引種雜交楊光合-光響應特征
2021-10-13 09:23:54古麗娜兒王興勝夏詩書
中國農學通報 2021年28期
陳 勇,古麗娜兒,王興勝,夏詩書,程 平,李 宏
(1伊犁哈薩克自治州林木良種繁育試驗中心,新疆伊犁835311;2新疆林業科學院,烏魯木齊830000)
0 引言
光合作用是植物吸收光能轉化為生物能的重要生命活動,光是進行光合作用的前提條件。光合-光響應曲線反映了光合有效輻射與植物凈光合速率之間的關系[1],對植物進行光合-光響應曲線測定,可以計算得到植物重要的光合生理參數,如最大凈光合速率(Pmax)、光補償點(LCP)、光飽和點(LSP)、表觀量子效率(AQY)、暗呼吸速率(Rd),最大凈光合速率反映植物的最大光合潛力,光補償點反映光合作用過程中呼吸消耗與光同化作用相等時的光照強度,光飽和點反映植物對強光的利用能力,表觀量子效率反映植物在低光環境對弱光的利用轉化效率[2]。不同植物的光響應特征相異,研究植物的光合-光響應特征是分析環境變化對植物生長影響的可靠手段[3]。光合-光響應模型是模擬植物光合作用對光合有效輻射的核心構建,目前用于光合-光響應擬合的模型有直角雙曲線模型[4]、非直角雙曲線模型[5]、指數模型[6]和直角雙曲線修正模型[7-8]。其中非直角雙曲線模型應用最為廣泛[3,9-11],但其模型曲線不存在極點[12],無法擬合出LSP,且擬合出的Pmax大于實測值,諸多學者[13-18]證明直角雙曲線模型能夠克服這些不足,擬合值貼近實測值,擬合效果好。因此,筆者選用直角雙曲線修正模型對伊犁地區22個引種雜交楊品系進行光合-光響應擬合,闡明引種雜交楊光合生理特性,研究結果可用于篩選適宜伊犁地區的高光效引種雜交楊,旨在為該地區優良樹種選育提供理論依據。
1 材料與方法
1.1 研究區概況
試驗地位于新疆伊犁州林木良種繁育試驗中心(43°45′N,81°09′E),平均海拔660 m,地下水位1~1.5 m。該區屬溫帶大陸性氣候,年均氣溫6.7~9.9℃,極端高溫40.0℃(1997年),極端低溫-43.2℃(1969年),1月均溫-12.2℃,7月均溫22.7℃。年降水量417.6 mm,年蒸發量1425 mm,無霜期164天。試驗地土壤類型為沙壤土,pH 8.4,土壤有機質含量14.95 g/kg,水解氮含量78.86 mg/kg,有效磷含量42.96 mg/kg,速效鉀含量150 mg/kg,土壤肥力中等。
1.2 試驗材料
研究對象為2016年從遼寧省楊樹研究所引種的雜交楊,生長良好的22個楊樹品系(表1)。2017年4月,按照完全隨機試驗設計,每個品系扦插100株,行向為南北走向,每個品系之間以當地品種‘加小×俄9#’間隔。田間管理采用常規方式。

表1 參試引種雜交楊品系
1.3 測定方法
2017年8月選擇晴朗天氣進行多天測量(8月16—22日),測定時間為10:00—12:00,使用配備6400-02B Led光源的Li-6400XT便攜式光合測定儀器測定光合-光響應(Pn-PAR)曲線,選用Light-Curve Auto程序進行測定,流量設定為500 μmol/s,通過CO2注入系統使CO2濃度保持在自然狀態(400 μmol/mol),光合有效輻射值(PAR)依次設定為0、20、50、100、150、200、500、800、1100、1300、1500、1700、1900、2100、2300、2500 μmol/(m2·s)共 16 個梯度,即從 0 μmol/(m2·s)到2500 μmol/(m2·s)逐漸遞增的順序測定光合-光響應(Pn-PAR)曲線。
1.4 數據處理及應用模型
使用Photosynthesis Workbench分析軟件進行光合-光響應(Pn-PAR)曲線的擬合分析,使用直角雙曲線修正模型[7]對(Pn-PAR)曲線進行修正,如式(1)。
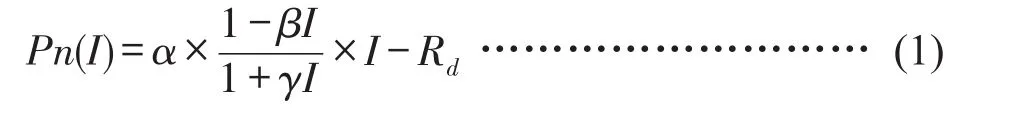
式中,α表示初始量子斜率,I表示光合有效輻射,Rd為暗呼吸速率,β為修正系數,γ為與光合有效輻射無關的系數。
對弱光條件下[I≤200 μmol/(m2·s)]的光響應曲線進行直線回歸,利用所得方程計算表觀量子效率(AQY)、光補償點(LCP)、暗呼吸速率(Rd),回歸方程如式(2)。

使用SPSS 21.0進行單因素方差分析與多重比較,使用Excel 2013制圖。
2 結果與分析
光合-光響應曲線能夠反映植物凈光合速率(Pn)隨外界光環境變化的動態規律。不同引種雜交楊品系光合-光響應曲線擬合決定系數均在0.9900以上,部分品系擬合決定系數達到1.0000,擬合效果好。各參試品系光合-光響應變化趨勢不盡相同(圖1),0~200 μmol/(m2·s)光照條件下,‘93-8-17’、‘93-8-18’、‘108’、‘D5-9’和‘D5-26’的Pn上升速度較快,而‘D5-20’、‘I-262’、‘LX-1’、‘LX-2’、‘TL-13’和‘中綏12#’在該光照條件下Pn上升速度較慢;Pn在光照較弱時會隨著PAR值的升高而急劇上升,在達到光飽和點后Pn值上升態勢減慢甚至下降,在超過負荷強光不能及時有效進行吸收利用時,便會受強光照脅迫,造成光抑制。各參試品系Pn值隨著光照增強急劇上升,但參試品系飽和光強過高,未達到光飽和點,因此參試品系均未出現光抑制現象。
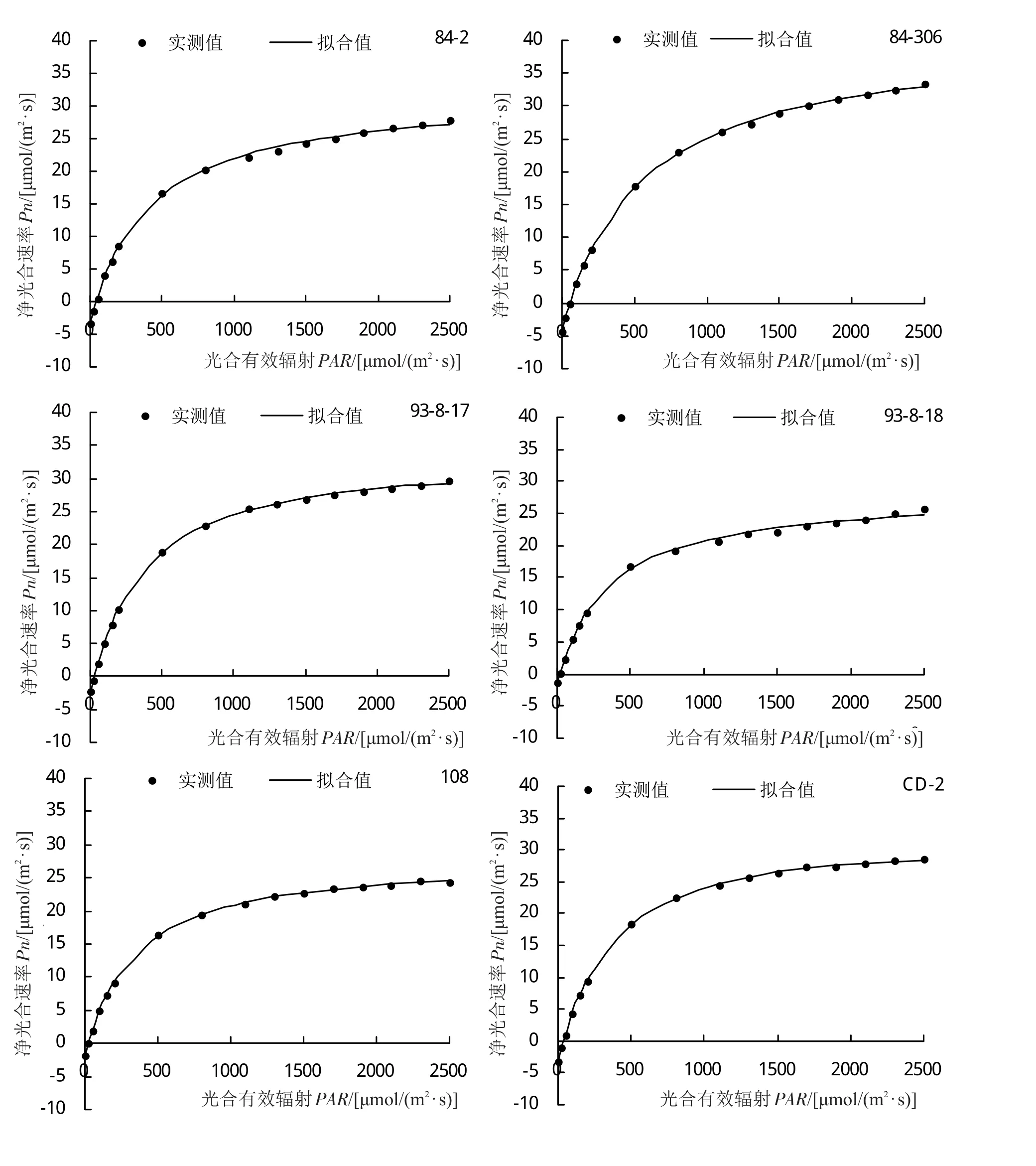
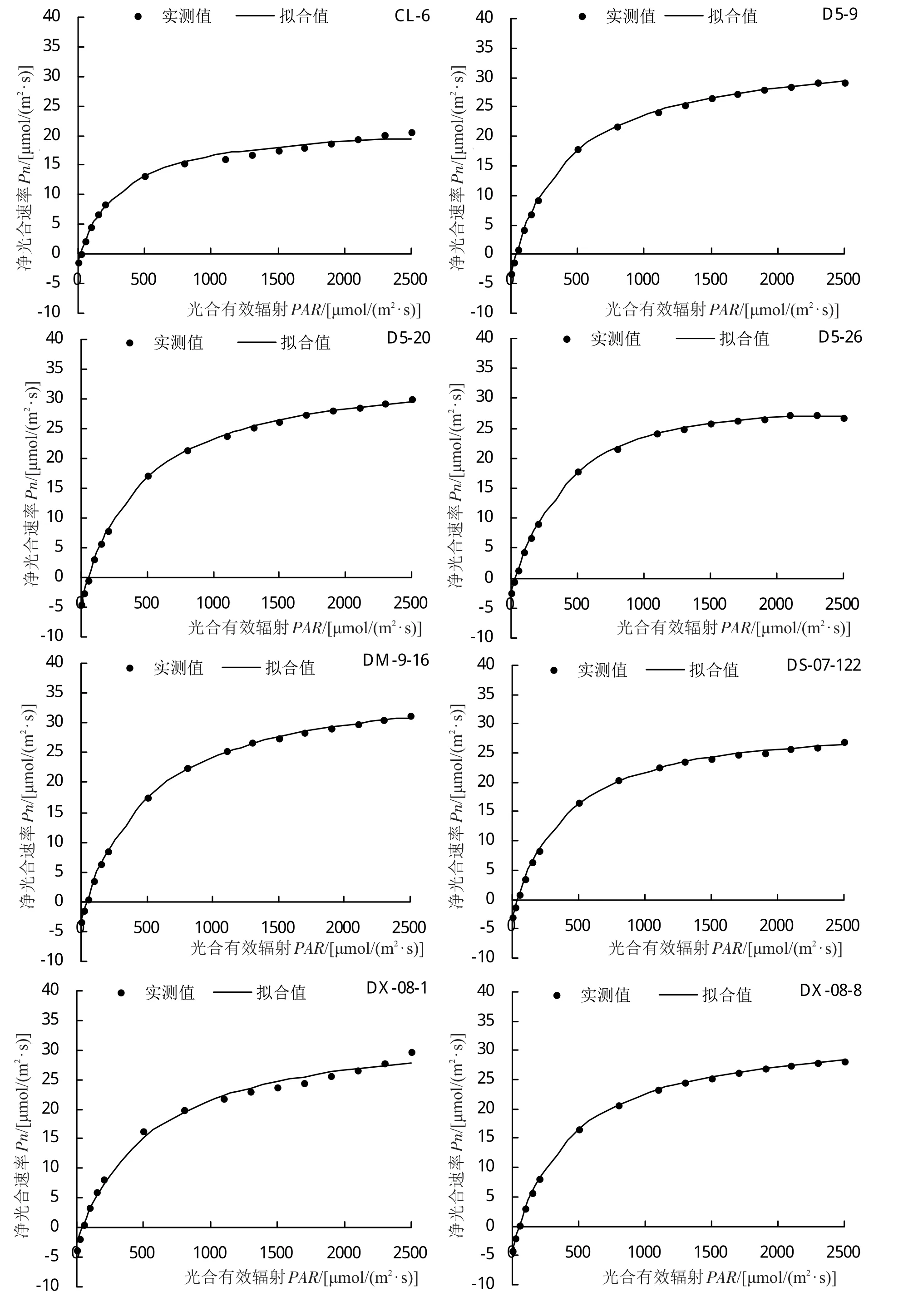
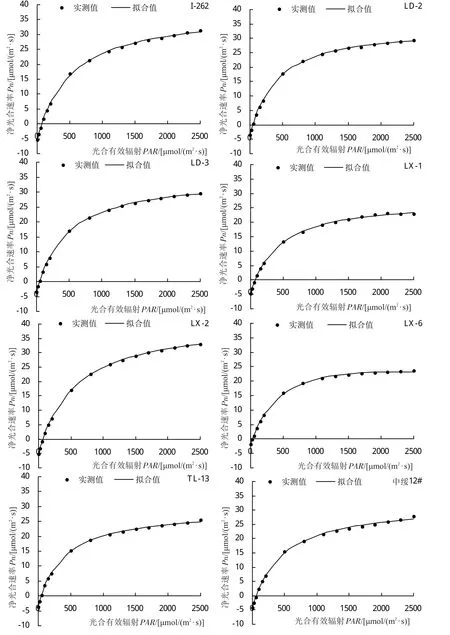
圖1 不同引種雜交楊光合-光響應(Pn-PAR)曲線
將所測得數據用直角雙曲線修正模型進行擬合分析(表2),不同引種雜交楊的最大凈光合速率(Pmax)變化范圍為22.12~41.40 μmol/(m2·s),其中‘LX-2’的Pmax值最大,‘CL-6’的Pmax值最小,二者較Pn日變化最高值分別高 2.88 μmol/(m2·s)、低 0.9 μmol/(m2·s),Pmax較Pn日變化最高值低的參試品系有‘108’、‘93-8-18’、‘CD-2’、‘CL-6’、‘D5-26’、‘DS-07-122’、‘DX-08-1’、‘DX-08-8’、‘LD-2’、‘LD-3’、‘LX-1’、‘LX-6’和‘中綏 12#’,其中‘93-8-18’、‘108’、‘CD-2’、‘D5-26’、‘DS-07-122’、‘LD-2’、‘LX-1’和‘LX-6’相差較大,相差幅度范圍為16.48%~29.10%,其余品系Pmax與Pn日變化最高值大致接近。AQY變化范圍為0.0243~0.0631,品系‘CD-2’具有最大AQY,說明‘CD-2’具有較強的光能轉化效率,而‘LX-6’最小,且在參試品系是唯一AQY低于0.03的品系,其在弱光條件下,二氧化碳同化能力較弱。參試品系暗呼吸速率(Rd)變化范圍為 1.14~5.23 μmol/(m2·s),‘I-262’具有最高Rd,‘CL-6’最低,參試品系Rd存在一定差異,其中‘93-8-18’、‘108’、‘CL-6’和‘LX-6’的Rd較小,而‘I-262’、‘LX-2’和‘D5-20’要高于其2~4倍,由此說明,‘I-262’、‘LX-2’和‘D5-20’等Rd較高的品系需要消耗更多的有機物以維持自身生命活動。參試品系LCP變化范圍為 16.16~70.17 μmol/(m2·s),參試品系開始進行光合作用的光照條件差異較大,‘93-8-17’、‘93-8-18’、‘108’、‘CL-6’和‘LX-6’的LCP低于 30 μmol/(m2·s),說明這 5 個品系對低光環境適應能力 強 ,‘I-262’、‘LX-1’、‘LX-2’則具有 較 高 的LCP,其對低光的適應能力較弱,這與Rd和AQY的分析結果一致。
3 結論
利用直角雙曲線修正模型擬合計算得到引種雜交楊的光合-光響應特征參數,‘LX-2’、‘84-306’、‘I-262’、‘D5-20’、‘D5-9’、‘DX-08-1’和‘DM-9-16’最大光合潛力較強;‘CL-6’、‘93-8-18’、‘108’、‘93-8-17’和‘LX-6’則更能適應弱光環境 ;‘CD-2’、‘D5-20’、‘D5-9’和‘93-8-17’對光能的轉化效率較高。伊犁地區光照強度大,適宜高光強的品系更能夠充分利用當地光熱資源用于自身生長,因此,最大光合潛力較強的品系更能夠適應伊犁地區高光環境,而對于光合潛力相對較小的品系,可在當地進行多年栽植馴化,使其盡可能適宜當地光熱環境,保留種質資源。
4 討論
光合-光響應模型是植物光合作用對光照強度響應模擬的系統分析,通過光合-光響應曲線的測定,能夠有效了解光合作用過程對光強的利用效率[7,12]。光合-光響應曲線的參數(Pmax、LSP、LCP、AQY、Rd)已成為植物速生豐產的判定依據[19]。
Pmax是植物最大光合潛力的直接標識,反映植物單位葉面積同化二氧化碳的最大潛力,在一定程度上反映了生長速度[20]。參試品系中以‘LX-2’的Pmax最大,較最低Pmax的‘CL-6’高出了19.28 μmol/(m2·s),兩者最大光合潛力相差甚大。本研究結果顯示,部分參試品系Pmax較日變化最高值小,甚至差值較大,江錫兵[21]認為其可能原因是影響光合作用的內部因子與外界環境相關,測定日變化進程中較低的RH使得葉片與大氣水汽壓差增大,葉片部分氣孔關閉導致Pn下降。本研究參試品系Pn日變化測定時間為8月上中旬,而光合-光響應曲線測定時間為8月中下旬,除環境水熱因子、光照條件變化外,植物本身的生理機能亦將隨著生長季的變化而變化,植物整體生理機能呈衰老下降狀態;另外,光合-光響應曲線測定過程中,對標記樣葉在設定光強下的誘導時間亦可能影響最終測定結果。
一般而言,植物葉片的LSP和LCP反映了植物光合作用對光照條件的要求,較高的LSP和較低的LCP說明植物對光環境較強的適應性,反之則說明植物對光環境適應性較弱。筆者采用的直角雙曲線修正模型對光合-光響應曲線進行擬合分析,發現得到的參試品系LSP普遍高于光合-光響應曲線PAR值設定范圍和實際野外光環境光照強度,原因可能在于,直角雙曲線修正模型對光合-光響應曲線出現光抑制現象的參試品系特征參數估算貼近實測值,但模型自身參數之間相互約束,有時不能直接得到趨于飽和型的光合-光響應曲線的光合特征參數LSP[12]。因此,不對參試品系光合-光響應特征參數LSP做分析。
植物的光補償點(LCP)反映植物對弱光的利用能力,其值越低,說明植物越能在弱光條件下進行最大限度的光合作用,是判定植物耐蔭性的重要指標[8]。參試品系LCP范圍較大,‘I-262’具有較高LCP,其在弱光條件下較引種雜交楊要弱,參試品系多數集中于30~60 μmol/(m2·s)之間。蔣高明[22]認為陰性植物 LCP 低于20 μmol/(m2·s),陽性植物LCP高于50 μmol/(m2·s),本研究結果認為,模型計算得到的LSP多數高于實際光環境測試最高值,由此不易判定參試品系的喜陰或喜陽屬性,光合-光響應曲線測定時間的選擇對計算結果有較大影響,雜交楊不同生長期的各光合特性參數不同,需進行不同生長階段的光合-光響應曲線測定來進行整個生長階段的分析。此外,參試品系存在一定程度的種間競爭和種內競爭,這可能對參試品系樣株有一定影響。
表觀量子效率(AQY)代表弱光條件下植物對光的吸收利用能力以及對蔭蔽環境的適應能力[23-24],一般情況下,植物的AQY介于0.03~0.07之間,其值越高說明植物葉片對光能的轉化效率越高,植物在早晚低光條件下光能利用能力就越強[25-26]。參試品系AQY介于0.0243~0.0631之間,多數品系AQY水平處于中上等,說明多數品系仍有較強的光能轉化效率,以‘CD-2’最高。
暗呼吸作用通過消耗代謝光合產物為植物生理活動提供能量,與植物的光合作用相輔相承。不同植物以及同種植物不同無性系之間的暗呼吸作用也存在差異,因而通常將暗呼吸速率作為評定優良品種的重要指標之一,通常情況下選擇低暗呼吸速率的品種,利于生產率的提高[20,27]。參試品系Rd介于1.14~5.23 μmol/(m2·s)之間,以最低為擇優的標準,‘93-8-18’、‘108’、‘CL-6’和‘LX-6’表現良好,而‘I-262’、‘LX-2’和‘D5-20’要高于其數倍,需消耗更多的有機物以維持其活躍的生理活動,不利于有機物的積累。
引種馴化和推廣栽培國內外優良林木種質資源,不斷提高林木品種的生態多樣性、遺傳多樣性和種質優異性是促進中國現代林業科學發展和生態文明建設的一項重要工作[28]。本研究對象為2016年從遼寧省楊樹研究所引種的22個楊樹品系,在新疆范圍內屬首次引進;新疆伊犁地區與遼寧省緯度相近,冬季寒冷,氣候相似,增加了引種的成功性。從光響應曲線看,‘LX-2’、‘84-306’、‘I-262’、‘D5-20’、‘D5-9’、‘DX-08-1’和‘DM-9-16’表現出的最大光合潛力并不弱于‘84-I-101’、‘銀×新4#’、‘銀×新6#’、‘銀×新192’等當地品種[29];生長特性(樹高、地徑)方面也表現良好[30]。后期要繼續加強引種適應性方面的觀測,主要病蟲害、抗旱性以及耐鹽堿方面的持續監測,為后期推廣良種奠定基礎。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
光學精密工程(2016年6期)2016-11-07 09:07:19
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
核科學與工程(2015年4期)2015-09-26 11:59:03
