絨毛白蠟耐鹽堿響應機制的研究進展
2021-10-13 09:23:56范馨月趙繼業杜子沫叢日晨
中國農學通報 2021年28期
范馨月,張 華,趙繼業,杜子沫,叢日晨
(1北京市園林科學研究院,北京100102;2河北農業大學,河北保定071000)
0 引言
植物在遭受非生物脅迫時可以感應脅迫信號并且觸發相應響應機制,通過改變自身的生長、代謝和發育過程來應對脅迫,以避免其受到傷害[1-2]。植物應對各種不同脅迫有不同的響應機制,但也會存在部分交叉。
土壤鹽堿化是一個日益嚴重的全球性問題,造成土壤鹽堿化的原因除了自然發生的土壤鹽堿化,氣候變化和不良的灌溉方式也被認為是主要因素[3]。糧農組織最近的一項報告顯示,目前全球20%以上的灌區受到土壤鹽堿化的影響,到2050年,這一數字將可能擴大到50%[4]。中國是鹽堿地大國,2015年國家林業局調查國內鹽堿土地面積為17.79萬km2[5],約占全世界的10%。
絨毛白蠟(Fraxinus velutina)又叫絨毛梣,屬木犀科白蠟屬,與其他白蠟對比,絨毛白蠟具有較強的耐鹽堿能力[6],在天津、秦皇島等濱海地區表現良好,是優良的鹽堿地造林樹種,適用于濱海城市綠化。目前,關于絨毛白蠟的耐鹽性的研究主要集中在耐鹽品種的選育和培育以及生理特性上,對于絨毛白蠟在鹽脅迫下的響應機制研究較少。耐鹽堿基因的表達、滲透調節、離子的選擇吸收等是絨毛白蠟耐鹽堿的關鍵,探討絨毛白蠟耐鹽堿的生理和分子響應機制有利于通過分子手段改良品種,為喬木耐鹽堿研究提供部分依據,豐富鹽堿地綠化喬木樹種。
1 鹽堿脅迫對絨毛白蠟的影響
鹽漬環境對植物的影響幾乎囊括了從種子萌發到植株生長最后死亡的整個生命過程,鹽脅迫會造成生理干旱、減弱葉片的光合作用、損壞細胞膜、造成代謝紊亂等,最終可能使植物營養不良甚至死亡[7]。
1.1 對種子萌發影響
種子能在鹽堿地順利萌發是植物具有耐鹽堿能力的重要表現之一。研究表明隨著脅迫的增強,絨毛白蠟發芽率基本上是先升高后降低。低濃度的鹽脅迫可以適當提高絨毛白蠟種子的發芽率,高濃度則會抑制種子的發芽率。劉萍等對鹽脅迫下未萌發的種子進行復水實驗,表明在一定臨界值內沒能萌發的種子在復水實驗后可以恢復萌發,恢復率達90%左右,但是當鹽濃度超過臨界值時,恢復率逐漸下降。說明絨毛白蠟的種子對于在一定臨界值內的鹽脅迫有一定的適應能力,且保持萌發的潛力[8-9]。雖然絨毛白蠟植株具有較強的耐鹽堿性,但鹽脅迫也會明顯影響種子的形成質量,鹽堿地種子千粒重僅為非鹽堿地的78%。
1.2 對絨毛白蠟形態及生長的影響
鹽分對植物整個生命周期都有較為顯著的影響,鹽脅迫會抑制組織和器官的生長,導致生長不良等[10]。在鹽脅迫下,植物芽部和根部的生物量都會顯著降低[11]。對鹽脅迫下絨毛白蠟的各項生長指標的測定發現,在鹽脅迫下,絨毛白蠟的株高、地莖以及葉片面積均隨著脅迫強度的增大和時間的延長呈現下降趨勢,在高濃度鹽處理中,葉片開始出現干枯脫落現象。隨著鹽脅迫濃度的升高,植株整體和各器官的生物量均大幅下降,且降低的速度加快、幅度增大[12]。
1.3 對光合速率和葉綠素含量的影響
光合作用為植物提供生長所需能量。在鹽脅迫下,絨毛白蠟的光合速率隨鹽濃度的增加而逐步降低。主要原因是光合作用需要吸收二氧化碳,而鹽脅迫會造成氣孔導度下降使葉片不能順利吸收二氧化碳。此外,鹽脅迫還會降低葉綠素含量,破壞葉綠體的結構。
葉綠體是植物進行光合作用的重要細胞器,在鹽堿脅迫下容易受到影響。植物進行光合反應時起主要作用的色素是葉綠素。研究表明,鹽脅迫會造成絨毛白蠟體內葉綠素含量的降低,葉綠素含量與脅迫強度呈反比。葉綠素含量下降的主要原因是葉綠素被葉綠素酶分解,同時光合酶受到抑制。另外鹽脅迫還會破壞葉綠體的結構,鹽脅迫下產生的大量自由基破壞了葉綠體質膜,阻礙了類囊體膜的形成,也破壞了類囊體膜的結構,使垛疊狀態的類囊體膜比例下降[13]。
1.4 對細胞膜透性的影響
鹽脅迫下細胞質膜最先受到傷害,鹽脅迫導致膜脂過氧化,積累大量的丙二醛,使得細胞質膜透性增大,細胞膜的相對電導率增大[14]。
1.4.1 相對電導率 鹽脅迫下植物的相對電導率是研究植物抗鹽性的理想指標之一,細胞膜越完整相對電導率就會越低[15]。研究表明,鹽脅迫會破壞白蠟葉細胞中的細胞膜系統,在鹽堿脅迫對白蠟種子的研究中發現,在相同鹽脅迫下,相比采于保定的白蠟種子,秦皇島的白蠟種子具有更低的相對電導率,這說明采于秦皇島種子的抗鹽性更強,當地的濱海氣候和土壤環境可能是誘因[16]。
1.4.2 MDA(丙二醛)植物體內丙二醛的含量是衡量細胞膜膜質過氧化程度的一個指標,也可以用來判斷植物抗性的強弱。隨著鹽濃度和時間的增加,絨毛白蠟種子和葉片中的MDA含量增加,但葉片中MDA含量的增長速度不同,在絨毛白蠟受到鹽脅迫的初始階段,MDA的含量增長幅度較小且增速較慢,這可能是由于絨毛白蠟在鹽脅迫初期會通過自身調控來降低鹽脅迫對細胞的傷害,保護自身膜質不被破壞。但隨著脅迫的增強,葉片中的MDA含量會呈現較大幅度的上升且增長較快,說明長時間的高鹽環境會使自身調節機制受損,加快了細胞膜的過氧化進程[16-17]。
2 絨毛白蠟對鹽脅迫的響應機制
2.1 絨毛白蠟響應鹽脅迫的生理機制
2.1.1 形態結構對鹽脅迫的適應性變化 在長期的生物進化過程中,幾乎每種植物都形成了適應自身生長環境的形態結構,在鹽堿環境下各類植物的結構也會為適應環境而產生一系列的變化。
絨毛白蠟葉片表面存在可分泌鹽分的腺體,可以將鹽分排出體外。在鹽脅迫下絨毛白蠟的葉面積隨著鹽濃度的升高而降低,出現干物質大量消耗的情況,表現為生物量減小[18],而且在鹽脅迫下的絨毛白蠟葉片有肉質化趨勢。鹽脅迫不僅會影響植物莖葉等外部結構,也會影響植物的內部結構。在重度鹽堿地的白蠟葉片的柵欄組織和海綿組織厚度的比值在所有樹種中最大,海綿組織排列較為疏松,細胞間空隙會形成發達的通氣組織,表現出對鹽漬環境較強的適應性[19]。
根是從土壤中吸取營養的重要器官,根系的解剖結構與植物耐鹽性有直接關系。研究發現,絨毛白蠟具有根系直徑較大、側根發達、延伸較長等特點,并且絨毛白蠟的根系有一定的避鹽性[20],且相比于正常生長條件,鹽堿地的絨毛白蠟根莖比會增大,這可能是絨毛白蠟可以在鹽堿環境下生存的原因之一。
2.1.2 離子轉運與區域化 鹽脅迫會引起離子毒害,對有害離子轉運和區域化的限制可以有效減少危害。引起離子毒害的主要離子是Na+,相關研究已經發現了幾種Na+轉運方式[21-22],如圖1所示。
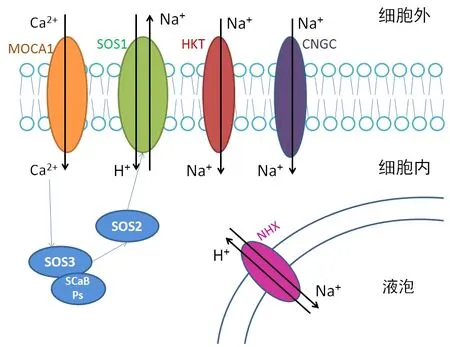
圖1 植物細胞內Na+的4種轉運方式
植物體內的Na+、Cl-的分布可以在一定程度上反映植物對于鹽脅迫的耐受能力,在絨毛白蠟中Na+的分布大致為細根>主根>莖>葉,細根是直接從外界吸收水分以及營養物質的部位,是最先接觸到Na+和Cl-的部位,因此是積累離子最多的部位。鹽分主要積累在絨毛白蠟根的皮層外圍細胞內,而且絨毛白蠟的葉肉細胞內有大液泡,也可以將離子區域化,這可能是絨毛白蠟具有較強耐鹽能力的原因之一[23-24]。
2.1.3 滲透調節 滲透脅迫會造成植物缺水、植物氣孔關閉、細胞擴張受到抑制、生長速度減緩等[25]。調節水勢的方法主要有2種,一種是積累無機離子,另一種是積累有機滲透物質。細胞內有過多的無機離子的積累,會產生離子毒害,所以植物可以通過積累脯氨酸、可溶性糖等有機物滲透物質來調節滲透平衡[26-27]。
(1)可溶性糖。植物組織中普遍存在可溶性糖,種類較多,有葡糖糖、果糖等。絨毛白蠟體內的可溶性糖含量隨著鹽脅迫強度的增加而增加[12]。武德等[17]研究表明,在對絨毛白蠟進行鹽脅迫的開始階段,體內可溶性蛋白含量變化是先下降后逐漸增加,下降的原因可能是鹽分將可溶性蛋白破壞分解成了氨基酸,之后含量增加可能是植物啟動了自我防護機制,產生了一些可溶性蛋白以抵抗逆境,因此絨毛白蠟對鹽脅迫有一定的抵抗和適應能力。
(2)脯氨酸。脯氨酸也是一種有機滲透物質,對提高植物的耐鹽堿性有重要作用,還能增加植物體內抗氧化酶的活性[28]。閆文華等[29]研究表明,當鹽脅迫的強度逐漸增大時,脯氨酸的含量也在逐步增加,在達到一定強度時,脯氨酸含量達到峰值,之后脯氨酸含量開始逐步減少,這可能是由于鹽分對植物的傷害超出了植物所能承受的范圍,滲透調節物質合成受阻。
2.1.4 保護酶的作用 在氧的代謝過程中會產后一種激發狀態的氧分子被稱為活性氧。活性氧具有很強的氧化能力,可引起細胞膜老化、造成不可逆的傷害甚至是細胞死亡[30],超氧化物歧化酶(SOD)是植物清除活性氧的第一個屏障[31],在保護酶系統中處于中心地位,另外還包括過氧化物酶(POD)、過氧化氫酶(CAT)等保護酶。這些抗氧化酶活性可以作為反映植物抵御鹽脅迫能力的關鍵指標。
研究表明,鹽脅迫強度加大會引起絨毛白蠟體內的SOD含量呈先升高后下降的趨勢,先上升的原因是,植物在遭受不良環境時為了使活性氧處于相對平衡的水平,會產生SOD清除植物體內多余的活性氧。但隨著脅迫時間和強度的上升,SOD活性下降,表明SOD對植物的保護作用是在一定范圍內的,超過一定限度植物的自我調節機制被破壞。
保護酶系統除SOD外還有過氧化物酶(POD)和過氧化氫酶(CAT)。研究表明,隨著脅迫時間的延長,POD活性呈現“M”的趨勢,POD可以有效清除植物體內的過氧化氫。推測在鹽脅迫過程中,植株通過提高POD的活性來保護植物免受傷害,以提高絨毛白蠟的抗性。吳麗云等[32]通過對絨毛白蠟種子的研究表明,CAT活性隨著鹽脅迫強度的升高而增加。POD和CAT酶活性與鹽濃度變化的關系還有待進一步研究。
2.2 絨毛白蠟響應鹽脅迫的分子機制
2.2.1 差異表達基因 對于絨毛白蠟的耐鹽堿性,無論國內外的研究大部分停留在生理生化層面,而對更加深入的分子調控機制方面研究較少,缺乏較為全面的研究和分析。目前已經有不同種白蠟在不同鹽漬環境下的差異基因分析以及絨毛白蠟在不同脅迫時間下的基因分析。
通過對在鹽脅迫下和對照組的絨毛白蠟和中國白蠟進行轉錄組測序,對差異基因進行分析,研究中檢測了差異表達基因(DEGs),并分析了相關信號通路,上調基因主要集中在生物堿合成、氧脂質的生物合成過程、嘌呤核苷酸轉運、細胞銅離子穩態、果糖刺激產生植物類金屬硫蛋白、光系統Ⅰ中的光合電子傳遞、類黃酮生物合成、促性腺激素信號通路、淀粉和蔗糖代謝等。下調的基因主要與光合作用、硒復合代謝過程、雙受精形成合子和胚乳的過程以及苯丙類生物合成有關[33-34]。
研究通過對不同鹽處理時間下(0.5、24 h)的絨毛白蠟進行表達譜分析,發現持續0.5 h鹽脅迫的絨毛白蠟與對照組相比,并沒有發現較多的差異表達基因,而當鹽脅迫持續24 h后差異基因大量增多,說明要引起顯著的基因表達量差異需要長時間的鹽脅迫。且持續24 h鹽脅迫與0.5 h相比差異基因中所涉及的功能顯著增多。通過分析絨毛白蠟在不同脅迫時間下的差異基因,篩選出一些用于響應鹽脅迫的關鍵基因,其功能主要有滲透保護物質合成、調節離子平衡、水通道蛋白合成、抗氧化酶的生成、激素信號轉導、光系統保護和修復、轉錄調控等多方面[35]。
“懷舊”,一方面是兩岸聽眾的審美特征,而另一方面也反映出當代華語流行樂壇存在的問題。如當前各個經紀公司在籌劃歌手個人演唱會時,都在打“懷舊”牌,列出該歌手多年來唱過的經典曲目以增加演唱會的市場號召力,讓歌迷到現場“見證青春歲月”。[29]這一方面證明了這些流行歌手的影響力,另一個方面也顯示了整個流行樂壇一定程度上的青黃不接。[29]
差異基因所涉及的基因功能與上述生理相應機制有一定重疊,絨毛白蠟響應鹽脅迫的調控機制很復雜,受多基因控制,涉及多種功能基因以及調控基因,并且與其他脅迫的響應機制存在交叉現象。
2.2.2 關鍵基因的分析
(1)FvNCED3基因。脫落酸(ABA)具有調節種子胚的發育、引起氣孔閉合以及促進休眠和器官衰老等作用,可以增強植株對干旱、鹽堿和高溫等非生物脅迫的抗性。植物組織中內源ABA水平受脫落酸合成、羥基化代謝、可逆糖基化代謝以及ABA長距離轉運等過程的動態調控[36-37]。ABA可以通過參與信號的傳導來調控相關基因的表達,以增強對脅迫的應對能力[38]。
目前關于高等植物中ABA的合成途徑已經較為清晰,如圖2所示[39]。
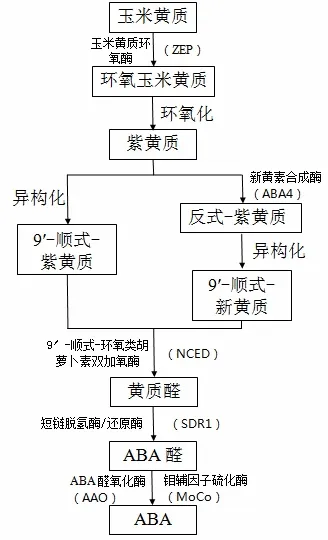
圖2 ABA合成途徑
研究表明,鹽脅迫使植物體內ABA的含量上升,基因NCED可以調控鹽脅迫下ABA的合成[40]。目前已知的體內包含此類基因的植物除了絨毛白蠟外還有海棠、擬南芥、煙草等[41]。相關研究表明,在鹽脅迫條件下NCED的表達水平越高則ABA含量越高,因此想增強植物的抗性可以通過提高NCED基因的表達水平來實現[42],目前關于NCED基因在植物抵抗非生物脅迫方面的研究多數集中在干旱脅迫方面[43]。
研究人員已經分離和克隆出了絨毛白蠟體內的FvNCED3基因及其啟動子,并且進行了基因功能分析。發現FvNCED3在不同組織中的表達存在差異,葉片中的表達水平高于根中。在轉錄水平上,在脫水、鹽脅迫和外源ABA的作用下FvNCED3的表達量升高,在高溫和低溫脅迫下表達量降低,對紫外線無反應,在干旱和鹽脅迫條件下,FvNCED3啟動子誘導基因表達的能力顯著增強,有效提高了報告基因的表達,表明FvNCED3可以通過對外界刺激的響應來增強植物下游基因的表達和抗逆性[44]。
目前ABA激活SnRK2s的機制基本被闡明,如圖3所示。在正常生長條件下,蛋白磷酸酶PP2C會抑制SnRK2s的活性,而在植物遭受滲透脅迫時則會激發其響應機制。研究表明SnRK2主要激活ABA的上游。首先滲透壓力通過一個假定感受器OSCA1來觸發鈣通道,Ca2+可以激活SnRK2s,導致ABA含量增加,ABA與受體PYLs結合可以抑制蛋白磷酸酶PP2C,從而激活第三類SnRK2,使其磷酸化[46-47],同時也會磷酸化下游的轉錄因子AREB/ABFs/NTL,最后使相關脅迫響應基因表達。SnRK2s也會激活其他響應因子來增強抗性。
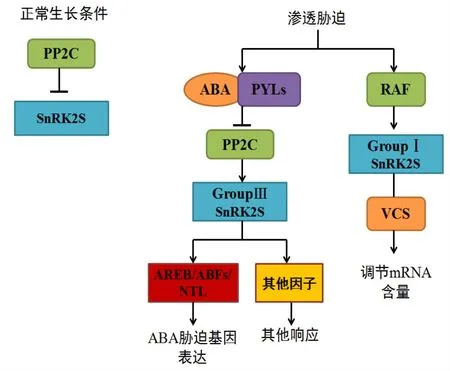
圖3 植物SnRK2s參與的主要調控途徑
研究表明,I類SnRK2s可以通過磷酸化mRNA-decapping激活子VCS,調控mRNA的含量。東京大學分析鑒定出3個RAF激酶作用于I類SnRK2s上游,以此來參與植物在滲透調節下的生長,加深了對非依賴于ABA的滲透脅迫信號調控機制的認識[48]。同時也發現部分RAF激酶會受到ABA和滲透脅迫2個方面的影響[49]。
通過對絨毛白蠟進行轉錄組測序可以知道,在鹽脅迫條件下,FvSnRK2基因的表達量顯著增加,由此可以推測FvSnRK2是使絨毛白蠟具有較強抗鹽性的關鍵基因。目前在絨毛白蠟抗鹽脅迫研究中,發現3個SnRK2家族的相關基因,分別為FvSnRK2182、FvSnRK5271和FvSnRK748,其編碼蛋白均為親水性。通過對前2個基因的克隆和分析可知,FvSnRK2182基因全長為1773 bp,共編碼363個氨基酸,FvSnRK5271基因全長1267 bp,編碼了343個氨基酸[50]。通過將基因在擬南芥中異源表達顯示基因過量表達的植株對鹽脅迫具有更強的抗性,側根數量明顯增多,地上干鮮重、地下干鮮重、葉綠素含量、K+含量明顯上升,而Na+含量明顯降低。
(3)轉錄因子的表達。植物在響應非生物脅迫時,轉錄因子是植物中一類重要的調節因子,與結構基因不同,轉錄因子與逆境調控基因并不是一一對應的。研究人員已經在馬鈴薯、擬南芥等植物中發現了與植物耐逆性相關的MYB轉錄因子。目前已經克隆了絨毛白蠟的FvMYB1、FvMYB2基因,cDNA全長分別為1203和1201 bp。研究結果表明,FvMYB1的表達量具有組織差異性,莖中表達量最高,而FvMYB2沒有組織差異性,在各組織中表達差異不顯著。兩者都在一定程度上受鹽脅迫誘導表達,對植物增強自身抗鹽性有一定作用,主要表現在增強保護酶活性、增加脯氨酸和葉綠素含量等方面[35]。
2.2.3 DNA甲基化 植物對非生物脅迫的響應還包括轉錄前表觀遺傳調控的參與,以此調控基因來編碼相關蛋白質[51]。越來越多的研究表明,表觀遺傳機制在植物應答非生物脅迫時發揮著重要作用[52],通過表觀修飾會導致基因表達以及基因功能發生變化。一般來說甲基化程度高會造成基因表達量減少,相關研究表明將絨毛白蠟和水曲柳進行雜交得到的子代植株具有明顯的雜種優勢,通過比較雜交子代和親本種內開放授粉植株,發現F1雜交種在抗鹽脅迫方面有了明顯提高。研究表明F1雜交種的DNA甲基化程度低于親本種內開放授粉植株的甲基化程度,且隨著鹽脅迫時間的增加,雜交種和種內授粉植株的DNA甲基化程度都增加,表明脅迫會使植物生長過程中DNA甲基化修飾程度提高,鹽脅迫與DNA甲基化有關,并且會改變甲基化的模式[53]。
3 展望
通常所說的鹽堿脅迫是鹽化和堿化混合,人們經常將鹽堿混為一談,其實很多研究表明,鹽脅迫和堿脅迫并不相同,是2種非生物脅迫類型[54]。2種脅迫對植物的傷害不同,鹽脅迫一般會引起離子脅迫和氧化損傷,而堿脅迫除了造成上述傷害外,還會引起高pH脅迫[55]。響應鹽脅迫和堿脅迫的分子機制會有共同點,也會有不同[56]。目前對于堿脅迫對植物的影響已經有了一些研究,但相對鹽脅迫來說要少得多。對于絨毛白蠟在堿性脅迫下的反應,已經有了相關研究,實驗證明堿性鹽脅迫下脯氨酸的變化與中性鹽脅迫不同,并且葉片中POD、SOD、CAT的變化比較復雜,并不成單一的變化趨勢,這與中性鹽脅迫不相同,其原因還有待進一步研究[17]。
植物一般具有交叉適應性。交叉適應性指植物遭受某一種逆境,啟動一種自我調控機制時,同時也會提高對另一種逆境的抵抗能力。目前對于植物應對單一脅迫的研究已取得大量進展,而植物在自然界中遭遇的不良條件通常是幾種脅迫同時存在,因此對植物響應聯合脅迫的機制研究已成為重點方向[57]。因此各脅迫之間以及各調控機制之間會存在交叉重疊。干旱通常會造成滲透脅迫,而鹽分則會造成滲透脅迫和離子毒害2個方面的影響,因此干旱脅迫和鹽脅迫有一些重疊信號。已對一些物種進行過旱鹽脅迫的研究,例如文冠果、沙棗、灰胡楊等。對絨毛白蠟的耐鹽堿性可以集中于同時發生多種脅迫時的調控機制,以及生物脅迫和非生物脅迫信號之間的交互作用上。
目前對植物鹽脅迫的研究實驗大多在培養的無菌苗上進行,然而在自然環境中,植物是與微生物等共同生存的,今后的研究也可集中在了解相關微生物對植物抗性的影響上,有利于更加系統和完整地了解植物響應非生物脅迫的機制。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
文苑(2018年21期)2018-11-09 01:23:06
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
中國衛生(2015年9期)2015-11-10 03:11:12
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
