東太平洋赤道海域鳶烏賊(Sthenoteuthis oualaniensis)和莖柔魚(Dosidicus gigas)的食性比較研究*
2021-10-11 09:53:18李云凱高小迪陳新軍
海洋與湖沼 2021年5期
李云凱 馮 丹 高小迪 陳新軍 貢 藝①
(1. 上海海洋大學海洋科學學院 上海 201306; 2. 農業農村部遠洋與極地漁業創新重點實驗室 上海 200009; 3. 青島海洋科學與技術試點國家實驗室海洋漁業科學與食物產出過程功能實驗室 青島 266237; 4. 大洋漁業資源可持續開發省部共建教育部重點實驗室 上海 201306)
柔魚科(Ommastrephidae)頭足類動物廣泛分布在除北冰洋以外的全球各大洋, 是海洋生態系統的重要生態類群, 也是頭足類商業捕撈中產量最高的種類(De La Chesnaiset al, 2019)。鳶烏賊(Sthenoteuthis oualaniensis)和莖柔魚(Dosidicus gigas)同屬柔魚科、柔魚亞科, 是各國頭足類商業捕撈的主要物種(陳新軍, 2004; 劉金立等, 2019; 陳新軍等, 2019)。鳶烏賊分布于印度洋和太平洋的赤道與亞熱帶海域(38°N—40°S) (Roperet al, 1984), 而莖柔魚分布于北至美國加利福尼亞(30°N), 南至智利圣地亞哥(45°S)的東太平洋海域, 并在赤道向西延伸到125°W。兩種頭足類在東太平洋赤道海域呈同域分布的狀態(Staafet al, 2010)。此外作為近緣種, 兩種頭足類外部形態相似, 且都具有生長快、攝食量大和晝夜垂直洄游等生物學特性(Bazzinoet al, 2010; Staafet al, 2010; 貢藝等, 2019; 韓飛等, 2019)。根據穩定進化對策(evolutionary stable strategy), 近緣種會通過調節自身的攝食策略來減少食物競爭, 從而可能達到平衡而共存(Schoeneret al, 1974)。前期利用穩定同位素和脂肪酸分析技術研究發現, 同域分布的鳶烏賊和莖柔魚存在營養生態位分化的現象(李云凱等, 2020)。但因缺少穩定同位素基線和餌料生物的穩定同位素和脂肪酸組成信息, 為了更直觀反映鳶烏賊和莖柔魚的攝食習性及其差異, 本研究擬利用胃含物分析法對兩種頭足類的餌料生物進行分析和比較。由于柔魚科頭足類特殊的攝食方式, 食物會被其角質顎(beak)和齒舌(radula)撕咬、磨銼成糜狀或碎塊狀, 影響對食物組成的準確判斷(Braleyet al, 2010)。國內外學者已開始運用DNA 條形碼技術輔助識別頭足類的食物組成(龔玉艷等, 2018; 劉夢娜等, 2020; Boet al, 2020)。據此, 本文以東太平洋赤道海域的鳶烏賊和莖柔魚為研究對象, 通過結合傳統胃含物分析和DNA 條形碼技術對二者的食物組成開展研究, 分析同域分布頭足類近緣種食性特化現象, 為解析頭足類近緣種共存機制提供數據支撐。
1 材料與方法
1.1 樣品采集與處理
鳶烏賊和莖柔魚均采自中東太平洋赤道海域(圖1),由科學觀察員2017—2019 年于我國遠洋魷釣船捕撈作業期間采集(表1)。樣品采集后立即冷凍, 經冷凍運回實驗室后進行生物學解剖和測量, 包括胴長(mm)和體質量(g), 胃含物樣本編號裝袋后保存于95%的乙醇中。為減少采樣點時空分布、體型差異和樣本量對研究結果的潛在影響, 僅選擇相同月份捕獲且胴長和體質量相近的兩種頭足類樣品開展研究, 樣本量均為80 尾。
圖1 鳶烏賊與莖柔魚采樣站點圖Fig.1 Sampling locations of S. oualaniensis and D. gigas
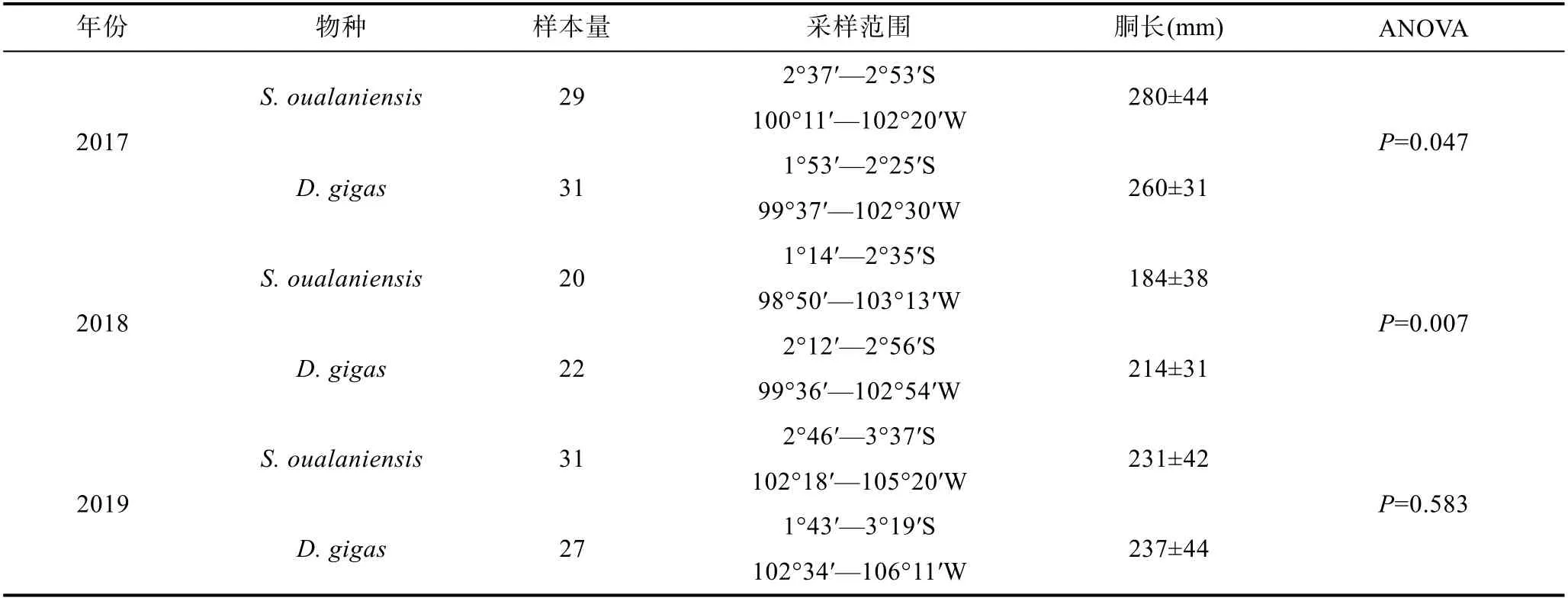
表1 鳶烏賊和莖柔魚采樣信息Tab.1 Sampling information of S. oualaniensis and D. gigas
1.2 胃含物分析
用無菌鑷子去除胃壁, 濾紙吸干多余的水分, 取全部胃含物稱重。在Olympuss ZX7 體視顯微鏡下對餌料生物進行鑒定并計數, 對照現場收集的餌料生物、海洋生物分類資料以及耳石圖譜庫等將胃含物盡可能鑒定至最低分類階元, 然后用濾紙將食物團表面水分吸干, 并用Shimadzu 精密電子天平(精確到0.000 1 g)分別稱重。暫未能鑒定的耳石和頭足類角質顎保存于95%乙醇中以便進一步鑒定, 未能通過形態鑒定的未知組織置于 95%乙醇溶液中冷凍保存,用DNA 條形碼技術進一步鑒定。
1.3 DNA 條形碼法鑒定餌料生物
取適量糜爛程度較小的食物塊, 用無菌水多次沖洗, 確保組織表面沒有胃液沉積后, 取100 mg 左右樣品置于新的滅菌離心管中, 用PBS 沖洗后換入新的離心管中, 使用無菌剪刀將組織剪碎用于DNA提取。采用海洋動物組織基因組DNA 提取試劑盒(天根)提取胃含物基因組DNA, 利用Nano Drop 2000 超微量分光光度計進行DNA 濃度測試。
用 Leray 等(2013)設計改進的 mlCOlintF、mlCOIintR 和Lco1490、Hco2198 兩對引物(Gelleret al,2013), 選取線粒體基因細胞色素 C 氧化酶亞基 I(mitochon-drial cytochrome oxidase subunit I, COI)作為分子標記進行PCR 擴增, 引物由生工(上海)生物公司合成, 序列見表2。
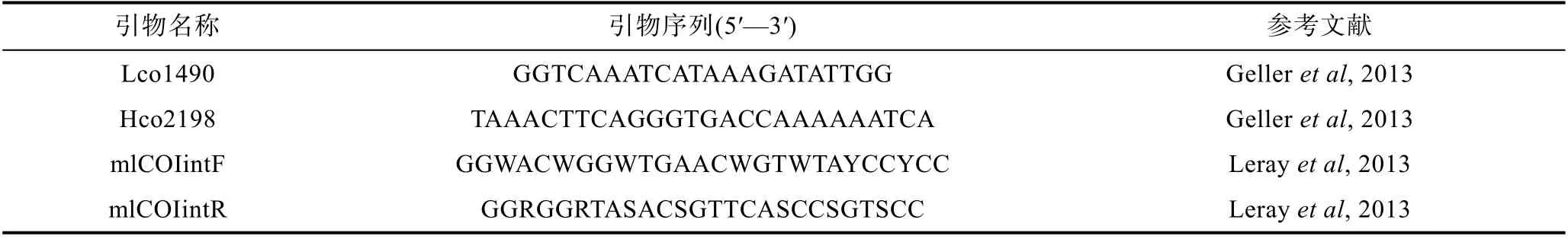
表2 PCR 引物信息Tab.2 Information of the PCR primers
PCR 擴增25 μL 體系: 6.5 μL H2O, 12.5 μL Premix Ex Taq 酶(TaKaRa) (2×), 正反向引物(0.25 μmol/L)各0.5 μL 以及DNA 模板5 μL。反應條件為: 95 °C 預變性5 min, 95 °C 變性10 s, 56 °C 退火30 s, 72 °C 延伸1 min, 退火溫度以1 °C 降序進行變性、退火、延伸16 個循環, 至退火溫度為46 °C 時, 重復25 次循環。72 °C 延伸10 min, 4 °C 結束反應(劉夢娜等,2020)。在1%的瓊脂糖凝膠電泳之后, 將PCR 產物送生工(上海)生物公司進行純化、反向測序。DNA 測序結果在 NCBI (National Center for Biotechnology Information)網站進行核酸比對, 對于魚類及頭足類,將BLAST (Basic Local Alignment Search Tool)最高匹配相似度超過99%的餌料生物確定到種, 低于99%高于90%的確定到屬(Wardet al, 2005); 對于甲殼類,將超過90%的餌料生物確定到種, 低于90%高于80%的確定到屬(Costaet al, 2007; Xuet al, 2019; 劉夢娜等, 2020)。
1.4 數據處理
評價某一頭足類胃含物中各餌料重要性的指標,主要包括個數百分比N%、出現頻率F%、餌料質量百分比W%、相對重要指數IRI, 相對重要性指數百分比IRI% (Hyslop, 1980)和Shannon-Wiener 多樣性指數, 計算公式如下:

式中,Pi為第i種餌料生物在食物組成中的數量百分比。
2 結果
2.1 食物組成
鳶烏賊和莖柔魚的胴長范圍分別為143—380 mm和172—380 mm, 二者無顯著差異(ANOVA,F=0.128,P=0.72)。體質量分別為101—1 552 g 和165—1 577 g,也無顯著差異(ANOVA,F=0.86,P>0.05)。
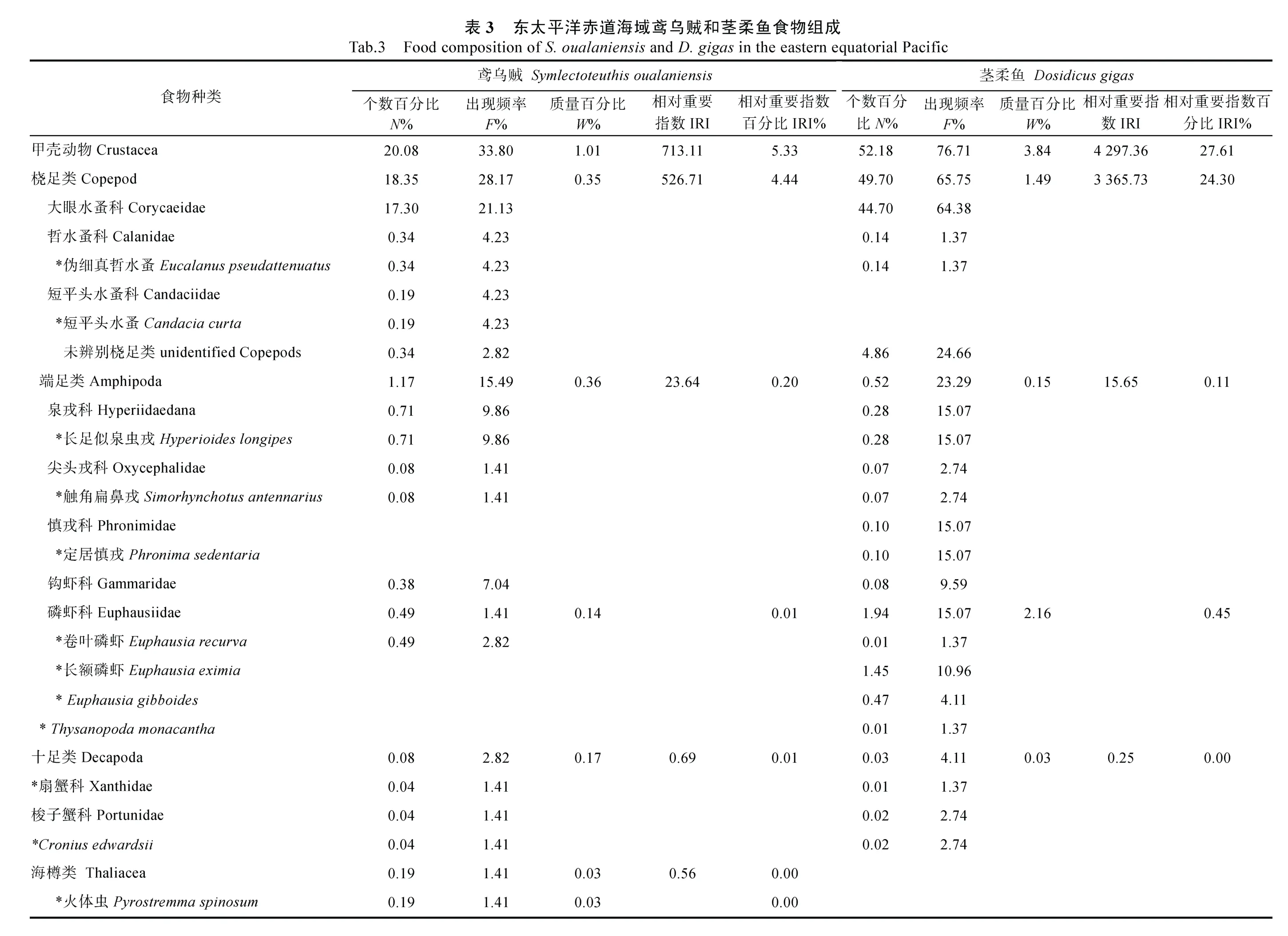
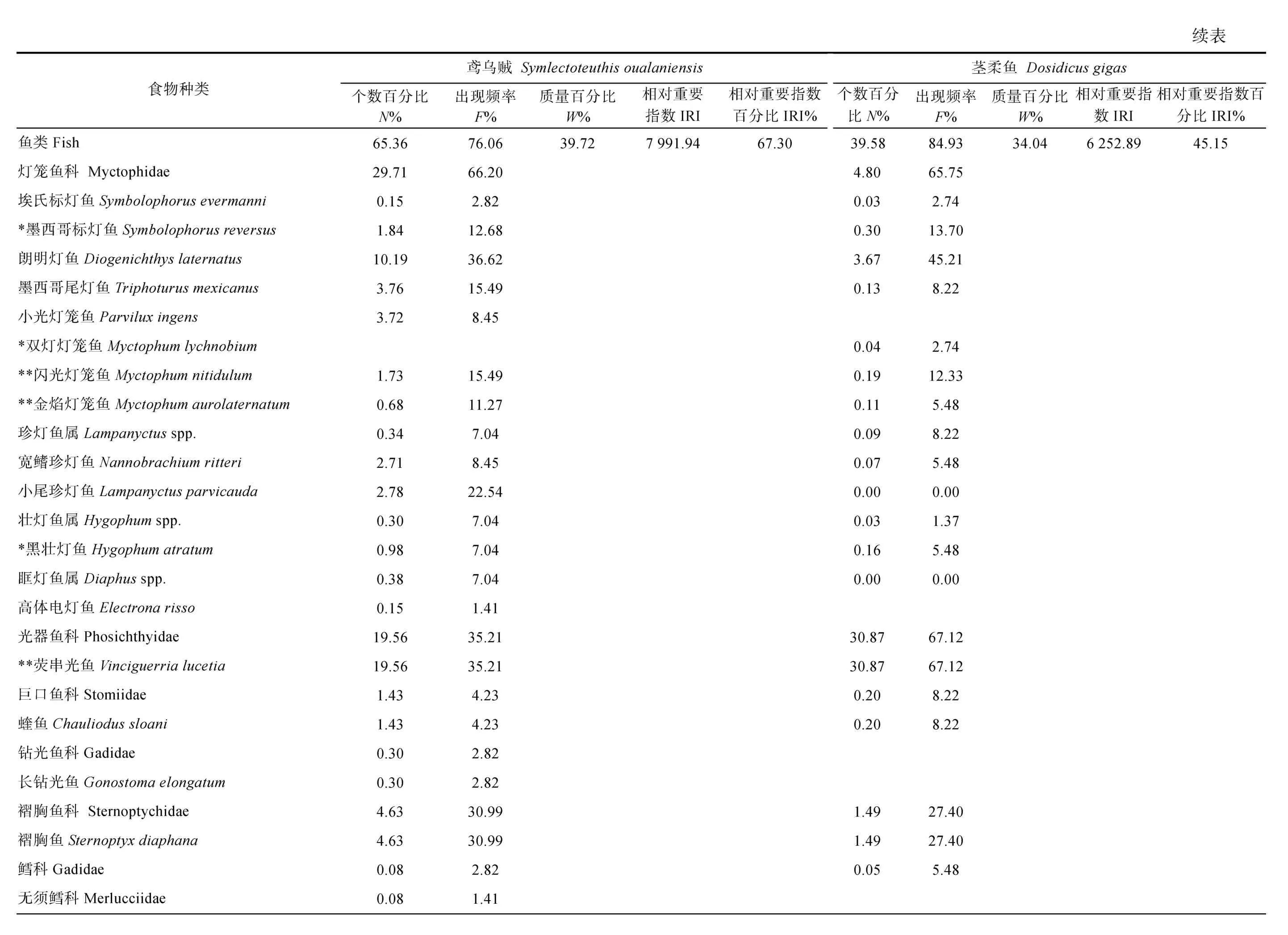
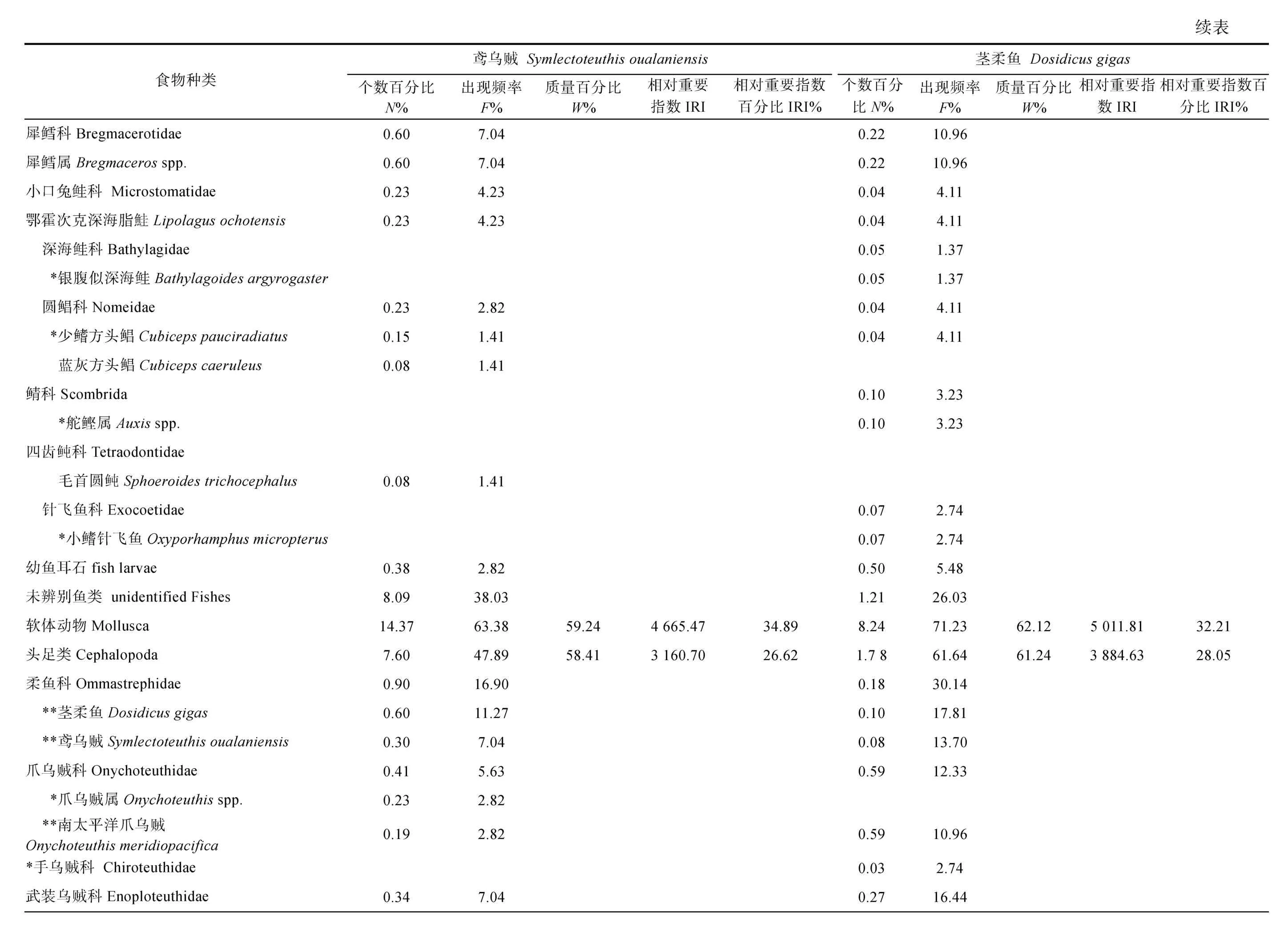
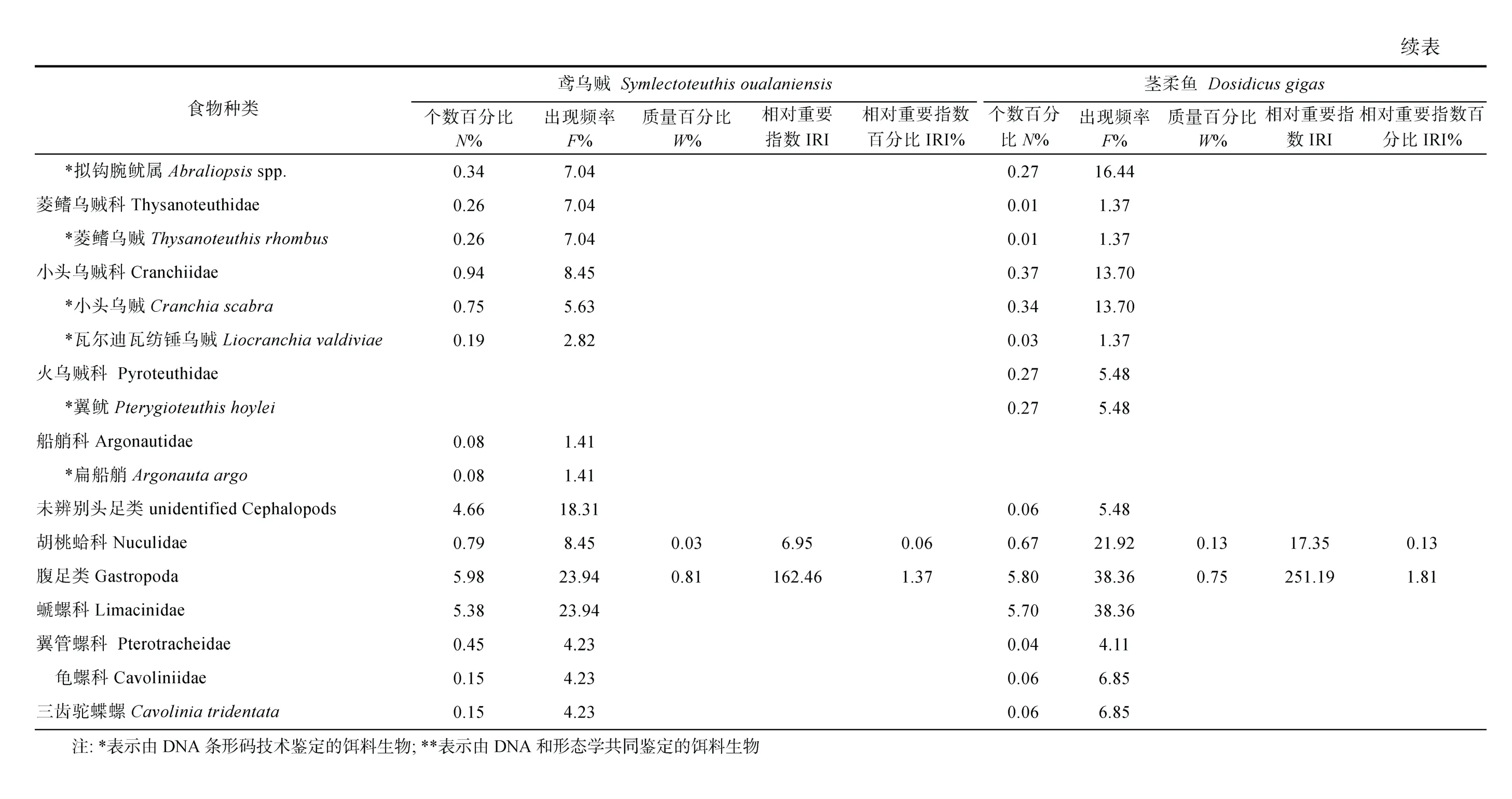
對鳶烏賊和莖柔魚的傳統胃含物分析共鑒定出餌料生物31 種, 包括16 種魚類、3 種頭足類和1 種腹足類可直接鑒定到種, 4 種魚類可鑒定至屬, 2 種魚類、2 種甲殼類和3 種軟體動物可鑒定至科(表3)。而DNA 條形碼鑒定共識別出33 種, 包括7 種魚類、6種頭足類、9 種甲殼類和1 種海樽類, 剩下的3 種魚類、5 種頭足類和2 種甲殼類可鑒定至屬(表4)。兩種方法所獲結果經比對后發現, 鳶烏賊和莖柔魚共攝食餌料生物4 大類64 種, 分屬37 科, 包括甲殼動物、魚類、軟體動物和海樽類(表3)。甲殼動物主要為橈足類(Copepods)、端足類(Amphipoda)、磷蝦類(Euphausiacea)和十足類(Decapoda); 魚類主要包括燈籠魚科(Myctophidae)、光器魚科(Phosichthyidae)、巨口魚科(Stomiidae)、鉆光魚科(Gadidae)、鱈科(Gadidae)和圓鯧科(Nomeidae)等; 軟體動物主要包括頭足類(爪烏賊科Onychoteuthidae、手無賊科Chiroteuthidae、菱鰭烏賊科 Thysanoteuthidae 和小頭烏賊科Cranchiidae 等)、雙殼類(胡桃蛤Nuculidae)和腹足類(螔螺科Limacinidae、翼管螺科Pterotracheidae、龜螺科Cavoliniidae)。鳶烏賊和莖柔魚的餌料生物分別占其中的53 和50 種。
?
?
?
?
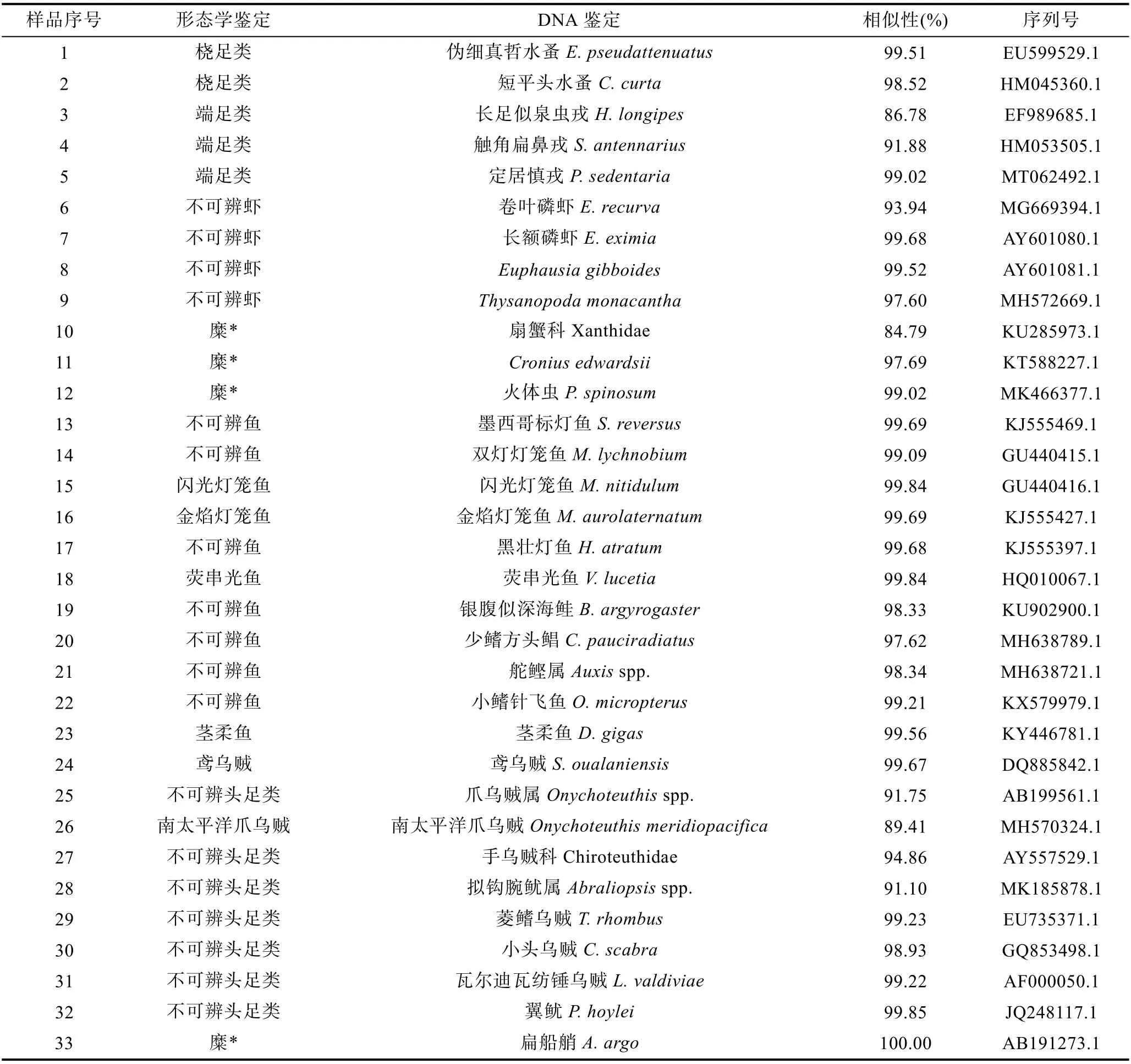
表4 部分形態學鑒定結果與DNA 分子鑒定對比Tab.4 Comparison of partial morphological identification results with DNA molecular identification
鳶烏賊胃含物中魚類的N%最高(65.36%), 以燈籠魚科(29.71%)和光器魚科(19.56%)為主, 其次是橈足類(18.35%)和頭足類(7.60%), 腹足類、端足類和磷蝦類等數量較少。而莖柔魚胃含物中橈足類的N%最高(49.70%), 其中大眼水蚤科(Corycaeidae)占比最高為 44.70%, 其次是魚類(39.58%)和腹足類(5.80%),磷蝦類、雙殼類和端足類等的數量較少。
對餌料生物出現次數的分析顯示, 鳶烏賊胃含物中F%最高的是魚類(76.06%), 并以燈籠魚科(66.20%)為主, 其次是頭足類(47.89%)和橈足類(28.17%)。莖柔魚胃含物同樣以魚類(84.93%)的F%最高, 其次是橈足類(65.75%)和頭足類(61.64%)。但是與鳶烏賊相比, 莖柔魚胃含物中F%最高的魚類屬于光器魚科(67.12%)。
從餌料質量看, 頭足類在鳶烏賊胃含物中的W%最高(58.41%), 其次為魚類(39.72%), 而腹足類和橈足類僅占0.81%、0.35%; 莖柔魚胃含物中頭足類的W%也最高(61.24%), 魚類(33.04%)次之, 腹足類和橈足類較鳶烏賊占比更大, 為1.75%和1.49%。
對比IRI 和IRI%可知, 魚類、頭足類和甲殼類是研究海域鳶烏賊(IRI>713.11; IRI%>5.33%)和莖柔魚(IRI>3884.63; IRI%>28.05%)最重要的餌料生物類群,而端足類、磷蝦類和海鱒類的IRI 和IRI%均較低。
2.2 種間差異
對各餌料指標差異性分析(ANOVA)結果顯示,餌料生物頭足類和魚類N%在鳶烏賊和莖柔魚間存在顯著差異, 但這兩類的W%無顯著性差異(表5)。甲殼類的N%和W%也無種間差異。此外, 各餌料生物的F%在鳶烏賊和莖柔魚間存在差異。甲殼類動物出現在約77%的莖柔魚胃中, 而鳶烏賊胃含物中甲殼類F%及N%均低于莖柔魚。此外, 在76%的鳶烏賊胃中發現了魚類, 占餌料總個數的65%, 就F%及N%而言,燈籠魚科是最豐富的餌料生物。雖然魚類在85%的莖柔魚胃中出現, 但N%僅40%, 且以光器魚科為主。為進一步比較相同取樣時間兩種頭足類的食性, 對個體大小相似的2019 年樣品進行分析(表1)。結果表明, 2019 年鳶烏賊和莖柔魚在魚類N%、W%和頭足類N%上存在顯著性差異, 與2017—2019 年的分析結果基本相符(表5)。
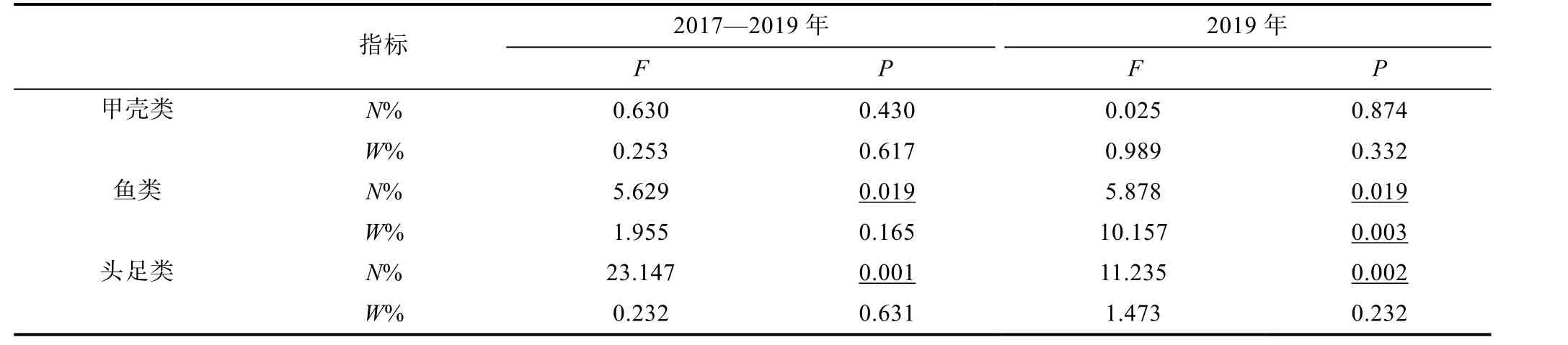
表5 鳶烏賊和莖柔魚食性差異性比較Tab.5 Comparison of dietary differences between S. oualaniensis and D. gigas
總體而言, 鳶烏賊(H=2.79)餌料生物的多樣性要高于莖柔魚(H=1.56; ANOVA,F=4.820,P=0.030)。多維尺度分析(multidimensional scaling analysis, MDS)也發現鳶烏賊食性更泛化, 莖柔魚個體間食性較相似(圖2)。
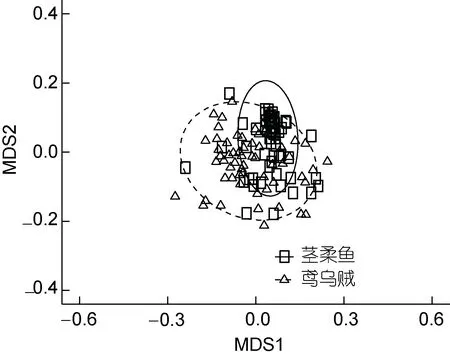
圖2 鳶烏賊和莖柔魚食物組成多維尺度分析Fig.2 MDS analysis of S. oualaniensis and D. gigas food composition
根據SIMPER 分析結果(圖3), 鳶烏賊和莖柔魚食物組成差異主要可以通過大眼水蚤科、熒串光魚(Vinciguerria lucetia) 、 朗 明 燈 魚(Diogenichthys laternatus) 、螔 螺科、褶胸魚(Sternoptyx diaphana)和閃光燈籠魚(Myctophum nitidulum)的個數來描述。因此, 魚類是體現兩種頭足類食物組成差異的最重要類別, 總非相似性貢獻率達33%。
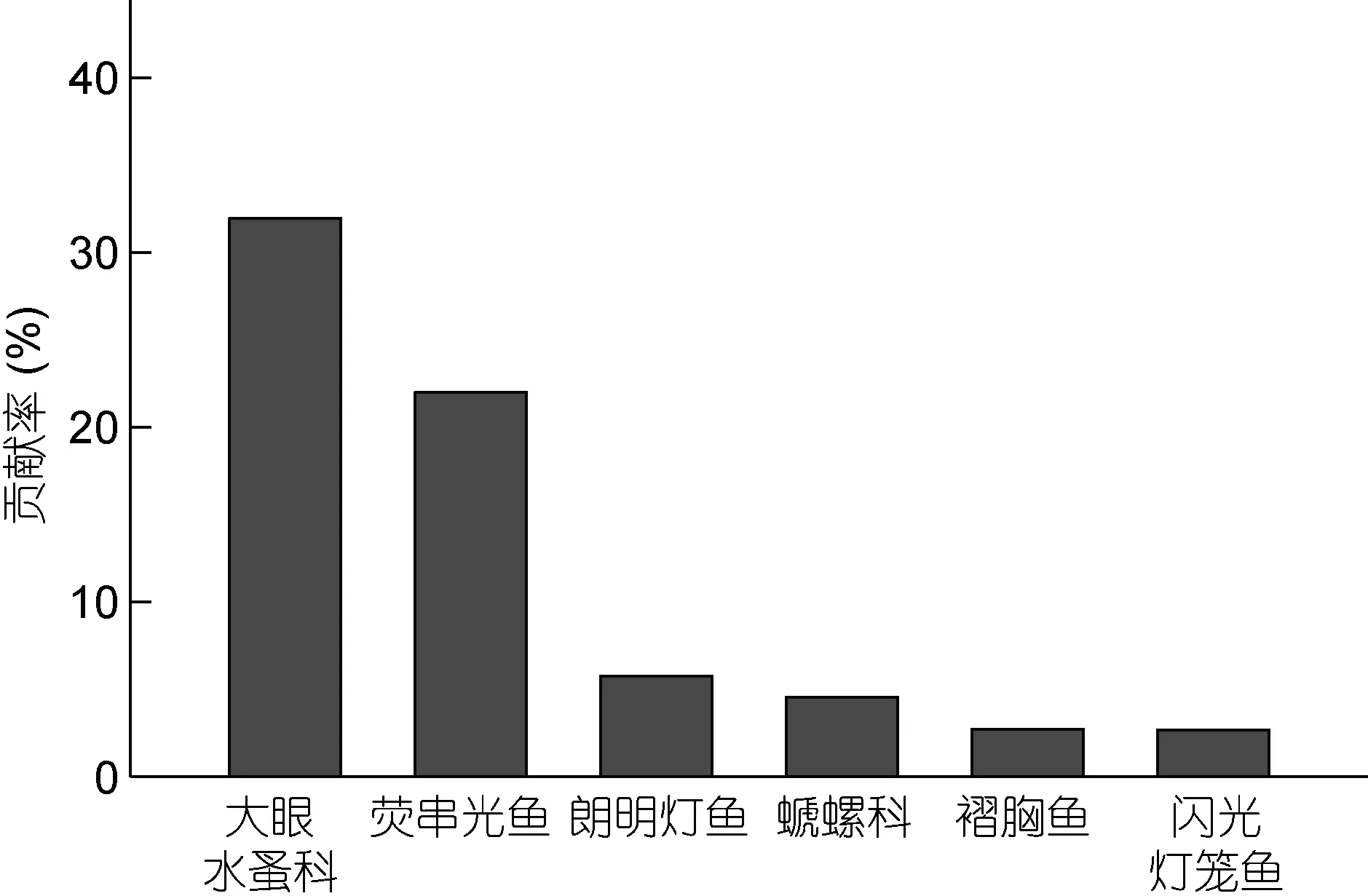
圖3 鳶烏賊和莖柔魚食物組成SIMPER 分析Fig.3 SIMPER analysis of S. oualaniensis and D. gigas food composition
3 討論
3.1 DNA 條形碼的應用
由表4 形態學不可辨認部分與DNA 分子鑒定結果可知, 分子鑒定通過提取胃含物中不可辨認組分,經 COI 序列擴增測序, 共鑒定出餌料生物 33 種,66.67%的序列可以確定到種。其中閃光燈籠魚、金焰燈籠魚、熒串光魚、莖柔魚、鳶烏賊和爪烏賊等6 個物種通過兩種鑒定方法均能準確鑒定, 但總體上兩種方法得到的鑒定結果重疊較少, 糾其原因可能是因為形態學鑒定主要通過胃含物中較難消化的硬組織進行物種識別, DNA 條形碼則通過提取胃含物中未能辨認的食物糜、未知肌肉組織等的DNA 片段進行鑒定。由于頭足類會選擇性攝食獵物的身體部位,很多餌料生物的頭部、骨骼等會被丟棄, 導致胃含物中缺少硬組織作為鑒定依據(Braleyet al, 2010)。此外,頭足類極高的消化率也會使胃含物中殘留的餌料生物消化程度過高, 無法辨別。因此, 造成了兩種鑒定結果間的差異。
總的來說, 形態學鑒定對胃內留存時間較長的硬組織有較好的鑒定率, 但大多數鑒別的生物分類階元較低不能準確鑒定至種, 且頭足類的胃含物細碎存在大量不可辨認食物組分, 形態學觀測無法發揮作用(Symondson, 2002)。采用DNA 條形碼可對形態學鑒定無法識別的較小體積生物組織、未消化的食物糜和表觀相似度較高的物種進行鑒定, 且大部分能鑒定至種, 避開了傳統胃含物分析對形態完整及特定硬組織的嚴格要求。因此, 在小型甲殼動物、軟體動物和高度消化的餌料生物中更具優勢(席曉晴等,2015)。但DNA 條形碼技術也存在一定的局限性, 一方面是引物的選擇, 需要根據不同的目的基因設計不同的特異性引物才能精準地擴增出目的片段, 或者采用多種引物結合使用才能獲取更加精確的結果。另一方面, DNA 條形碼只能達到定性鑒定的目的, 而缺少定量分析數據, 劉夢娜等(2020)基于DNA 條形碼研究中國槍烏賊和鳶烏賊的食物種類組成中, 成功鑒定出中國槍烏賊餌料物種13 種, 鳶烏賊餌料物種8 種, 但未能進行定量分析。因此, DNA 條形碼并不能取代傳統的分類學, 而是作為鑒定未知生物種類的輔助工具, 相比于僅采用單一種的鑒定方法, 兩種方法結合能夠獲得更多樣化的物種信息, 使得食性分析結果較為完整。此外, 在樣品有限的情況下兩種方法結合也可以反映更多的種類信息。
3.2 食性分析
本研究發現東太平洋赤道海域鳶烏賊和莖柔魚均以魚類、頭足類和甲殼類為主要餌料生物。根據各餌料重要性指標判斷, 魚類和頭足類為鳶烏賊的優勢餌料生物, 其中燈籠魚科等小型深海魚類出現頻率最高,F%達66.20%, 而頭足類在鳶烏賊胃含物中比重最大,W%達59.24%。這與Shchetinnikov(1992)對相同海域鳶烏賊胃含物的分析結果一致, 研究發現燈籠魚(燈籠魚屬、標燈魚屬和壯燈魚屬)是該海域鳶烏賊的主要餌料生物。此外, 龔玉艷等(2016)等對我國南海鳶烏賊胃含物的研究也發現, 魚類(主要為燈籠魚科)、頭足類和甲殼動物為鳶烏賊優勢餌料生物。莖柔魚同樣以魚類和頭足類為優勢餌料生物, 但胃含物中甲殼類的F%顯著高于鳶烏賊, 達到65.75%,而鳶烏賊胃含物中甲殼類F%僅33.80%。此外, 莖柔魚餌料生物中出現頻率最高的是熒串光魚(67.12%),占比最重的是頭足類(61.24%), 這與已有研究結果基本一致。Markaida(2006)對加利福尼亞灣莖柔魚胃含物研究表明, 其主要以熒串光魚、巴拿馬底燈魚(Benthosema panamense)和墨西哥尾燈魚等中上層魚類為食。熒串光魚是東太平洋溫熱帶水域最豐富的中上層魚類之一(Ahlstrom, 1968), 垂直遷移是此類魚種最主要的行為特征之一(Franket al, 2002), 并影響著魚類及頭足類(主要是莖柔魚)的生活史及攝食行為(Benoit-Birdet al, 2002; Bertrandet al, 2002)。Rosas-Luis 等(2011)結合胃含物分析和聲學定位觀測到東太平洋海域熒串光魚的晝夜垂直移動與莖柔魚的洄游密切相關, 兩者均在夜晚洄游至海表面, 而白天則遷移至120—300 m 的深海水域, 分布范圍高度重合, 這是熒串光魚在莖柔魚胃含物中占優勢地位的主導因素。綜上所述, 同一海域的鳶烏賊和莖柔魚雖都以魚類、頭足類和甲殼類為餌料生物, 但存在食性選擇差異。可見, 燈籠魚和頭足類是鳶烏賊的優勢餌料生物, 而莖柔魚除頭足類和魚類外, 食物組成中甲殼類的出現頻率也較高, 兩者的攝食選擇具有顯著性差異。對各餌料重要性指標進一步分析顯示。鳶烏賊和莖柔魚胃含物中頭足類和魚類的N%存在顯著差異。因此, 鳶烏賊和莖柔魚食性差異主要體現在魚類的組成差異。SIMPER 分析也表明, 大眼水蚤科、熒串光魚、朗明燈魚、螔 螺科、褶胸魚和閃光燈籠魚等6 種餌料生物是導致鳶烏賊和莖柔魚食性差異的主要種類。但有研究表明(Eduardoet al, 2006), 大眼水蚤等橈足類是熒串光魚的主要餌料生物, 而熒串光魚作為優勢餌料生物被莖柔魚攝食后, 會導致莖柔魚胃含物中大眼水蚤的F%和N%過高; 此外, 在實驗過程中觀察到莖柔魚胃含物中的大眼水蚤多包含于一胃囊狀組織中, 所以推測大眼水蚤等橈足類并不是莖柔魚和鳶烏賊主動攝食的餌料, 而是由魚類帶來的間接性餌料。因此本研究可能會高估大眼水蚤等橈足類在莖柔魚餌料生物中的重要性。該結果有待引入新的判別方法或輔助方法, 以準確區分莖柔魚主動攝食的餌料和餌料生物帶來的間接性餌料。
此外, 本研究發現鳶烏賊和莖柔魚均存在一定程度的同類相食。這在其他頭足類研究中已得到廣泛驗證。頭足類是貪婪的機會主義捕食者(Markaidaet al, 2003), 由于快速生長和高代謝需求, 在種群密度過高或食物缺乏的情況下會發生同類相食的現象(Ibá?ezet al, 2010; Brunoet al, 2021)。已有研究發現,漁業活動會加劇頭足類同類相食現象的發生。Ibarra-Garcia 等(2014)對采集自加利福尼亞灣漁區和非漁區的莖柔魚胃含物比較發現, 漁區內樣本胃含物中莖柔魚的總重量和質量百分比均顯著高于非漁區, 認為是因捕撈作業聚集了大量易被捕食的同類個體, 導致自食現象的加劇。本研究中鳶烏賊和莖柔魚均在捕撈作業時采集, 捕撈活動可能會導致較高程度的同類相食現象發生。
4 結論
綜上所述, 本研究通過結合胃含物分析和DNA條形碼技術, 對東太平洋赤道海域鳶烏賊和莖柔魚的食性進行了比較研究, 發現兩者均以魚類、頭足類和甲殼類為主要餌料生物, 但它們在食物資源利用方面各有偏好, 在Shannon 多樣性指數和食物組成上均存在顯著差異, SIMPER 分析進一步表明, 魚類是導致鳶烏賊和莖柔魚食物組成差異的重要原因, 兩者的攝食選擇具有顯著性差異。該結果驗證了前期對兩種頭足類肌肉穩定同位素和特征脂肪酸的分析結果(李云凱等, 2020), 兩者雖生物學特性相似且棲息在同一海域, 但兩種頭足類會通過食性分化降低種間競爭, 從而達到穩定共存。本研究中, 主要通過多種餌料重要性指標反映了鳶烏賊和莖柔魚食物組成差異, 從食物分化的角度探討了兩者的攝食策略差異。此外, 攝食時間、水層等可能也是影響攝食策略差異的因素, 因此, 在未來的研究中, 應該全面考慮鳶烏賊和莖柔魚在不同采樣年份和水層等方面是否具有差異, 從而得到更加全面的食性分析結果。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
音樂探索(2022年2期)2022-05-30 21:01:37
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
