藜麥CqKEA基因家族的鑒定及表達
2021-10-14 10:24:08張東亮吳筱林田曉芹郭善利陳世華
煙臺大學學報(自然科學與工程版) 2021年4期
關鍵詞:分析
張東亮,吳筱林,田曉芹,褚 晶,郭善利,陳世華
(煙臺大學生命科學學院,山東 煙臺 264005)
KEA(K+/H+逆向轉運蛋白,K+efflux antiporter)在植物細胞鉀離子的積累和平衡過程中發揮重要作用。KEA基因屬于陽離子/質子逆向轉運體(Cation/proton antiporter,CPA) 的CPA2亞家族。目前對KEA的研究較少,已有研究主要集中在模式植物擬南芥。擬南芥中有6個AtKEA基因,與細菌K+/H+反向轉運蛋白具有同源性,由此推測其在K+的積累和平衡過程中發揮功能[1-2]。現有研究結果表明,AtKEA在花萼、保衛細胞和維管組織等不同部位中均存在[3]。AtKEA1—AtKEA3對光合作用、pH和葉綠體滲透具有調節作用,同時會對多種脅迫產生響應[2,4-7]。AtKEA4通過調節內質網的pH來調控蛋白運輸等過程[8]。
藜麥(ChenopodiumquinoaWilld., Cq)是一種起源于南美安第斯山脈的一年生作物,營養價值極高,被聯合國糧農組織(FAO)認定為唯一一種單體作物即可滿足人類所需全部營養的糧食[9],且藜麥對諸如鹽堿、干旱等非生物脅迫具有較強的耐受性[10]。2017年,藜麥高質量參考基因組公布,促進了藜麥基因組的研究[11]。本研究運用生物信息學的方法,對藜麥CqKEA基因家族成員進行鑒定,對其理化性質、基因結構、染色體定位和系統發育關系進行分析,并根據RNA-seq數據對CqKEA基因組織特異性表達及非生物脅迫條件下的表達模式進行探究,為進一步研究藜麥抗逆的分子機理與藜麥遺傳改良提供理論基礎。
1 材料與方法
1.1 CqKEA基因家族成員鑒定與理化性質分析
藜麥基因組數據來源于Chenopodium DB (https://www.cbrc.kaust.edu.sa/chenopodiumdb/download/dow-nload-auth.html)。使用CPA蛋白保守結構域的HMM模型(PF00999)對藜麥所有蛋白質序列進行搜索,Evalue限定為1,獲得候選蛋白;使用NCBI的CDD工具(https://www.ncbi.nlm.nih.gov/cdd/)對候選蛋白的結構域進行鑒定,去除沒有保守結構域的蛋白,從而得到所有CqCPA基因家族成員。從TAIR(https://www.arabidopsis.org/)和NCBI(https://www.ncbi.nlm.nih.gov/)下載擬南芥和水稻的KEA蛋白序列,將它們與CqCPA導入MEGA7.0軟件,采用鄰接法構建系統發育進化樹,從而將CqKEA從中分出。根據其與擬南芥AtKEA的同源關系進行重命名。使用ExPASy的Protparam工具(https://web.expasy.org/protparam/)對CqKEA蛋白序列進行理化性質分析。
1.2 CqKEA基因系統進化與跨膜結構分析
根據藜麥、擬南芥和水稻KEA系統發育進化樹,對其系統進化關系進行分析。同時將鑒定出的CqKEA基因家族成員蛋白序列提交到TMHMM Server v. 2.0(http://www.cbs.dtu.dk/services/TMHMM/#openne-wwindow)進行跨膜結構預測。
1.3 CqKEA基因結構與蛋白motif分析
從參考基因組的基因注釋文件中提取所有CqKEA基因家族成員的基因結構信息,提交到GSDS 2.0在線網站(http://gsds.gao-lab.org/index.php)進行作圖分析;使用MEME 5.1在線網站(http://meme-suite.org/tools/meme)對CqKEA蛋白序列進行motif搜索,可被搜索的motif數量設置為10。
1.4 CqKEA基因表達模式分析
從NCBI下載藜麥生長發育過程中各組織器官及非生物脅迫條件下地上和地下組織的RNA-Seq數據(SRP226463, SRP116149, SRS1538629),使用Kallisto計算表達水平[12]。使用TBtools[13]軟件繪制CqKEA基因家族的組織表達熱圖,使用Origin軟件繪制非生物脅迫條件下地上和地下組織的表達量柱狀圖,分析CqKEA基因的表達模式。
2 結果與分析
2.1 CqKEA基因家族成員鑒定與理化性質分析
本研究共鑒定到10個CqKEA基因家族成員,根據其與AtKEA基因的同源關系將其命名為CqKEA1—CqKEA5c,它們的理化性質如表1所示。蛋白長度從486(CqKEA4a)到1227(CqKEA1)個氨基酸殘基不等,等電點從5.01(CqKEA3b)到7.08(CqKEA4a,4b)不等,大多數蛋白偏酸性。
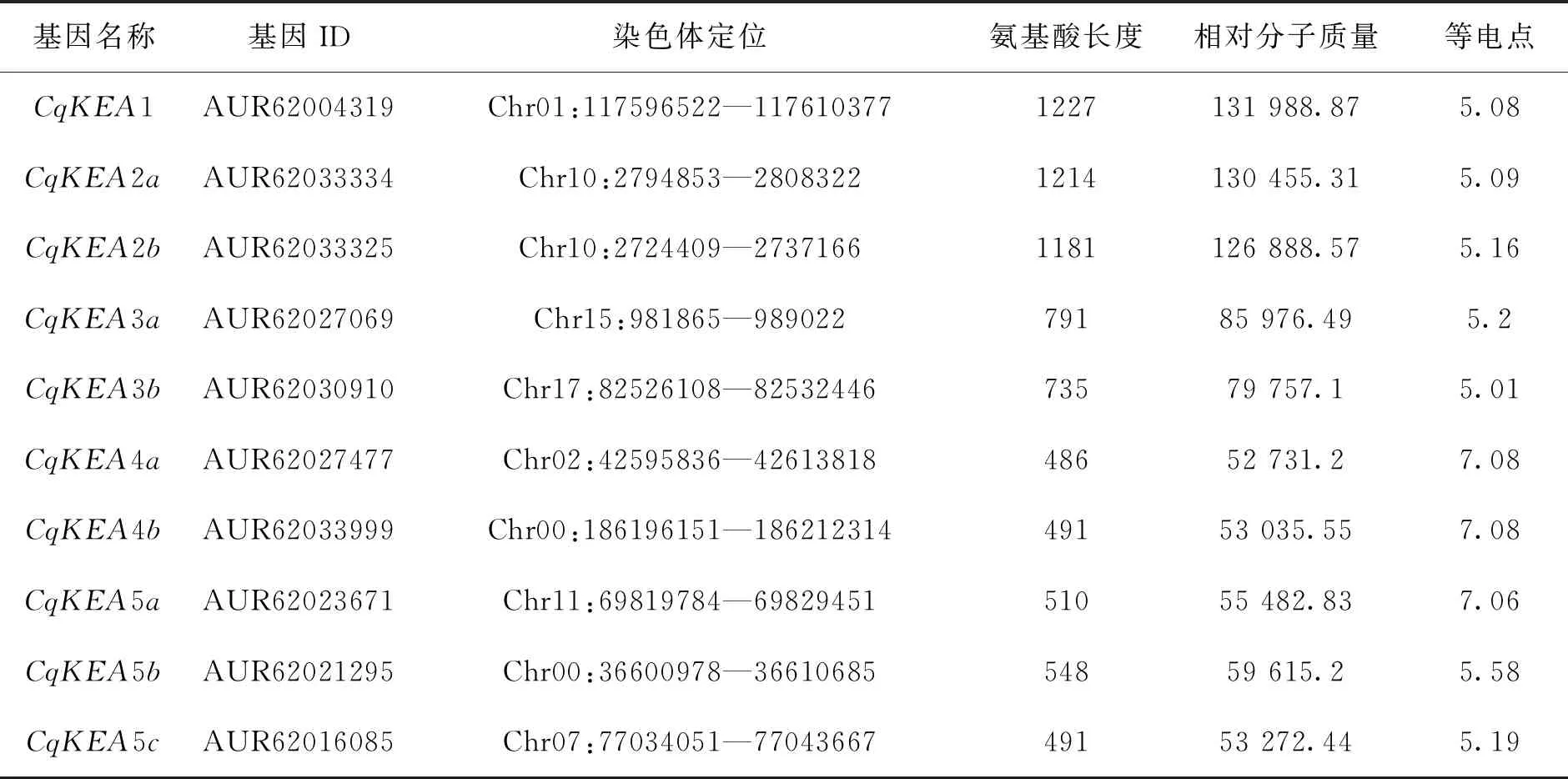
表1 CqKEA基因家族成員理化性質
2.2 CqKEA基因系統進化與跨膜結構分析
以藜麥、擬南芥與水稻的KEA蛋白序列為基礎,使用MEGA7.0構建NJ系統進化樹(圖1)。結果表明,來自上述3個物種的KEA蛋白被分為三個亞組(Class Ⅰ,Class Ⅱ和Class Ⅲ)。Class Ⅰ中包括5個CqKEA蛋白,3個AtKEA蛋白和2個OsKEA蛋白;Class Ⅱ中包括2個CqKEA蛋白,1個AtKEA蛋白和1個OsKEA蛋白;Class Ⅲ中包括3個CqKEA蛋白,2個AtKEA蛋白和1個OsKEA蛋白。

圖1 CqKEA蛋白系統發育進化樹
使用TMHMM對10個CqKEA進行跨膜結構分析,結果如表2所示。CqKEA2a、CqKEA2b和CqKEA4b含有12個跨膜結構域,CqKEA4a和CqKEA5b含有11個跨膜結構域,CqKEA1、CqKEA3a、CqKEA3b和CqKEA5a含有10個跨膜結構域,而CqKEA5c僅有9個跨膜結構域。
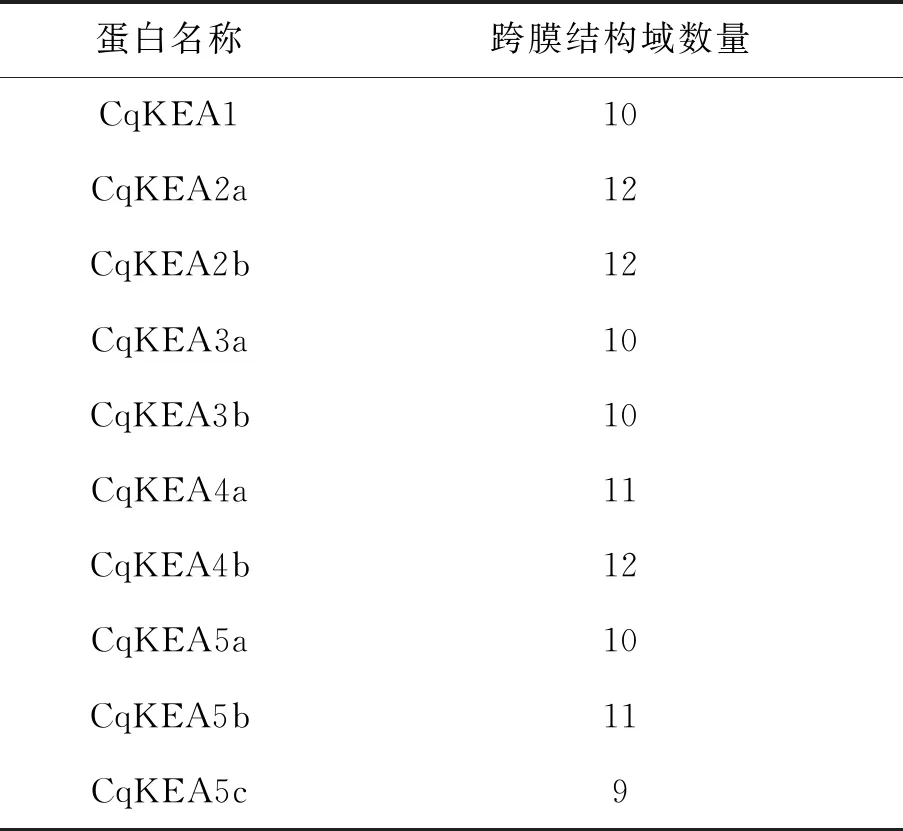
表2 CqKEA蛋白跨膜結構數量
2.3 CqKEA基因結構與蛋白motif分析
使用GSDS對CqKEA基因家族成員的基因結構進行作圖分析(圖2),結果表明,所有CqKEA基因均包含外顯子和內含子,同一亞組基因的結構相似,不同亞組基因結構存在較大差異。外顯子數目在16~21之間,內含子數目在15~20之間。CqKEA5c有16個外顯子和15個內含子,外顯子和內含子的數目最少,而CqKEA2b有21個外顯子和20個內含子,外顯子和內含子的數目最多。
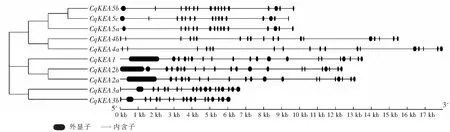
圖2 CqKEA基因結構
使用MEME對CqKEA基因家族成員蛋白序列進行motif分析(圖3),結果表明,在同一亞組的蛋白質所含有的motif種類和數量基本一致,表明CqKEA在進化過程中變化不大,具有較為穩定的功能。Class Ⅲ中3個CqKEA均含有9個motif;Class Ⅱ中CqKEA3a含有7個motif,CqKEA3b含有6個motif;Class Ⅰ中CqKEA4a和CqKEA4b均含有6個motif,而CqKEA5a、CqKEA5b和CqKEA5c均含有8個motif。
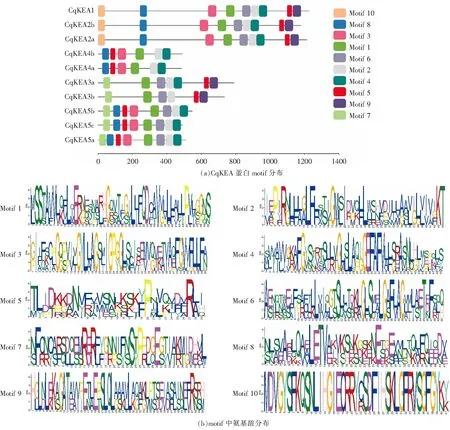
圖3 CqKEA蛋白motif分布
2.4 CqKEA基因表達模式分析
由圖4(a)的組織特異性表達分析結果可知,除CqKEA5a在葉柄中不表達,其余基因在所有組織中均有表達。同一亞組的基因在13個組織中的表達趨勢大致相同,Class Ⅰ中的基因在所有組織中的表達都相對較低,而Class Ⅱ和Class Ⅲ中的基因表達則較高,尤其是CqKEA3a和CqKEA3b在幼苗中的表達最高;CqKEA2a在白甜藜種子和黃苦藜種子中的表達存在差異,這說明在同一物種不同品種之間CqKEA基因的表達模式也存在一定差異。
干旱、高溫和鹽脅迫下表達分析結果表明所有CqKEA基因都會對脅迫產生響應。由圖4(b)可見,在地上部分中CqKEA1、CqKEA2a和CqKEA2b在所有脅迫下均表現上調的趨勢,而CqKEA3a和CqKEA3b在鹽脅迫下上調最明顯,CqKEA4a、CqKEA4b、CqKEA5a、CqKEA5b和CqKEA5c表達量較低,受三種脅迫影響不明顯。由圖4(c)可見,在地下部分中,大多數基因在脅迫條件下呈下調趨勢,只有CqKEA1和CqKEA2a在干旱和高溫脅迫下呈上調趨勢,CqKEA5c在高溫脅迫下呈上調趨勢。
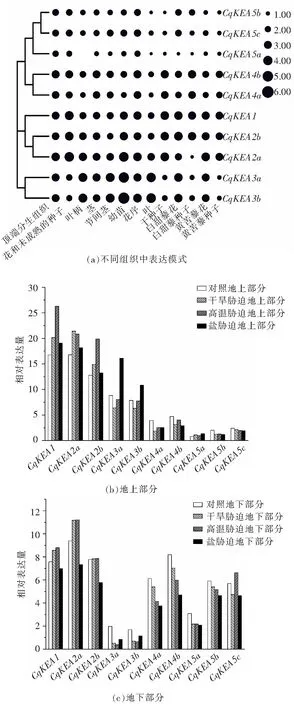
圖4 CqKEA基因表達模式
3 討 論
隨著全基因組測序技術的不斷發展,越來越多的物種的基因組被測序完成,目前已被測序的植物已超過70個[14]。大量的基因組數據為植物基因家族的鑒定和基因功能的驗證提供了極為有利的條件。目前對KEA的研究較少,主要集中在擬南芥。AtKEA1和AtKEA2存在于葉綠體內壁上,AtKEA3存在于葉綠體類囊體膜上,它們通過維持pH和K+平衡來保證葉綠體正常的生物功能[7]。通過系統進化分析表明CqKEA1、CqKEA2a和CqKEA2b與AtKEA1、AtKEA2歸為同一亞組,CqKEA3a和CqKEA3b與AtKEA3歸為同一亞組,由此推測它們具有相似的功能,對光合作用、pH和葉綠體滲透具有調節作用[2,4-7];CqKEA4a和CqKEA4b與AtKEA4歸為同一亞組,其可能參與蛋白運輸、定位和膜微囊融合等過程[8]。
植物通過多種方式來抵抗逆境脅迫帶來的影響,但是在耐受干旱、鹽和高溫等脅迫的過程中,植物的基因作用機制表現出相似性[15]。本研究對10個CqKEA基因的組織特異性表達和非生物脅迫條件下的表達模式進行了探究。絕大多數基因在所有組織中均有表達,說明CqKEA基因可能是維持植物基本生命活動所必需的。CqKEA3a和CqKEA3b在幼苗中的表達量最高,推測其可能參與維持植株正常生命活動中的離子穩態和K+利用。在地上部分中CqKEA4a、CqKEA4b、CqKEA5a、CqKEA5b和CqKEA5c的表達水平相對較低,而在地下部分中表達水平相對較高,推測這5個基因主要在地下部分發揮功能;CqKEA3a和CqKEA3b則主要在地上部分中發揮功能,同時當植株受到鹽脅迫后這兩個基因表達水平上調,推測其可以通過維持植株正常的離子穩態來保證植株正常的生命活動。在CqKEA1、CqKEA2a和CqKEA2b中,除地下部分CqKEA2b對干旱和高溫脅迫不敏感,其余基因均對干旱和高溫脅迫有響應,推測其可能在這兩種脅迫下行使功能。綜上所述,本研究對CqKEA基因家族進行了系統分析,為CqKEA基因功能研究奠定了理論基礎,為研究藜麥耐逆機理和分子育種提供了基因資源。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
財經界(學術版)(2015年20期)2015-12-23 09:20:13
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
