兩類典型荒漠生物結皮覆蓋土壤碳通量月變化特征及其影響因子
2021-10-11 05:14:32韋應欣周利軍卜崇峰鞠孟辰李亞紅李炳垠
水土保持研究 2021年6期
關鍵詞:生物
韋應欣,周利軍,卜崇峰,駱 漢,鞠孟辰, 王 春,李亞紅,郭 琦,李炳垠
(1.西北農林科技大學 水土保持研究所,陜西 楊凌 712100;2.中國科學院 水利部 水土保持研究所,陜西 楊凌 712100; 3.長江水利委員會 長江流域水土保持監測中心站,武漢 430010;4.黃河水利委員會 黃河上中游管理局,西安 710021)
生物土壤結皮(Biological Soil Crust,BSC,簡稱生物結皮)是由光合自養型的藍藻、綠藻、地衣、苔蘚類植物和異養型的真菌、細菌、古細菌等生物群落,通過菌絲體、假根和分泌物等與表層土壤顆粒膠結而成的復合體,結皮類型以蘚結皮和藻結皮為主,蘚結皮是以苔蘚類植物為優勢組成的生物結皮,通常呈綠色、金黃色、棕色或黑色,藻結皮是以藍藻、綠藻為優勢組成的生物結皮,通常比周圍土壤的顏色黑,覆蓋了旱區約40%的地表面積[1]。生物結皮作為具有光合活性的土壤—大氣系統界面層,在光照條件下,固定大氣中的CO2,其光合固定的碳是旱地碳輸入的重要來源[2],同時,也能通過其自身所包含的有機成分與生命體的呼吸作用向大氣中釋放CO2[3]。因此,生物結皮是影響干旱與半旱區生態系統碳循環過程的重要因素[4-5]。
在干旱半干旱區生態系統中,水分是主要的調控和限制因子,能夠直接影響生物結皮的生理活性[6-8]。旱地土壤表面濕潤時間的長短決定了生物結皮覆蓋土壤的碳源—匯效應[6]。此外,生物土壤結皮存在凈碳固定的“水分閾值”,低于閾值時出現碳損失,較好的水分條件有利于生物結皮的光合固碳過程[8]。水分充足條件下,溫度是影響生物結皮覆蓋土壤碳通量的關鍵因子[9]。高溫條件下生物結皮的呼吸活性強于光合活性,使其CO2排放量增加,凈碳吸收量減少,低溫條件下則相反[7,9]。此外,溫度可以通過影響結皮層含水量與濕潤周期間接影響生物結皮的光合、呼吸過程[6,9-13],高溫會增加結皮層水分蒸發,縮短其代謝活動時長,從而減少凈碳輸入量[13-14]。根據氣候變化情景預測,未來世界上大多數地區的氣溫將升高,導致降水模式發生變化,干旱期延長[15-16],可能使生物結皮在未來65 a內覆蓋面積減少25%~40%[17],氣候變暖與生物結皮覆蓋度下降的協同作用將削弱旱地吸收大氣CO2的能力,在中長期內減少土壤碳儲量[18]。因此量化生物結皮覆蓋土壤碳通量對大氣、土壤溫度與濕度等環境因子的響應,對于預測旱地生態系統碳儲量的變化以及了解全球碳循環和平衡的意義至關重要[19]。
當前國內外關于生物結皮覆蓋土壤碳通量的研究主要集中于極地和苔原地區、美國西南部莫哈維沙漠、科羅拉多高原、以色列內蓋夫沙漠和我國的古爾班通古特沙漠、騰格里沙漠、黃土高原等地區[3,20]。這些地區的研究對生物結皮覆蓋土壤碳通量的影響因素探討不足[21],且多集中于室內模擬研究,原位觀測數據仍然較少[11],不同類型生物結皮覆蓋土壤碳通量研究還很匱乏[11],多數研究只關注生物結皮覆蓋土壤碳損失或碳固定的某一方面[2,20-21],很少綜合考慮多種環境因素對生物結皮覆蓋土壤碳通量的影響,并探討區域氣候變化和土地利用方式對生物結皮覆蓋土壤碳通量的影響。這些問題極大地限制了旱區土壤碳交換的估算,及未來氣候變化對土壤與大氣碳交換影響的預測[22]。毛烏素沙地位于我國農牧交錯區,農業、畜牧業發達、盛產石油、煤炭等資源,是生態恢復及評價實踐研究的重點區域。然而該區目前還沒有生物結皮覆蓋土壤碳通量月動態變化及其驅動因子的研究。鑒于此,我們選擇毛烏素沙地中處于生長季(6—11月,水、溫因子變化劇烈)的蘚結皮、藻結皮,進行兩類生物結皮覆蓋土壤的呼吸速率(DR)、凈碳通量(NCF)、光合速率(Pn)以及土壤溫度、含水量、光照強度等相關環境因子的動態檢測,以了解荒漠生態系統中蘚結皮、藻結皮的碳通量月變化特征及其與環境因子的關系,探討氣候變化對兩類生物土壤結皮覆蓋下碳通量的潛在影響,為該區生物土壤結皮的碳交換研究提供參考資料。
1 研究區概況
研究區地處內蒙古自治區鄂托克前旗敖勒召其鎮(東經107°50′40.51″,北緯38°05′53.39″),海拔1 344 m,屬于毛烏素沙地西南部。該區氣候干旱少雨,風大沙多,日照充足,屬中溫帶溫暖型干旱、半干旱大陸性氣候。年均氣溫7.1℃,最低月平均氣溫-10.2℃,最高月平均氣溫22.4℃。降水年際變率大,時空分布不均勻。年平均降水量265 mm,主要集中在6—9月,占全年降雨量的70%,年均蒸發量2 514.8 mm。樣地位于固定沙丘頂部,樣地內土壤類型為風沙土,植被種類較少,主要為沙蒿(Artemisiaordosica)、沙柳(SalixcheilophilaSchneid),蘚、藻兩類結皮發育良好,分布廣泛[23]。
2 方法與數據
2.1 研究方法
2.1.1 試驗設計 選取植被灌叢間發育較的蘚結皮、藻結皮(兩類結皮的覆蓋度接近100%,遠離植被冠層及其根系,距離植株2.5 m左右(圖1A—B)作為研究對象。在鄂托克前旗境內,為兩類結皮各選取1處地勢平坦、生境條件相似的樣地(30 m×30 m)。在樣地內生物結皮發育狀況相近、不受植被影響的地點設置3個2 m×2 m的樣方作為重復,共6個樣方,調查樣方內生物結皮生長狀況及其下伏土壤理化性質(表1)。為降低生物結皮發育狀況差異以及樣方周圍植被的干擾,試驗前1月(2016年5月初)移除樣方內除生物土壤結皮外其他植物,放置時先將生物土壤結皮潤濕,然后在每個樣方中心垂直嵌入1個PVC環(不透光,高10 cm,直徑15 cm,圖1A)和1個有機玻璃環(透光率>99%,高10 cm,直徑15 cm,圖1B),PVC環和有機玻璃環插入深度為6 cm,插入時保證環內結皮完整,以利于擾動土壤盡快恢復,降低測定誤差。表1為調查樣方內的各項基本指標。

圖1 土壤碳通量觀測樣點
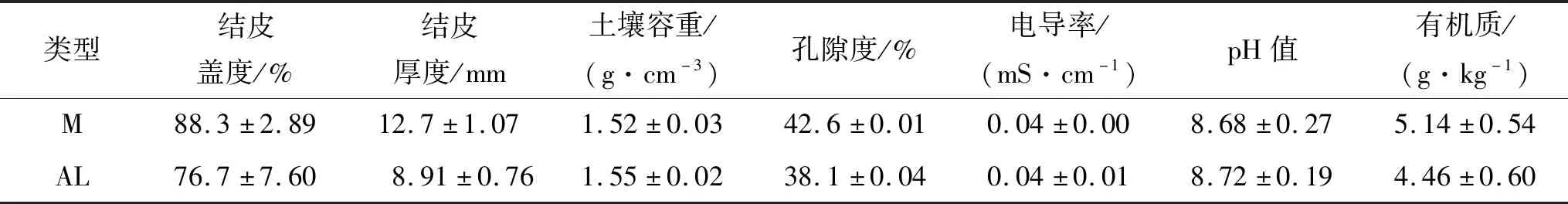
表1 樣點生物土壤結皮發育狀況及下伏土壤理化性質
2.1.2 指標觀測 試驗開展于2016年6—11月,為兩類生物結皮的生長期,采用LI-8100自動土壤CO2通量系統(LICOR,Lincoln,Nebraska,USA,圖1C)連接便攜式測量室8100-103和有機玻璃氣室LI-104測定BSC土壤CO2通量[24]。在PVC環內,用LI-8100系統連接便攜式測量室8100-103(氣室使用金屬材質,閉合時,密閉遮光,在黑暗條件下測定生物結皮層呼吸作用和下覆土壤呼吸作用總和,圖1C)測定兩類生物結皮覆蓋土壤呼吸速率(DR);在有機玻璃環內,用LI-8100系統連接有機玻璃氣室LI-104(氣室采用有機玻璃制成,透光率>99%,在光照條件下測定生物結皮層光合作用、呼吸作用以及下伏土壤的呼吸作用總和,圖1C)測定兩類生物結皮覆蓋土壤凈碳通量(NCF)。每月選取晴朗的天氣(測定3 d前無降雨事件),觀測時間為7:00—19:00,觀測頻次為每2 h一次,每次觀測3 min,包括測量時長90 s,測量前與測量后氣體平衡時間各45 s,測定前,去除處理樣方中的雜草,以降低測定誤差。
兩類生物土壤結皮的光合速率(Pn)計算公式為[3]:
Pn=DR-NCF
式中:DR為土壤呼吸速率;NCF為凈碳通量。Pn為正且光合速率越高,表示結皮的光合速率越高,碳吸收能力越強。
同時,用數字式地溫計(MannixTM,USA)測定土壤2,5,10 cm處的土壤溫度(ST);用TDR(TRIMEPICO32)測定土壤2,5,10 cm處的土壤含水量(SWC);用數字式溫濕度計(GM1361)測定近地表空氣溫度(AT)和空氣濕度(AM);用數顯照度計(TES1339)測定光照強度(Li)。
2.2 數據處理
消除光照強度變化對蘚結皮與藻結皮光合速率影響的誤差,光照強度采用了蘚結皮與藻結皮樣方內多次測量結果的均值。采用逐步回歸分析法對兩類生物結皮覆蓋土壤碳通量與環境因素(2,5,10 cm土壤溫度、2,5,10 cm土壤含水量、光照強度、近地表空氣溫濕度)進行多元線性回歸分析。采用了典范對應分析(CCA)評價了環境參數解釋兩類結皮的光合速率及其覆蓋下的土壤碳通量變化的能力。基于最大調整R2和最小AIC(Akaike Information Criterion,赤池信息量原則)選擇了最佳擬合的CCA模型。擬合的最佳CCA模型中所有變量的方差膨脹因子(VIF)均低于10。采用線性模型擬合兩類結皮覆蓋土壤凈碳通量與土壤溫度、含水量的關系。數據分析與繪圖采用R軟件(4.0.2版本)和R-studio(1.3.1093版本)完成。
3 結果與分析
3.1 生物結皮覆蓋土壤碳通量月變化特征
如圖2所示,蘚結皮(M)、藻結皮(AL)的土壤呼吸速率(DR)月變化為先升后降,在8月出現峰值[M:1.96 μmol/(m2·s);AL:1.66 μmol/(m2·s)];光合速率(Pn)月變化為先升后降,9月出現峰值[M:2.75 μmol/(m2·s);AL:1.39 μmol/(m2·s)];凈碳通量(NCF)月變化為先升后降再升,7月出現峰值[M:1.38 μmol/(m2·s);AL:1.43 μmol/(m2·s)],9月出現谷值[M:-1.24 μmol/(m2·s);AL:-0.18 μmol/(m2·s)]。
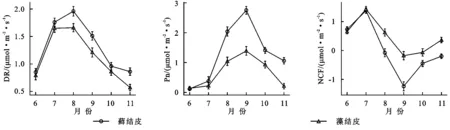
圖2 兩類結皮覆蓋土壤碳通量月變化特征
生長期中,蘚結皮覆蓋土壤呼吸速率與光合速率顯著(p<0.05)高于藻結皮,凈碳通量顯著低于藻結皮。兩類生物結皮覆蓋土壤凈碳通量均大于0,表現為凈碳釋放(表2)。

表2 不同處理碳通量的差異 μmol/(m2·s)
3.2 生物結皮覆蓋土壤環境因子月變化特征
土壤溫度(ST)、土壤含水量(SWC)、光照強度(Li)、空氣溫度(AT)、空氣濕度(AM)的月變化均為“先升后降”的趨勢,土壤溫度在7月達峰值,光照強度、近地表空氣濕度、近地表空氣溫度均8月達峰值,土壤含水量在9月達峰值(圖3)。
蘚結皮覆蓋土壤含水量、近地表空氣濕度均顯著(p<0.05)高于藻結皮,而土壤溫度、近地表空氣溫度均顯著低于藻結皮(表3)。

表3 不同處理環境因子的差異
3.3 生物結皮覆蓋土壤碳通量的關鍵影響因子
逐步回歸分析(表4)顯示:5,10 cm土壤溫度、2 cm土壤含水量、光照強度是影響蘚結皮覆蓋土壤呼吸速率的關鍵因子;2,5 cm土壤溫度,5,10 cm土壤含水量,以及光照強度是影響藻結皮覆蓋土壤呼吸速率的關鍵因子。2 cm土壤含水量是影響蘚結皮光合速率的關鍵因子;2 cm土壤溫度、2,5 cm土壤含水量、近地表空氣溫度是影響藻結皮光合速率的關鍵因子。2 cm土壤含水量是影響蘚結皮土壤覆蓋土壤凈碳通量的關鍵因子;2 cm土壤溫度、5 cm土壤含水量、近地表空氣溫度是影響藻結皮土壤覆蓋凈碳通量的關鍵因子。

表4 兩類生物結皮覆蓋土壤碳通量與環境因子的逐步回歸方程
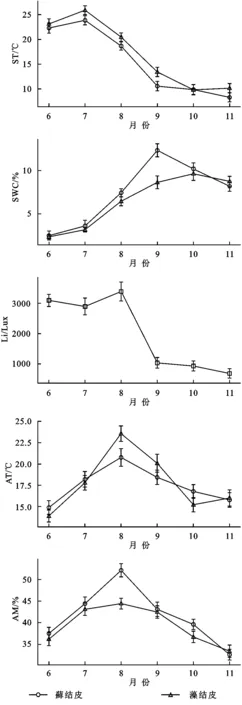
注:蘚結皮與藻結皮光照強度(Li)一致,具體原因詳見實驗方法。
3.4 生物結皮覆蓋土壤碳通量與環境因子的關系
CCA分析結果(圖4,表5)顯示:僅第一排序軸(CCA1)就解釋了蘚、藻兩類生物結皮與環境關系的100%,蒙特卡洛置換檢驗通過(p=0.001),環境因子解釋了蘚結皮覆蓋土壤碳通量變異的58%,解釋了藻結皮56%,對于兩類結皮覆蓋土壤碳通量與環境因子關系的還原能力和預測能力均較強。所有的環境因子中,2,5,10 cm土壤溫度,2,5 cm土壤含水量是影響蘚結皮土壤覆蓋土壤碳通量的關鍵因子;2,5,10 cm土壤溫度,光照強度、空氣濕度是影響藻結皮覆蓋土壤碳通量的關鍵因子(表5)。
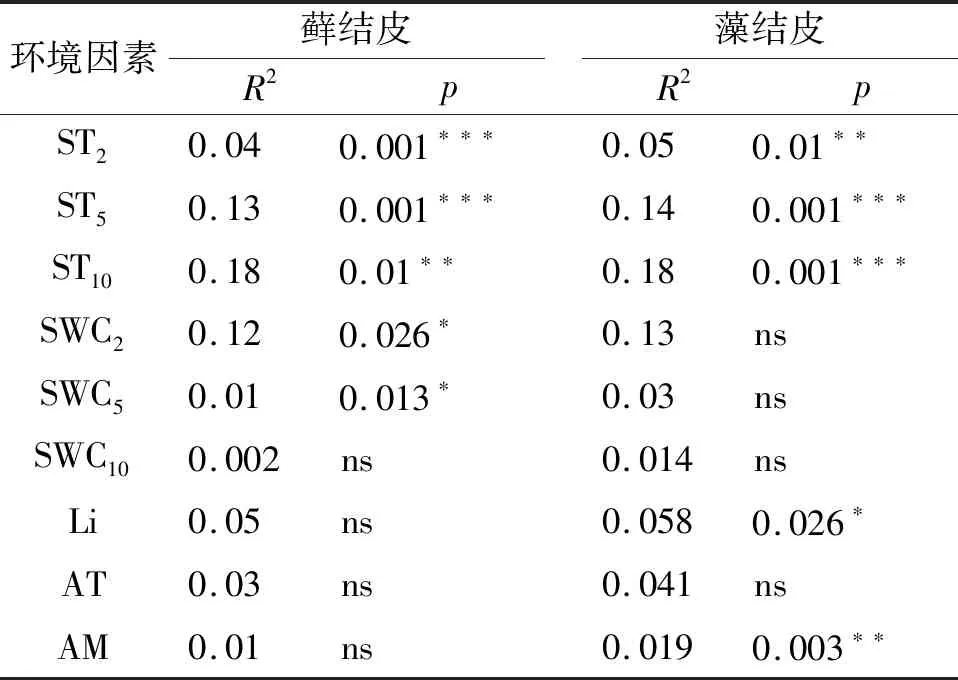
表5 兩類結皮覆蓋土壤碳通量與環境的分析結果
3.4.1 呼吸速率與環境因子的關系 2 cm土壤含水量與蘚結皮覆蓋土壤呼吸速率(DR)負相關,與藻結皮正相關;2,5,10 cm土壤溫度及5,10 cm土壤含水量與兩類結皮覆蓋土壤的呼吸速率均正相關;光照強度與蘚結皮覆蓋土壤呼吸速率正相關,與藻結皮相關性較弱;近地表空氣溫度與藻結皮覆蓋土壤呼吸速率正相關,與蘚結皮相關性較弱;近地表空氣濕度與藻結皮覆蓋土壤呼吸速率正相關,與蘚結皮相關性較弱(圖4)。
3.4.2 光合速率與環境因子的關系 兩類結皮光合速率(Pn)均與2 cm土壤含水量正相關,與2,5,10 cm土壤溫度、5,10 cm土壤含水量負相關;光照強度與蘚結皮光合速率負相關,與藻結皮正相關;近地表空氣溫度與蘚結皮光合速率負相關,與藻結皮相關性較弱;近地表空氣濕度與蘚結皮光合速率正相關,與藻結皮負相關(圖4)。
3.4.3 凈碳通量與環境因子的關系 兩類生物結皮覆蓋土壤凈碳通量(NCF)與5,10 cm土壤溫度、光照強度、近地表空氣溫度正相關,與2,5,10 cm土壤含水量、近地表空氣濕度負相關(圖4)。
3.5生物結皮覆蓋土壤凈碳通量變化對土壤溫度、含水量變化的響應
CCA分析顯示(圖4),2 cm土壤含水量(SWC2),10 cm土壤溫度(ST10)是兩類結皮覆蓋土壤碳通量的關鍵影響因素。擬合ST10,SWC2與兩類生物結皮覆蓋土壤凈碳通量的關系,對比多種線性與非線性的擬合模型,發現線性模型的擬合度與顯著性最高,能較好的擬合變量間的關系。其他土層溫度與含水量對兩類結皮覆蓋土壤碳通量影響較小,或與SWC2,ST10對生物結皮覆蓋土壤凈碳通量的擬合曲線變化規律相似,不再贅述。
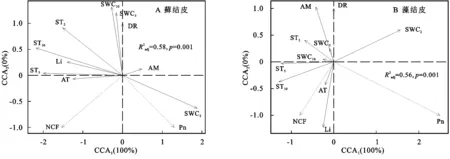
圖4 兩類結皮覆蓋土壤碳通量與環境因子的CCA排序
3.5.1 對土壤溫度變化的響應 線性回歸分析結果(圖5)顯示,蘚結皮、藻結皮覆蓋下土壤凈碳通量(NCF)與10 cm土壤溫度(ST10)呈線性正相關關系(蘚結皮:R2=0.55,p<0.01;藻結皮:R2=0.56,p<0.01)。兩類結皮覆蓋下土壤凈碳通量存在凈碳固定(NCF<0,生物結皮開始凈碳吸收的土壤溫度值,高于此值出現碳損失現象)的土壤溫度閾值。蘚結皮的閾值(ST=13.5℃)高于藻結皮(ST=7.3℃)。
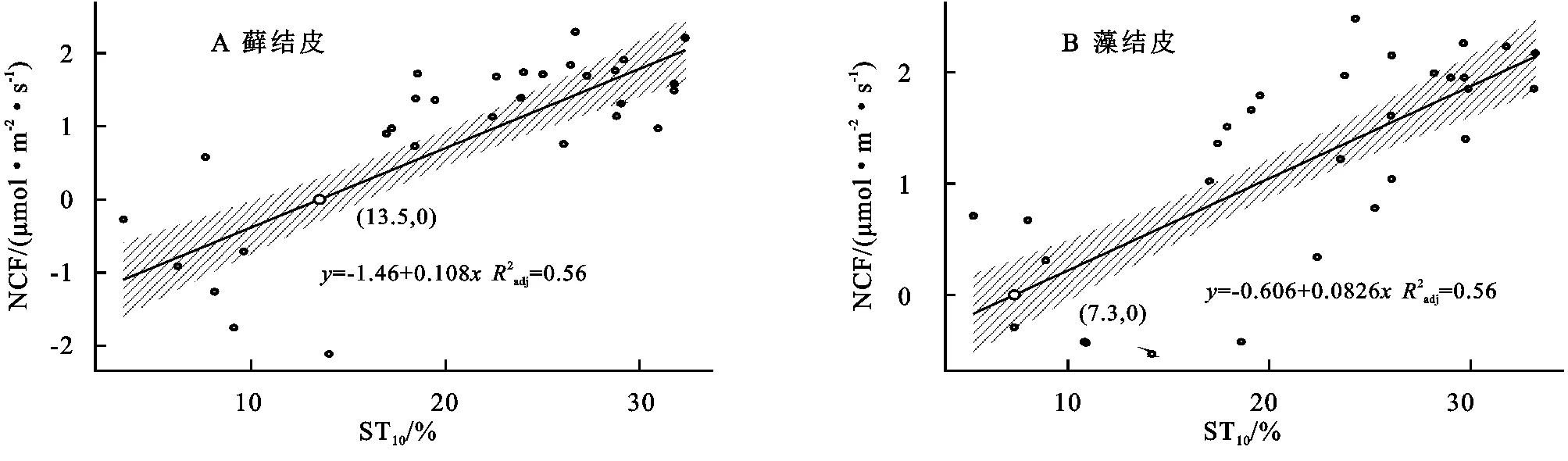
圖5 兩類結皮覆蓋土壤凈碳通量與10 cm土壤溫度的關系
3.5.2 對土壤含水量變化的響應 線性回歸分析結果(圖6)顯示,蘚結皮、藻結皮覆蓋下土壤凈碳通量(NCF)與2 cm土壤含水量(SWC2)呈線性負相關關系(蘚結皮:R2=0.34,p<0.01;藻結皮:R2=0.49,p<0.01)。兩類結皮覆蓋下土壤凈碳通量存在凈碳固定(NCF<0,生物結皮開始凈碳吸收的土壤含水量值,低于此值出現碳損失現象)的土壤含水量閾值。蘚結皮的閾值(SWC2=8.9%)高于藻結皮(SWC10=11.4%)。
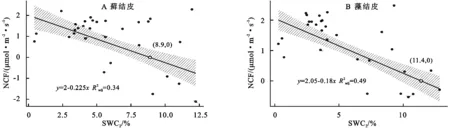
圖6 兩類結皮覆蓋土壤凈碳通量與2 cm土壤含水量的關系
3 討 論
3.1 生物結皮覆蓋土壤碳通量月動態變化及其驅動因子
兩類生物結皮覆蓋土壤的呼吸速率月動態變化為先升后降,在光照強烈、土壤含水量適中的8月達到峰值(圖2—3)。在騰格里沙漠,Guan等[21]也發現類似規律:蘚、藻兩類結皮覆蓋土壤呼吸速率在土壤溫度最高、降雨適中的7月達到峰值。逐步回歸(表4)與CCA分析(圖3,表4)表明:5,10 cm土壤含水量,2,5,10 cm土壤溫度以及光照強度與兩類生物結皮土壤覆蓋呼吸速率正相關性較強,是驅動兩類結皮土壤呼吸速率月動態變化的關鍵因子。土壤溫度的升高會刺激土壤酶活性,加速土壤有機質的分解[25]。土壤含水量的升高則會直接影響養分礦化速率和土壤微生物活性,改變土體氧化還原電位和氣體交換速率,進而改變土壤碳通量[26]。光照強度的升高(圖2)會促進兩類結皮的生理活性,為其自身呼吸提供了更多碳源[8]。8月前,干燥的生物結皮土壤已經積累了一定的呼吸底物,進入8月,土壤含水量上升、光照強度達到峰值,因此,兩類結皮覆蓋土壤呼吸速率迅速上升,并在8月達到峰值。生長期中,蘚結皮覆蓋土壤呼吸速率顯著高于藻結皮(表2)。不僅是因為蘚結皮比藻結皮有更強的呼吸活性,還存在其他影響因素。一方面蘚結皮比藻結皮能夠更長時間的保持水分有效性,減緩水分蒸發和下滲,維持其自身呼吸活性。另一方面蘚結皮比藻結皮覆蓋土壤孔隙度更大,黏粒和養分含量更高,更有利于其下伏土壤呼吸作用[3]。
兩類生物結皮的光合速率與土壤含水量月變化同步,先升后降,9月達到峰值。逐步回歸(表4)與CCA分析(圖4,表5)表明:2 cm土壤含水量與兩類生物結皮光合速率正相關性較強,是驅動兩類生物結皮光合速率月動態變化最關鍵的因子,此外2,5,10 cm土壤溫度、光照強度與兩類生物結皮光合速率負相關性較強,也是影響兩類結皮光合速率月動態變化的重要因子。進入9月,光照強度的大幅下降,土壤含水量的大幅上升(圖3),一方面使生物結皮土壤的濕潤周期變長,另一方面使土壤溫度與近地表空氣溫度處于較低的水平。因此兩類生物結皮的光合速率在9月達到峰值。盡管蘚結皮光合速率與光照強度顯示出了較強的負相關性(圖3),太陽輻射最為強烈的8月,蘚結皮光合速率仍然處于較高水平。這可能是因為水分充足時,蘚結皮的光合過程能夠很好地適應光照強度的變化。例如,趙允格等[13]發現光照對黃土丘陵區蘚類結皮的光合作用沒有抑制作用。同時,藻結皮光合速率在8月同樣處于較高水平,表明藻結皮光合作用過程受光照強度變化影響較小。例如,Karsten等[10]發現非洲南部干旱地區的熱沙漠和美國阿拉斯加州的冷沙漠的BSC中的4種絲狀綠藻在較低光照下光合產氧量依然較高,而高光照條件無抑制作用。藻結皮光合速率與光照強度存相關性較弱(圖4)也證實了這點。整個生長期中,蘚結皮光合速率顯著高于藻結皮。原因在于蘚結皮持水能力更強,葉綠素a,b含量高,其光合活性更高,能夠吸收更多的CO2。此外,蘚結皮覆蓋土壤含水量顯著高于藻結皮、土壤溫度顯著低于藻結皮(表3),更有利于其進行光合作用[6,26]。
兩類結皮覆蓋土壤凈碳通量月變化為先升后降再升,7月出現峰值,與土壤溫度月動態變化出現峰值的月份一致,9月出現谷值,與土壤含水量月動態變化出現峰值的月份一致。CCA分析表明(圖4),兩類生物結皮覆蓋土壤凈碳通量與5,10 cm土壤溫度、光照強度、近地表空氣溫度正相關性較強,與2,5,10 cm土壤含水量、近地表空氣濕度存在較強的負相關性較強。毛烏素沙地干濕季分明[27],在降雨集中、光照強烈、土壤溫度最高的7月(干季),兩類生物結皮覆蓋土壤的呼吸速率明顯高于光合速率,因此凈碳通量達到峰值,而在光照適宜、土壤含水量較高的9月(濕季),光合速率明顯強于呼吸速率,因此凈碳通量達到谷值(圖2—3)。整個生長期中,兩類結皮凈碳通量均大于0(表2),表明兩類生物結皮覆蓋土壤表現出碳源作用。可能是因為兩類生物結皮的光合速率較低,不能抵消土壤呼吸所致[3]。蘚結皮凈碳通量顯著低于藻結皮(表2)。表明蘚結皮的凈碳固定能力更強,能夠通過光合作用吸收更多自身及其下伏土壤呼吸產生的CO2。
3.2 生物結皮覆蓋土壤凈碳通量對氣候變化的潛在響應
安莉娟[28]研究顯示:未來毛烏素沙地氣溫呈上升趨勢,年降水減少,干旱加劇。兩類生物結皮覆蓋土壤凈碳通量的變化與土壤溫度及近地表空氣溫度升高正相關較強,與2 cm土壤含水量及近地表空氣濕度負相關性較強(圖4),表明氣溫的上升與降雨量的減少會增加兩類生物結皮碳損失的風險。雖然生物結皮光合過程對溫度升高的響應具有彈性[29],但鑒于適宜研究區兩類生物結皮凈碳固定(NCF<0)的土壤溫度范圍較窄(圖5),兩類生物結皮的群落組成及其對環境因子的生理適應能力未發生變化時,增溫減雨會導致生物結皮覆蓋土壤凈碳通量有升高,使其釋放的CO2更難被光合固定的CO2取代,進入“C饑餓”的狀態。蘚結皮凈碳固定的土壤溫度閾值(13.5℃)顯著高于藻結皮(7.3℃)(圖5),土壤水分閾值(8.9%)明顯低于藻結皮(11.4%)(圖6),表明增溫減雨的氣候變化更可能導致藻結皮覆蓋土壤的碳損失,因此未來需要關注野外環境中生物土壤結皮碳通量與環境因子之間復雜的聯結關系[6],以明確全球氣候變化下生物土壤結皮的碳效應。
4 結 論
蘚結皮與藻結皮覆蓋土壤碳通量月變化相似,并與土壤溫度、水分的月變化同步,其中呼吸速率和光合速率的月變化分別與土壤溫度、含水量月變化同步,表現為“先升后降”的趨勢,凈碳通量則受到了土壤溫度、含水量的交互作用,表現為“先升后降再升”的趨勢。兩類結皮的凈碳釋放量隨淺層含水量的增加而降低,隨深層土壤溫度的增加而升高,光照強度通過改變土壤、大氣溫度、濕度間接影響兩類結皮覆蓋土壤的碳通量。在相同的溫度與水分范圍內,蘚結皮的凈碳固定能力比藻結皮更強。蘚結皮的凈碳固定土壤水分閾值明顯低于藻結皮,土壤溫度閾值明顯高于藻結皮,使其在全球氣候變化引起的增溫減雨的背景下更不易發生碳損失。未來應結合室內模擬試驗長期原位觀測生物結皮土壤碳通量與環境因子間的相互作用,以反映生物結皮碳通量與環境因子間的復雜關系。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
