟種子采自寧夏六盤(pán)山林業(yè)局所屬龍?zhí)读謭?chǎng)的大倒溝林區(qū)樹(shù)齡約25年以上遼東櫟灌叢植株。于2017年9月上旬,在采種林區(qū)選擇相對(duì)較大(株高2 m以上)、冠層生長(zhǎng)健壯的灌叢植株,在結(jié)果集中的枝條上摘取足夠數(shù)量的不同大小的成熟遼東櫟種子,大、小種子鮮重分別為(3.05±0.38)g (n=100)和(1.46±0.27)g (n=100)。于采摘次日帶回實(shí)驗(yàn)室,以水浸法除去昆蟲(chóng)浸染的種子,于陰涼處晾干種子表面水分并經(jīng)短期貯藏(約1周)后用于播種和實(shí)驗(yàn)幼苗的繁育。
1.2 研究方案和幼苗繁育
為避免冬季和早春低溫造成幼苗死亡,盆栽實(shí)驗(yàn)在玻璃溫室內(nèi)進(jìn)行,溫室內(nèi)光照強(qiáng)度約為自然全光照的55%,溫室在冬季以普通暖氣片供暖(11月1日—3月31日),白天溫度約(23±5)℃,夜間溫度約(16±3)℃;考慮到遼東櫟幼苗發(fā)育早期喜蔭的特點(diǎn),在溫室內(nèi)用單層黑色尼龍網(wǎng)眼布進(jìn)行遮陰,遮陰棚下相對(duì)光強(qiáng)約為自然全光照的18.9%。于2017年9月20日,取32個(gè)高23.5 cm、內(nèi)徑20 cm的塑料花盆,等分為2組,分別播種大種子和小種子,每組的16盆再分成4個(gè)小組(4盆為1小組)分別作為大種子和小種子幼苗的4個(gè)不同程度子葉去除處理,各播入大、小種子的4盆分別作為大、小種子幼苗的4次重復(fù)。取上述處理好的大、小遼東櫟種子各480粒,將種子用75%乙醇進(jìn)行表面消毒和清水沖洗后,自種子基部一端用單面刀片劃開(kāi)并去除種皮,其中1組種子(大、小種子各120粒)直接播種于花盆中作為對(duì)照(不去除子葉處理),其余3組分別為去除子葉1/4/、1/2和3/4 3個(gè)處理,用單面刀片分別將3個(gè)處理的種子切除子葉基部長(zhǎng)度的1/4、1/2和3/4,上述4組處理分別定義為對(duì)照、輕度、中度和重度子葉去除處理。子葉去除處理后立即播入上述預(yù)先準(zhǔn)備好的花盆中。
播種前先將經(jīng)多年種植小麥?zhǔn)旎娘L(fēng)沙土裝入花盆并充分澆水,種子子葉去除處理完成后立即進(jìn)行播種;按上述方案在每一花盆內(nèi)播入對(duì)應(yīng)子葉去除處理的種子30粒,將種子胚根一端朝下播入土中,播種后覆蓋厚約1 cm的濕潤(rùn)沙土。播種后約1周左右胚根伸長(zhǎng)萌發(fā),翌年4月初胚芽伸長(zhǎng)出苗,約6周后出苗結(jié)束。出苗結(jié)束2周后,每盆選留生長(zhǎng)健壯、植株大小基本一致的幼苗15株定苗,澆水后連根挖除多余幼苗。試驗(yàn)期間適時(shí)澆水保持盆內(nèi)土壤濕潤(rùn)以滿足種子萌發(fā)和幼苗生長(zhǎng)對(duì)水分的需要,并定期進(jìn)行松土、除草等日常管理。
1.3 幼苗收獲和生長(zhǎng)參數(shù)的測(cè)定與計(jì)算
2018年10月20日,收獲上述各花盆的全部幼苗,花盆充分澆水后小心將幼苗從土中連根挖出,用自來(lái)水清洗干凈后帶回實(shí)驗(yàn)室,用潔凈濾紙吸干表面水珠后測(cè)定記錄幼苗的株高(shoot height,SH)、基徑(basal stem diameter,BSD,用電子數(shù)顯式游標(biāo)卡尺測(cè)定)、葉片數(shù)(leaf number,LN )、單株葉面積(leaf area per plant,LAPP,用美國(guó)產(chǎn)LI-3100葉面積測(cè)定儀測(cè)定);將幼苗分根、莖和葉片裝入紙袋中置于數(shù)字顯示電子烘干箱中烘干(控制在85 ℃條件下),48 h后用電子天平分別稱取根、莖和葉干質(zhì)量;根據(jù)上述測(cè)定的數(shù)據(jù)計(jì)算幼苗的總干質(zhì)量(total dry mass,TDM)、根冠比(root shoot ratio,RSR)、比葉面積(specific leaf area,SLA)、比根長(zhǎng)(special root length,SRL)和比枝長(zhǎng)(special shoot length,SSL)。相關(guān)參數(shù)的計(jì)算公式如下:TDM=根干質(zhì)量+莖干質(zhì)量+葉干質(zhì)量;RSR=根干質(zhì)量/(莖干質(zhì)量+葉干質(zhì)量);SLA=LAPP/葉干質(zhì)量(cm2·g-1);SRL=主根長(zhǎng)/主根干質(zhì)量(cm·g-1);SSL=主莖長(zhǎng)/主莖干質(zhì)量(cm·g-1)。
1.4 數(shù)據(jù)統(tǒng)計(jì)分析
所有實(shí)驗(yàn)數(shù)據(jù)在統(tǒng)計(jì)分析前經(jīng)平方根轉(zhuǎn)換,用兩因素方差分析法(two-way ANOVA)分析種子大小和子葉去除處理及其交互作用對(duì)幼苗生長(zhǎng)參數(shù)是否具有顯著影響,用最小顯著差異法(LSD)分析幼苗生長(zhǎng)參數(shù)在不同大小種子幼苗間和不同子葉去除處理間的差異顯著性。全部數(shù)據(jù)統(tǒng)計(jì)分析均在SPSS 13.0中進(jìn)行,結(jié)果中的數(shù)據(jù)以平均值呈現(xiàn)。
2 結(jié)果與分析
2.1 子葉去除對(duì)不同大小種子萌發(fā)幼苗生長(zhǎng)的影響
種子大小對(duì)幼苗基莖、葉片數(shù)和單株葉面積的影響均達(dá)極顯著水平,但對(duì)株高無(wú)顯著影響;子葉去除處理對(duì)上述參數(shù)均具有極顯著影響,但種子大小和子葉去除處理的交互作用對(duì)上述4個(gè)生長(zhǎng)參數(shù)的影響均不顯著(表1)。
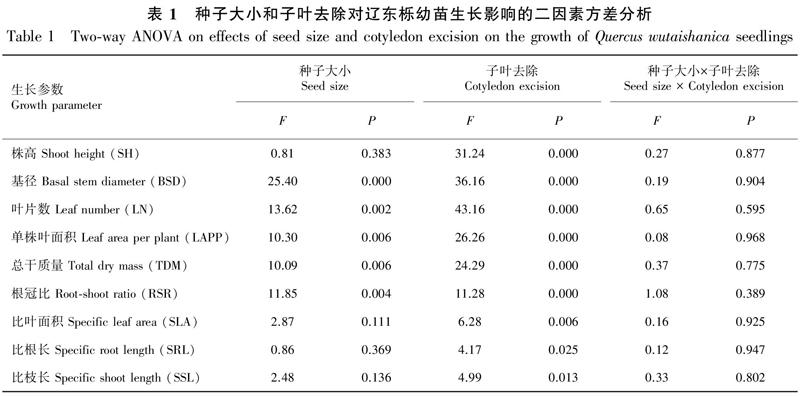
從圖1可以看出,不同子葉去除處理的大種子幼苗株高均不同程度大于小種子幼苗,但大、小種子幼苗間株高差異均不顯著;大、小種子幼苗株高在對(duì)照中均為最大(分別為14.40、13.37 cm),隨子葉去除強(qiáng)度增大都依次減小,其中大種子幼苗在重度、小種子幼苗在中度和重度子葉去除處理的株高均顯著小于對(duì)照(P<0.05),小種子幼苗在3個(gè)子葉去除處理間的差異也均達(dá)顯著水平(P<0.05)。大種子幼苗基徑均大于小種子幼苗,且對(duì)照和輕度子葉去除處理的不同大小種子幼苗間差異顯著(P<0.05);不同大小種子幼苗基徑都隨子葉去除強(qiáng)度增大而減小,大種子幼苗在不同子葉去除處理及其與對(duì)照間差異均顯著(P<0.05),小種子幼苗僅在重度子葉去除與對(duì)照、輕度子葉去除處理間差異顯著(P<0.05)。
大種子幼苗葉片數(shù)在各處理都大于小種子幼苗,且前者僅在對(duì)照和輕度子葉去除處理顯著大于后者(P<0.05);不同大小種子幼苗葉片數(shù)在對(duì)照中均為最大,3個(gè)子葉去除處理的大種子幼苗葉片數(shù)均顯著減小(P<0.05),重度子葉去除處理還顯著小于輕度子葉去除處理(P<0.05),小種子幼苗在不同子葉去除處理及其與對(duì)照間均差異顯著(P<0.05)。大種子幼苗單株葉面積也都大于小種子幼苗,前者在輕度子葉去除處理顯著大于后者(P<0.05);大種子幼苗單株葉面積在中度和重度子葉去除處理均顯著小于對(duì)照和輕度子葉去除處理(P<0.05),而小種子幼苗除在中度和重度子葉去除處理間無(wú)顯著差異外,其他處理及其與對(duì)照間均差異顯著(P<0.05)。
2.2 幼苗生物量積累和形態(tài)學(xué)可塑性
2.2.1 幼苗生物量積累及分配 種子大小和子葉去除均顯著影響幼苗總干質(zhì)量和根冠比,但兩者的交互作用對(duì)幼苗總干質(zhì)量及其分配(根冠比)無(wú)顯著影響(表1)。
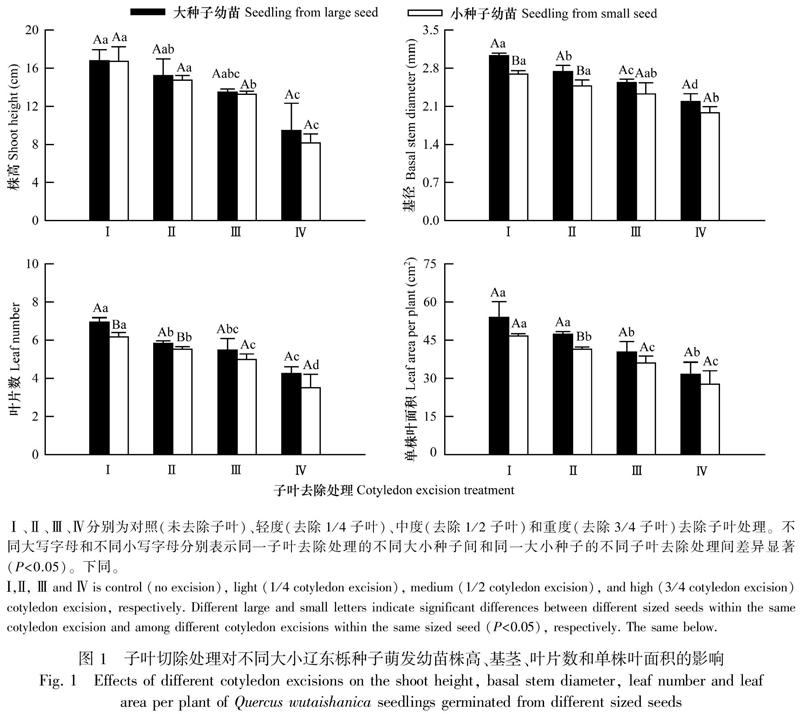
從圖2可以看出,大種子幼苗總干質(zhì)量均高于小種子幼苗,輕度和中度子葉去除處理的不同大小種子幼苗間差異顯著(P<0.05);隨著子葉去除強(qiáng)度增大,大、小種子幼苗總干質(zhì)量分別從對(duì)照的0.53、0.44 g減小至0.37、0.34 g,其中大種子幼苗除在輕度和中度子葉去除處理間無(wú)顯著差異外,其他處理及其與對(duì)照幼苗間差異均顯著(P<0.05),小種子幼苗總干質(zhì)量在3個(gè)子葉去除處理均顯著小于對(duì)照(P<0.05),中度和重度子葉處理間差異均顯著(P<0.05)。大種子幼苗根冠比在對(duì)照和3個(gè)不同強(qiáng)度子葉去除處理均大于小種子幼苗,不同大小種子幼苗間差異不顯著;子葉去除增大了幼苗根冠比,大、小種子幼苗根冠比均在重度子葉去除處理最大(分別為0.73、0.68),其中,前者在輕度子葉去除處理顯著大于對(duì)照(P<0.05),后者除在輕度子葉去除處理與對(duì)照間無(wú)顯著差異外,其他處理及其與對(duì)照間差異均顯著(P<0.05)。
2.2.2 幼苗形態(tài)學(xué)可塑性 種子大小對(duì)幼苗比葉面積、比根長(zhǎng)和比枝長(zhǎng)均無(wú)顯著影響,而子葉去除處理對(duì)這些參數(shù)影響均顯著,兩者的交互作用對(duì)上述3個(gè)參數(shù)無(wú)顯著影響(表1)。從圖3可以看出,大種子幼苗比葉面積在不同子葉去除處理均大于小種子幼苗,差異不顯著;大、小種子幼苗比葉面積均在對(duì)照最小(分別為244.53、220.47 cm2·g-1),且均隨子葉去除強(qiáng)度增大逐漸增大,在重度子葉去除處理最大(分別為364.07、307.66 cm2·g-1),其中,小種子幼苗在重度子葉去除處理顯著大于對(duì)照(P<0.05)。
大種子幼苗比根長(zhǎng)在對(duì)照和不同強(qiáng)度子葉去除處理均大于小種子幼苗,不同大小種子幼苗間差異不顯著;隨著子葉去除強(qiáng)度增大,大、小種子幼苗比根長(zhǎng)分別從對(duì)照的86.15、76.42 cm·g-1逐漸增大至重度子葉去除處理的114.39、109.95 cm·g-1,各處理及其與對(duì)照間無(wú)顯著差異。對(duì)照和不同強(qiáng)度子葉去除處理大種子幼苗比枝長(zhǎng)均小于小種子幼苗,差異不顯著;大種子幼苗比枝長(zhǎng)隨著子葉去除強(qiáng)度增大從對(duì)照的最大值(100.73 cm·g-1)逐漸減小至重度子葉去除處理的73.62 cm·g-1,但僅有中度子葉去除處理顯著小于對(duì)照(P<0.05),小種子幼苗在輕度子葉去除處理略大于對(duì)照,但在中度和重度子葉去除處理持續(xù)減小,各子葉去除處理及其與對(duì)照間差異均不顯著。
3 討論與結(jié)論
植物種群間、種群內(nèi)甚至同一植物個(gè)體內(nèi)部普遍存在種子大小的差異(朱金雷和劉志民,2012)。與小種子相比,大種子因含有較多營(yíng)養(yǎng)物質(zhì)而被種子擴(kuò)散者有效貯藏并建立幼苗的機(jī)會(huì)更大,且大種子幼苗對(duì)不利環(huán)境條件的耐受能力以及對(duì)光照和養(yǎng)分等資源的競(jìng)爭(zhēng)力更強(qiáng)(Lebrija-Trejos et al.,2016)。武高林和杜國(guó)禎(2006)認(rèn)為幼苗早期階段生長(zhǎng)與初始大小密切相關(guān),而幼苗初始大小可能直接取決于種子大小差異,因?yàn)樵诿劝l(fā)過(guò)程中種子營(yíng)養(yǎng)物質(zhì)可轉(zhuǎn)移并貯藏于幼苗中,從而使大種子幼苗發(fā)育出比小種子幼苗更為完善的根系(李榮平等,2004; 閆興富等,2012)。本研究發(fā)現(xiàn),遼東櫟大種子幼苗株高、基莖、葉片數(shù)和單株葉面積等生長(zhǎng)參數(shù)在各子葉去除處理均不同程度地大于小種子幼苗,其中輕度子葉去除處理幼苗基莖、葉片數(shù)和單株葉面積在不同大小種子幼苗間差異顯著,而中度和重度子葉去除處理在不同大小種子幼苗間差異不顯著。這些結(jié)果表明,遼東櫟種子大小是影響幼苗生長(zhǎng)的重要因素。一方面大種子借助其對(duì)捕食動(dòng)物的飽和效應(yīng)(Yi & Yang,2010)優(yōu)勢(shì)而具有比小種子更強(qiáng)的耐受動(dòng)物取食危害能力;另一方面,大種子可通過(guò)其特殊的萌發(fā)方式發(fā)育出更為完善的主根,從而具有比小種子幼苗更強(qiáng)的耐受不良環(huán)境能力和資源競(jìng)爭(zhēng)等方面的優(yōu)勢(shì)(Lebrija-Trejos et al.,2016)。
種子大小對(duì)幼苗存活的影響可能主要與幼苗早期階段生物量分配模式和資源利用方式有關(guān)(武高林和杜國(guó)禎,2008)。本研究中,遼東櫟大種子幼苗總干質(zhì)量和根冠比在對(duì)照和不同強(qiáng)度子葉去除處理都高于小種子幼苗,且總干質(zhì)量在輕度和中度子葉去除處理的不同大小種子幼苗間差異顯著。幼苗這一干物質(zhì)積累和分配模式可能與遼東櫟種子秋季胚根快速伸長(zhǎng)而胚芽翌年出土的出苗方式有關(guān)。一方面,處于萌發(fā)階段的種子盡可能多地將營(yíng)養(yǎng)轉(zhuǎn)移至主根以減少子葉被動(dòng)物取食造成的營(yíng)養(yǎng)損失(閆興富等,2012),從而使大種子幼苗積累干物質(zhì)更多(何彥龍等,2007; Jrgensen et al.,2019),尤其在高強(qiáng)度子葉損失情況下,具有比小種子幼苗更為明顯的干物質(zhì)積累優(yōu)勢(shì)(Yi et al.,2015);另一方面,隨著幼苗生長(zhǎng)后期源—庫(kù)關(guān)系轉(zhuǎn)化和對(duì)種子大小依賴的消失或減弱(何彥龍等,2007; Shi et al.,2017),對(duì)相對(duì)生長(zhǎng)率較低的大種子幼苗來(lái)說(shuō),其后期地上部分生長(zhǎng)可能主要依賴于土壤養(yǎng)分資源,生長(zhǎng)優(yōu)勢(shì)逐漸減弱(何彥龍等,2007; 黃雙全等,2007),因而其根冠比大于小種子幼苗。本研究發(fā)現(xiàn),盡管種子大小對(duì)遼東櫟幼苗比葉面積、比根長(zhǎng)和比枝長(zhǎng)影響不顯著,但大種子幼苗比葉面積和比根長(zhǎng)在對(duì)照和不同強(qiáng)度子葉去除處理均大于小種子幼苗,而比枝長(zhǎng)則相反,表明不同大小種子幼苗形態(tài)學(xué)可塑性存在一定差異,何彥龍等(2007)也曾報(bào)道,草本植物黃帚橐吾(Ligularia virgaurea)不同大小種子幼苗比葉面積并無(wú)明顯差異,比葉面積變化作為植物維持最優(yōu)捕獲光能能力的一種機(jī)制,可通過(guò)增大單位葉面積光能捕獲量而提高光能利用率和相對(duì)生長(zhǎng)率。本研究結(jié)果與一些研究報(bào)道的“小種子幼苗相對(duì)生長(zhǎng)率較高”(Wu et al.,2006; 何彥龍等,2007)的結(jié)論有較大出入,較小的比葉面積和相對(duì)生長(zhǎng)率可能有助于小種子幼苗通過(guò)維持更大的碳水化合物和礦質(zhì)資源庫(kù)來(lái)增強(qiáng)對(duì)環(huán)境脅迫的耐受能力(Westoby et al.,1992),而大種子幼苗較高的相對(duì)生長(zhǎng)率可能導(dǎo)致生長(zhǎng)后期主根營(yíng)養(yǎng)過(guò)度透支而變得纖細(xì),從而表現(xiàn)為比根長(zhǎng)增大的形態(tài)學(xué)可塑性變化(許中旗等,2009)。小種子幼苗比枝長(zhǎng)較大可能是其在發(fā)育早期采取的一種優(yōu)先保障地上部分營(yíng)養(yǎng)供應(yīng)的策略,但發(fā)育后期因營(yíng)養(yǎng)枯竭變得“消瘦”而增大比枝長(zhǎng),有關(guān)青藏高原高寒草甸植物的研究也報(bào)道了小種子物種幼苗具有更大形態(tài)可塑性的結(jié)論(Wu et al.,2006),但該結(jié)果與本研究的幼苗比葉面積和比根長(zhǎng)的結(jié)論并不一致,產(chǎn)生這些結(jié)果差異的原因還有待于進(jìn)一步研究。
櫟屬植物種子子葉是幼苗發(fā)育的主要營(yíng)養(yǎng)物質(zhì)來(lái)源,在種子萌發(fā)和幼苗的建立、生長(zhǎng)與存活中都具有重要作用(高賢明等,2003; 閆興富等,2012),較大種子能夠耐受一定程度子葉損失仍可建立幼苗(Fukumoto & Kajimura,2000; Giertych & Suszka,2011),而且適度子葉損失可能會(huì)減少種子內(nèi)多酚類(lèi)生長(zhǎng)抑制物質(zhì)向幼苗轉(zhuǎn)移而促進(jìn)幼苗生長(zhǎng)(劉艷等,2013),因?yàn)闄祵僦参锓N子的營(yíng)養(yǎng)貯藏遠(yuǎn)大于幼苗生長(zhǎng)發(fā)育的需要,甚至大種子的冗余營(yíng)養(yǎng)物質(zhì)還可通過(guò)快速萌發(fā)轉(zhuǎn)移至主根后以殘留的子葉“誘使”捕食動(dòng)物“優(yōu)先”將其取食(高賢明等,2003; 閆興富等,2012; Yi et al.,2015),從而降低整個(gè)種子損失的風(fēng)險(xiǎn)。因此,人工去除部分子葉可作為一種促進(jìn)種子整齊萌發(fā)的技術(shù)應(yīng)用于容器苗生產(chǎn)實(shí)踐(Mancilla-Leytón et al.,2013; 史文輝等,2018; Shi et al.,2018)。然而,高強(qiáng)度子葉損失顯著影響幼苗生長(zhǎng)和根系發(fā)育,并對(duì)后續(xù)的土壤養(yǎng)分和水分吸收產(chǎn)生影響(Giertych & Suszka,2011; Mancilla-Leytón et al.,2013),甚至有研究報(bào)道Pasania edulis幼苗的生長(zhǎng)受昆蟲(chóng)取食種子危害的負(fù)面影響可持續(xù)3年之久(Sone et al.,2016)。本研究中,隨著子葉去除強(qiáng)度增大,遼東櫟大、小種子幼苗株高、基莖、葉片數(shù)和單株葉面積等均呈減小趨勢(shì),且在重度子葉去除處理均顯著小于對(duì)照和輕度子葉去除處理,尤其小種子幼苗株高、葉片數(shù)和單株葉面積在中度子葉去除處理均顯著小于對(duì)照,表明高強(qiáng)度子葉損失嚴(yán)重影響幼苗存活與生長(zhǎng)(劉艷等,2013; 閆興富等,2014; 姜曉旭等,2018),甚至有研究發(fā)現(xiàn)高強(qiáng)度子葉損失可影響幼苗第二個(gè)生長(zhǎng)季節(jié)的株高生長(zhǎng)和干物質(zhì)積累(Giertych & Suszka,2011)。因此,櫟屬植物種子耐受子葉損失的量可能有一個(gè)臨界值,而且不同大小種子的臨界值存在差異。
研究發(fā)現(xiàn),隨著子葉去除強(qiáng)度增大,遼東櫟不同大小種子幼苗總干質(zhì)量均逐漸減小,根冠比則逐漸增大;總干質(zhì)量在不同強(qiáng)度子葉去除處理均顯著小于對(duì)照,小種子幼苗生物量分配對(duì)子葉去除處理的反應(yīng)比大種子幼苗更為敏感,尤其根冠比在中度和重度子葉去除處理均顯著大于對(duì)照,而大種子幼苗僅在輕度子葉去除處理與對(duì)照間差異顯著。一方面,低強(qiáng)度子葉損失不同程度地抑制幼苗莖、葉生物量和總生物量積累,對(duì)根冠比影響較小(劉影等,2017),但中度和重度子葉損失導(dǎo)致幼苗莖、葉生物量及總生物量積累顯著減少(Perea et al.,2011; 閆興富等,2014; 劉影等,2017),甚至導(dǎo)致幼苗建植失敗(Mancilla-Leytón et al.,2013);另一方面,幼苗子葉部分損失后傾向于優(yōu)先將更多營(yíng)養(yǎng)物質(zhì)分配至根系還可增強(qiáng)幼苗莖葉的萌生更新能力(Shi et al.,2017; Yi et al.,2019)和幼苗發(fā)育過(guò)程中對(duì)環(huán)境風(fēng)險(xiǎn)的防御能力(閆興富等,2012)。遼東櫟主根干物質(zhì)積累對(duì)種子子葉的依賴及幼苗可能在種子萌發(fā)期間建立起上述防御機(jī)制,其特殊的萌發(fā)和出苗方式既有利于幼苗營(yíng)養(yǎng)物質(zhì)分配在秋季和春季之間進(jìn)行時(shí)間上的權(quán)衡,又有利于在主根和留土子葉之間進(jìn)行空間上的權(quán)衡,從而確保幼苗莖桿發(fā)育的營(yíng)養(yǎng)來(lái)源在子葉遭動(dòng)物取食損失后轉(zhuǎn)向依賴主根貯藏的營(yíng)養(yǎng)物質(zhì)(高賢明等,2003; 閆興富等,2011,2012)。事實(shí)上,捕食動(dòng)物對(duì)遼東櫟幼苗子葉取食的同時(shí),往往連同頂芽一起取食(閆興富等,2012),主根之所以能穩(wěn)穩(wěn)地“鉚”在土壤中完全得益于其從子葉獲取的大量營(yíng)養(yǎng)物質(zhì)。因此,遼東櫟子葉受損幼苗根冠比增大可能是其應(yīng)對(duì)子葉營(yíng)養(yǎng)損失的一種重要機(jī)制,而這些貯藏于主根中的營(yíng)養(yǎng)物質(zhì)也為幼苗頂芽受損(包括幼苗發(fā)育期間莖葉受損)后的萌生提供了營(yíng)養(yǎng)保障(閆興富等,2012)。
遼東櫟幼苗可對(duì)子葉損失引起的營(yíng)養(yǎng)虧缺表現(xiàn)出一定的形態(tài)學(xué)可塑性(閆興富等,2014),隨著子葉去除強(qiáng)度增大,幼苗比葉面積和比根長(zhǎng)均逐漸增大,大種子幼苗比枝長(zhǎng)持續(xù)減小,而小種子幼苗比枝長(zhǎng)在輕度子葉去除處理略有增大,表明幼苗對(duì)子葉損失引起的營(yíng)養(yǎng)虧缺能夠在形態(tài)學(xué)方面做出有利于提高資源利用效率的補(bǔ)償性響應(yīng),即通過(guò)增大比葉面積來(lái)提高光能利用率,或通過(guò)增大比根長(zhǎng)來(lái)增加根系對(duì)土壤深層養(yǎng)分和水分的吸收(Westoby et al.,1992)。有研究認(rèn)為,子葉損失對(duì)幼苗地上部分營(yíng)養(yǎng)的限制可能更大(Giertych & Suszka,2011),從而加劇胚根在種子萌發(fā)期間快速伸長(zhǎng)對(duì)營(yíng)養(yǎng)的需求與地上部分間的競(jìng)爭(zhēng),子葉殘留的少量營(yíng)養(yǎng)在滿足幼苗發(fā)育早期的莖稈伸長(zhǎng)后,難以維持新梢抽生所需的營(yíng)養(yǎng)。因此,對(duì)子葉去除處理幼苗來(lái)說(shuō),實(shí)驗(yàn)結(jié)束時(shí)莖稈仍是種子萌發(fā)期間上胚軸伸長(zhǎng)的產(chǎn)物,比枝長(zhǎng)要小于對(duì)照幼苗。小種子幼苗比枝長(zhǎng)在輕度子葉去除處理最大可能與營(yíng)養(yǎng)較少的小種子在萌發(fā)過(guò)程中對(duì)子葉損傷刺激的應(yīng)激反應(yīng)有關(guān),即幼苗發(fā)育早期在莖生長(zhǎng)投資過(guò)大而后期缺乏持續(xù)的營(yíng)養(yǎng)供應(yīng)導(dǎo)致比枝長(zhǎng)增大。
參考文獻(xiàn):
CHENG JM,ZHAO LP,CHENG J,2009. Seed quality and forest regeneration of a 60-year Quercus liaotungensis forest in the Ziwuling Region,Northwestern China[J]. J Beijing For Univ,31(2):10-16.[程積民,趙凌平,程杰,2009. 子午嶺60年遼東櫟林種子質(zhì)量與森林更新[J]. 北京林業(yè)大學(xué)學(xué)報(bào),31(2):10-16.]
FINCH-SAVAGE WE,CLAY HA,1994. Water relations of germination in the recalcitrant seeds of Quercus robur L.[J]. Seed Sci Res,4(3):315-322.
FUKUMOTO H,KAJIMRUA H,2000. Effects of insect predation on hypocotyls survival and germination success of mature Quercus variabilis acorns[J]. J For Res,5(1):31-34.
GAO XM,DU XJ,WANG ZL,2003. Comparison of seedling recruitment and establishment of Quercus wutaishanica in two habitats in Dongling Mountainous area,Beijing[J]. Chin J Plant Ecol,27(3):404-411.[高賢明,杜曉軍,王中磊,2003. 北京東靈山區(qū)兩種生境條件下遼東櫟幼苗補(bǔ)充與建立的比較[J]. 植物生態(tài)學(xué)報(bào),27(3):404-411.]
GIERTYCH MJ,SUSZKA J,2011. Consequences of cutting off distal ends of cotyledons of Quercus robur acorns before sowing[J]. Ann For Sci,68(2):433-442.
HE YL,WANG MT,DU GZ,2007. Seed size effect on seedling growth under different light conditions in the clonal herb Ligularia virgaurea in Qinghai Tibet Plateau[J]. Acta Ecol Sin,27(8):3091-3098.[何彥龍,王滿堂,杜國(guó)禎,2007. 不同光照處理下青藏高原克隆植物黃帚橐吾(Ligularia virgaurea)種子大小對(duì)其幼苗生長(zhǎng)的影響[J]. 生態(tài)學(xué)報(bào),27(8):3091-3098.]
HOU XG,YI XF,YANG YQ,et al.,2010. Acorn germination and seedling survival of Quercus variabilis:effects of cotyledon excision[J]. Ann For Sci,67(7):711-718.
HUANG SQ,LIU GX,HAN JG,2007. Effects of seed mass and sowing depth on seedling establishment[J]. Partac Sci,24(6):44-49.[黃雙全,劉桂霞,韓建國(guó),2007. 種子大小和播種深度對(duì)種苗建植的影響[J]. 草業(yè)科學(xué),24(6):44-49.]
JIANG XX,LI GL,SHI WH,et al.,2018. Effects of cotyledon loss intensity and time on seedling growth status and reserves translocation in Quercus variabilis [J]. Sci Silv Sin,54(8):56-64.[姜曉旭,李國(guó)雷,史文輝,等,2018. 子葉丟失強(qiáng)度和時(shí)間對(duì)栓皮櫟幼苗生長(zhǎng)狀況和營(yíng)養(yǎng)物質(zhì)轉(zhuǎn)移的影響[J]. 林業(yè)科學(xué),54(8):56-64.]
JRGENSEN MS,LABOURIAU R,OLESEN B,2019. Seed size and burial depth influence Zostera marina L. (eelgrass)seed survival,seedling emergence and initial seedling biomass development[J]. PLoS ONE:e0215157.
LEBRIJA-TREJOS E,REICH PB,HERNNDEZ A,et al.,2016. Species with greater seed mass are more tolerant of conspecific neighbours:A key driver of early survival and future abundances in a tropical forest[J]. Ecol Lett,19(9): 1071-1080.
LI RP,JIANG DM,LIU ZM,et al.,2004. Effects of sand-burying on seed germination and seedling emergence of six psammophytes species[J]. Chin J Appl Ecol,15(10):1865-1868.[李榮平,蔣德明,劉志民,等,2004. 沙埋對(duì)六種沙生植物種子萌發(fā)和幼苗出土的影響[J]. 應(yīng)用生態(tài)學(xué)報(bào),15(10):1865-1868. ]
LIU Y,LI QM,LIU Y,et al.,2013. Effects of mechanical treatments on germination and growth of Quercus fanshanensis and Q. robur seeds[J]. Sci Silv Sin,49(4):39-45.[劉艷,李慶梅,劉勇,等,2013. 機(jī)械處理對(duì)房山櫟和夏櫟種子萌發(fā)和生長(zhǎng)的影響[J]. 林業(yè)科學(xué),49(4):39-45.]
LIU Y,ZHAO Y,CUI D,et al.,2017. Effect of cotyledon damage on the early growth of Sophora alopecuroides seedlings[J]. Acta Pratacult Sin,26(8):139-145.[劉影,趙玉,崔東,等,2017. 子葉損傷對(duì)苦豆子幼苗早期生長(zhǎng)的影響[J]. 草業(yè)學(xué)報(bào),26(8):139-145.]
LOMBARDO JA,MCCARTHY BC,2009. Seed germination and seedling vigor of weevil-damaged acorns of red oak[J]. Can J For Res-Revue Canadienne De Recherche Forestiere,39(8):1600-1605.
MANCILLA-LEYTN JM,CAMBROLL J,F(xiàn)IGUEROA ME,et al.,2013. Growth and survival of cork oak (Quercus suber)seedlings after simulated partial cotyledon consumption under different soil nutrient contents[J]. Plant Soil,370(1):381-392.
MANCILLA-LEYTN JM,CAMBROLL J,MARTN VICENTE ,2012. The impact of the common rabbit on cork oak regeneration in SW Spain[J]. Plant Ecol,213:1503-1510.
PEREA R,MIGUEL AS,GIL L,2011. Leftovers in seed dispersal:ecological implications of partial seed consumption for oak regeneration[J]. J Ecol,99(1):194-201.
PREWEIN C,ENDEMANN M,REINOHL V,et al.,2006. Physiological and morphological characteristics during development of pedunculate oak (Quercus robur L.)zygotic embryos[J]. Trees,20(1):53-60.
PYARE S,LONGLAND WS,2000. Seedling-aided cache detection by heteromyid rodents[J]. Oecologia,122(1):66-71.
SHI WH,BLOOMBERG M,LI GL,2017. Combined effects of cotyledon excision and nursery fertilization on root growth,nutrient status and outplanting performance of Quercus variabilis container seedlings[J]. PLoS ONE,12(5):e0177002.
SHI WH,LI GL,SU SC,et al.,2018. Combined effects of cotyledon excision and nursery fertilization on field performance of Quercus variabilis container seedlings[J]. Sci Silv Sin,54(1):64-73.[史文輝,李國(guó)雷,蘇淑釵,等,2018. 子葉切除與苗圃施肥對(duì)栓皮櫟容器苗造林效果的影響[J]. 林業(yè)科學(xué),54(1):64-73.]
SHI WH,VILLAR-SALVADOR P,JACOBS DF,et al.,2018. Simulated predation of Quercus variabilis acorns impairs nutrient remobilization and seedling performance irrespective of soil fertility[J]. Plant Soil,423:295-306.
SONE K,OISHI K,SHIBATO T,et al.,2016. Effects of acorn attack by curculio weevils on the germination and early growth of Pasania edulis (Makino)seedlings[J]. J For Res,21(4):185-192.
SUN SC,CHEN LZ,2001. The effects of animal removal and groundcover on the fate of seeds of Quercus liaotungensis [J]. Acta Ecol Sin,21(1):80-85.[孫書(shū)存,陳靈芝,2001. 動(dòng)物搬運(yùn)與地表覆蓋物對(duì)遼東櫟種子命運(yùn)的影響[J]. 生態(tài)學(xué)報(bào),21(1):80-85.]
WANG W,MA KP,LIU CR,2000. Seed shadow of Quercus liaotungensis in a broad-leaved forest in Dongling Mountain[J]. Acta Bot Sin,42(2):195-202.
WESTOBY M,JURADO E,LEISHMAN M,1992. Comparative evolutionary ecology of seed size[J]. Trends Ecol Evol,7(11): 368-372.
WU GL,CHEN M,ZHOU XH,et al.,2006. Response of morphological plasticity of three herbaceous seedlings to light and nutrition in the Qinghai-Tibetan Plateau[J]. Asian J Plant Sci,5(4):635-642.
WU GL,DU GZ,2008. Relationship between seed size and seedling growth strategy of herbaceous plant:A review[J]. Chin J Appl Ecol,19(1):191-197.[武高林,杜國(guó)禎,2008. 植物種子大小與幼苗生長(zhǎng)策略研究進(jìn)展[J]. 應(yīng)用生態(tài)學(xué)報(bào),19(1):191-197.]
WU GL,DU GZ,SHANG ZH,2006. Contribution of seed size and its fate to vegetation renewal:A review[J]. Chin J Appl Ecol,17(10):1969-1972.[武高林,杜國(guó)禎,尚占環(huán),2006. 種子大小及其命運(yùn)對(duì)植被更新貢獻(xiàn)研究進(jìn)展[J]. 應(yīng)用生態(tài)學(xué)報(bào),17(10):1969-1972.]
XU ZQ,HUANG XR,XU CL,et al.,2009. The impacts of light conditions on the growth and morphology of Quercus Mongolica seedlings[J]. Acta Ecol Sin,29(3):1121-1128.[許中旗,黃選瑞,徐成立,等,2009. 光照條件對(duì)蒙古櫟幼苗生長(zhǎng)及形態(tài)特征的影響[J]. 生態(tài)學(xué)報(bào),29(3): 1121-1128.]
YAN XF,DU Q,SHI C,et al.,2011. Seedling regeneration of Quercus liaotungensis in Liupan Mountains,China[J]. Chin J Plant Ecol,35(9):914-925.[閆興富,杜茜,石淳,等,2011. 六盤(pán)山區(qū)遼東櫟的實(shí)生苗更新及其影響因子[J]. 植物生態(tài)學(xué)報(bào),35(9):914-925.]
YAN XF,F(xiàn)ANG S,SHI C,et al.,2014. Effects of simulated cotyledon predation on the seed germination and early seedling growth of Quercus wutaishanica [J]. Chin J Ecol,33(4):973-981.[閆興富,方蘇,石淳,等,2014. 模擬動(dòng)物取食子葉對(duì)遼東櫟種子萌發(fā)和幼苗早期生長(zhǎng)的影響[J]. 生態(tài)學(xué)雜志,33(4):973-981.]
YAN XF,ZHOU LB,ZHANG KW,et al.,2012,Cotyledon loss and its effects on survival and growth of Quercus wutaishanica seedlings under different densities[J]. Chin J Plant Ecol,36(8):831-840.[閆興富,周立彪,張靠穩(wěn),等,2012. 不同密度下遼東櫟幼苗子葉丟失及其對(duì)幼苗存活和生長(zhǎng)的影響[J]. 植物生態(tài)學(xué)報(bào),36(8):831-840.]
YI XF,BARTLOW AW,CURTIS R,et al.,2019. Responses of seedling growth and survival to post-germination cotyledon removal:An investigation among seven oak species[J]. J Ecol,107(4):1817-1827.
YI XF,YANG YQ,2010. Large acorns benefit seedling recruitment by satiating weevil larvae in Quercus aliena [J]. Plant Ecol,209(2):291-300.
YI XF,WANG ZY,LIU CQ,et al.,2015. Acorn cotyledons are larger than their seedlings need:Evidence from artificial cutting experiments[J]. Sci Rep-UK,5:8112.
ZHU JL,LIU ZM,2012. Major terminologies and concepts in seed dispersal biology[J]. Chin J Ecol,31(7):2397-2403.[朱金雷,劉志民,2012. 種子傳播生物學(xué)主要術(shù)語(yǔ)和概念[J]. 生態(tài)學(xué)雜志,31(7):2397-2403.]
(責(zé)任編輯 李 莉)
