滸苔光照和鹽度脅迫響應基因UpMYB44 的克隆與表達分析*
2021-10-11 09:51:50楊娟娟沈頌東
海洋與湖沼 2021年5期
何 淵 楊娟娟 沈頌東
(蘇州大學基礎醫學與生物科學學院 蘇州 215000)
非生物脅迫嚴重影響了植物的生長和發育, 面對不利的環境, 植物可以通過引發一系列的生理生化反應去適應這種不良影響(Huanget al, 2012)。MYB轉錄因子可以作為植物響應非生物脅迫的重要調控因子,已被廣泛研究(Ambawatet al, 2013),MYB(V-myb avian myeloblastosis viral oncogene homolog)轉錄因子是植物中最大的轉錄因子家族之一, 存在于真核生物中, 廣泛參與植物細胞分化、信號轉導和次級代謝調控等多種生命活動, 特別在植物的轉錄調節中發揮著重要的作用(Mikamiet al, 2013)。根據MYB 蛋白含有的不同數量的結構域,MYB基因可以分為1R、R2R3、3R、4R 這四種類型(杜靜靜等, 2019)。尤其是R2R3-MYB類轉錄因子, 該類轉錄因子含有2 個MYB結構域, 參與調節植物在面對環境脅迫時的初級和次級代謝活動(Chenet al, 2019)。植物中最早發現MYB基因的是玉米體內的C-MYB, 該基因參與玉米的花青素合成過程(Maroccoet al, 1989), 在后續的研究中又陸續在多種植物中發現了MYB基因, 在棉花中發現了219 個基因(Baldoniet al, 2015), 在水稻中鑒定出183個MYB基因(Chenet al, 2006),GmMYB010參與了大豆抗蟲的過程(陸撿花等, 2017)。
在環境脅迫響應方面, 對MYB基因也進行了相應的研究。比如在剛毛檉柳(Tamarix hispida)中對MYB基因家族在環境脅迫下的表達進行了差異分析(Zhanget al, 2018),TaMYB參與了擬南芥(Arabidopsis thaliana)對干旱環境脅迫的響應過程(Zhaoet al,2018), 植物中大部分的R2R3-MYB能夠響應植物的鹽脅迫從而提高其耐鹽能力(張遵強等, 1998)。在諸多的環境因素中, 光照和鹽度是影響藻類生命活動的重要環境因子(Heet al, 2017)。在光照方面, 高光照條件下會影響杜氏鹽藻和雨生紅球藻類胡蘿卜素合成途徑中關鍵基因的表達水平(Picket al, 2019), 光照強度同樣對滸苔的生長狀態有關鍵的影響(Liet al, 2012); 在鹽度方面, 高鹽度的培養會促進杜氏鹽藻類胡蘿卜素的合成(Shanget al, 2018), 鹽度同樣會影響滸苔的氮元素富集從而影響其增殖(Zhenget al, 2019)。
滸苔(Ulva prolifera)是一種常見的大型海洋綠藻,具有單層細胞圍繞成的管狀結構, 廣泛分布于低潮區的灘涂、礁石和底泥之中(Fanet al, 2014)。滸苔具有細絲狀易于漂浮的外觀, 多樣的繁殖方式以及能夠抵御多種環境因子脅迫等生物學特征可以幫助滸苔承受多種環境壓力, 例如高鹽、干旱和強光照條件,且可以在較廣的溫度范圍內存活, 是一種生命力很強的海藻(Xiaoet al, 2016)。從2007 年起, 我國的黃海海域每年都會暴發以滸苔為主的“綠潮”災害, 據統計每年“綠潮”災害所引發的生物量達到了數百萬噸(Liuet al, 2015), 給沿海地區造成了巨大的經濟損失和嚴重社會影響(Zhanget al, 2017; 王宗靈等, 2018;Zhaoet al, 2019a)。相對于MYB家族在高等植物中的生長發育、生理代謝、生物脅迫與非生物脅迫功能研究的大量研究成果, 人們對MYB家族轉錄因子在滸苔中的結構和功能研究較少。滸苔能夠引發“綠潮”災害和滸苔具有抵御夏季海面的極端環境的能力密切相關, 研究表明MYB家族的轉錄因子具有應答外界環境脅迫的能力(Duboset al, 2010), 因此我們推測MYB類轉錄因子在滸苔夏季抵御海面嚴酷的環境脅迫大量增殖的過程中起到了一定的作用。
本研究依據前期已經完成的滸苔光照和鹽度脅迫下的轉錄組測序結果, 篩選出具有表達水平顯著變化的MYB類基因MYB44, 將其命名為UpMYB44,進一步對其進行序列擴增、生物信息學分析、實時熒光定量PCR 分析等實驗初步探究了該基因的表達模式和分子功能, 證明其在非生物脅迫例如鹽度和光照的響應過程中發揮了重要的調控作用, 為今后深入研究UpMYB44 基因的功能奠定了堅實的基礎。
1 材料與方法
1.1 實驗材料
于2020 年7 月在山東省青島市市南區棧橋潮間帶(37.46°N, 121.71°E)采集滸苔配子體樣品, 放入容器中帶回蘇州大學藻類實驗室。用無菌水清洗藻體,除去表面的泥沙和其他雜物, 放置于光照培養箱培養。培養條件為f/2 培養基, 溫度為20 °C, 光照強度為 90 μmol photons/(m2·s), 鹽度為 24, 光周期為:L:D=12 h : 12 h。每3 d 換一次過濾海水, 預培養7 d。
1.2 光照和鹽度脅迫處理
用三種不同光照培養滸苔 5 d, 18 μmol photons/(m2·s) 設 置 為 低 光 照 組(l), 90 μmol photons/(m2·s) 設 置 為 中 光 照 組(m), 216 μmol photons/(m2·s)設置為高光照組(h); 用三種不同鹽度海水培養滸苔5 d, 低鹽度組(L)設置為12, 中鹽度組(M)設置為24, 高鹽度組(H)設置為40, 培養密度為20 g/L, 溫度為20 °C, 光周期為: L:D=12 h : 12 h, 1 d換一次過濾海水, 以上處理的每種樣品均設置3 個生物學重復。
1.3 RNA 提取和cDNA 合成
參照(Heet al, 2018)的方法使用Takara 的總RNA提取試劑盒對三種不同光照和鹽度處理的滸苔進行總RNA 的提取, 同時通過Thermo Fisher NanoDrop 2000 和瓊脂糖凝膠電泳對所提取的RNA 的濃度和純度進行檢測, 使用Takara Reverse Transcription kit 將2 μg 檢測合格的RNA 合成為cDNA 第1 鏈。
1.4 UpMYB44 基因的cDNA 全長克隆
根據實驗室獲得的滸苔光照和鹽度脅迫下的轉錄組數據(GenBank: SRX4552126, SRX5502767)設計一對特異性引物UpMYB44F/R,UpMYB44F: ATGAC TTCCGAGGGGATGGAG,UpMYB44R: GACGAGCT GCTCGCGGTTAAA。以之前獲得的cDNA 為模板,使用 2×HieffTMPCR Master Mix (Yeasen) 擴增UpMYB44的ORF 全長序列, 50 μL PCR 反應體系如下:2×HieffTMPCR Master Mix 25 μL, 上游引物和下游引物(10 μmol/L)各2 μL, cDNA 模板2 μL, 加19 μL 的ddH2O 補至50 μL。PCR 反應程序如下: 94 °C 預變性5 min, 94 °C 變性30 s, 55 °C 退火30 s, 72 °C 延伸1 min, 30 個循環, 72 °C 延伸10 min。取上述PCR 產物 10 μL 進行 1%瓊脂糖凝膠電泳, 使用 Gel Extraction Kit (康為世紀)切取符合目的基因片段大小的條帶進行回收。使用 Hieff Clone?Zero TOPOBlunt Cloning Kit (Yeasen)進行目的片段平末端擴增,將構建好的質粒接種于LB 固體培養基上過夜培養,第二天挑取陽性克隆測序, 獲得UpMYB44基因ORF區全長序列。
1.5 UpMYB44 生物信息學分析
使用DNAMAN 軟件對UpMYB44 進行氨基酸保守序列分析; 利用 Expays (https://web.Expasy.Org/protparam/)在線軟件分析編碼蛋白的大小、氨基酸數量和等電點; 使用 NCBI 的 CDD 數據庫
(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)
對蛋白結構域進行預測; 利用 ProtScale 工具(http://web.Expasy.org/protscale/)對 UpMYB44 蛋白進行親水性/疏水性分析; 采用 ExPaSy-SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)在線軟件預測蛋白質的二級結構;利用 SWISS-MODEL (https://swissmodel.expasy.org/interactive) 對UpMYB44 蛋白3D 結構進行分析。
1.6 表達載體的構建及轉化
參照Al-Muhanna 等(2018)的方法采用T4 連接酶將UpMYB44 目的片段與pET-32a 載體(反應體系包含0.1 pmol 目的片段, 0.03 pmol 載體片段, 3 U T4 DNA 連接酶, 1×T4 DNA 連接酶緩沖液)在16 °C 反應 12 h。采用熱激法將連接產物轉入大腸桿菌Trans-T1 感受態細胞。用雙酶切進行(Nde I, Xba I)鑒定, 將陽性質粒送去蘇州金唯智公司進行測序鑒定。
1.7 UpMYB44 的蛋白表達
參照 B a n e y x(1 9 9 9)的方法將 p E T-3 2 a+UpMYB44(+)1 μL 加入100 μL BL21(DE3)感受態細菌中, 置冰上20 min, 37 °C, 220 r/min 振搖1 h, 離心后全部涂布于含50 μg/mL Amp 的LB 平板, 37 °C 倒置培養過夜。挑取轉化平板上的單克隆接種在含50 μg/mL Amp 的3 mL LB 液體培養基的培養管中,在220 r/min, 37 °C, 4 h 搖床里進行培養, 當分光光度計測量值顯示其OD600(600 nm處的吸光值)≈ 0.6—0.8時, 滴入濃度為1.0 mmol/L IPTG, 37 °C 220 r/min 經過振搖4 h 誘導蛋白表達后取2 mL, 然后14 000 r/min離心, 棄去上清液, 采用PBS 來重懸沉淀, 加2×十二烷基硫酸鈉(sodium dodecyl sulfate loading buffer,SDS)上樣緩沖液混合均勻之后, 沸水煮沸10 min。然后進行十二烷基硫酸鈉聚丙烯酰胺凝膠電泳 (sodium dodecyl sulfate-polyacrylamide gel electrophoresis, SDSPAGE)超聲破碎后收集上清, 用0.45 μm 濾膜過濾,用3 倍體積的平衡液[ 25 mmol /L Tris( pH 值 8.0),300 mmol/L NaCl, 10 mmol/L 咪唑] 平衡 His 純化基質, 將過濾后的上清與His 純化基質于50 mL 離心管中4 °C 混勻1 h。用洗滌緩沖液[25 mmol /L Tris ( pH值 8.0), 300 mmol/L NaCl, 30 mmol/L 咪唑] 沖洗4 次,用洗脫緩沖液[25 mmol/L Tris ( pH 值 8.0),300 mmol/L NaCl, 250 mmol/L 咪唑]洗脫3 次, SDSPAGE 電泳檢測融合蛋白純化效果。
1.8 UpMYB44 亞細胞定位
參照Xiong 等(2019)的方法設計亞細胞定位引物YFP-UpMYB44F/R, YFP-UpMYB44F: AAGAGA CAGGATCCGAATTCATGACTTCCGAGGGGATGGA G, YFP-UpMYB44R: ACCTCCGACCGGTGCACTA GTGACGAGCTGCTCGCGGTTAAA。將酶切位點BamH I 和Sac I 加入目的基因ORF 兩端, 將測序成功的陽性質粒T-blunt-UpMYB44 為模板進行擴增和切膠回收。使用BamH I 和Sac I 對質粒pC131-YFP 進行雙酶切, 產物純化。將目的基因片段和載體pC131-YFP 進行連接轉入大腸桿菌DH5α 感受態細胞,獲得重組質粒 pC131-UpMYB44-YFP。將空載質粒pC131-YFP 和重組質粒pC131-UpMYB44-YFP 分別轉入農桿菌GV3101 菌株中, 將陽性克隆放入含有50 μg/mL 卡那霉素和35 μg/mL 利福平的LB 培養基中進行培養, 放置于28 °C 搖床中以250 r/min 速度培養過夜, 收集并重懸菌體, 當菌體濃度的OD600值在0.5—1.0 之間, 靜置3 h 后注射入煙草葉片中, 注射后的煙草放置于28 °C 培養箱中培養2 d。用激光共聚焦顯微鏡觀察侵染的煙草葉片, 進行UpMYB44蛋白亞細胞觀察。
1.9 實時熒光定量PCR
參照He 等(2019)的方法設計實時熒光定量PCR并以不同光照和鹽度處理的滸苔cDNA 為模板進行實時熒光定量PCR 實驗, 熒光定量PCR 采用 Hieff?qRT-PCR SYBR Green Master Mix (Yeasen)試劑, 使用ASA-4800 Real-Time PCR 儀器(百源基因)進行反應, 以18S rDNA為內參基因, 具體引物序列如表1所示, 上述實驗重復3 次。結果分析采用 2-ΔΔCT法進行相對定量, 采用軟件Origin 8.0 對表達結果進行單因素方差統計(ANOVA)。數據使用 SPSS 21.0 進行統計分析,P< 0.05 設定為顯著性差異,P< 0.01 設定為極顯著性差異。

表1 實時熒光定量PCR 所用引物Tab.1 Primers used for quantitative real-time PCR
1.10 UpMYB44 和UpCPP5 互作驗證
參照Cai 等(2016)的方法制備Y2H Gold 酵母感受態細胞, 將UpCPP5 作為誘餌構建PGBKT7-CPP5重組質粒進行毒性檢測和自激活檢測: 于100 °C 沸水煮carrier DNA 5 min, 立即置于冰上2 min, 如此重復一次, 取1.5 mL 無菌EP 管, 配制轉化體系, 50 μL感受態細胞+5 μL carrier DNA+100 ηg 誘餌質粒+100 ηg 獵物空載, 加入500 μL PEG/LiAc (8 mL 50%PEG+1 mL LiAc+1 mL TE), 30 °C 水浴30 min, 加入20 μL DMSO, 42 °C 水浴15 min, 800 ×g離心1 min,去上清, 加入800 μL YPDA 重懸, 30 °C, 150 r/min,振蕩培養1.5 h, 800 ×g離心5 min, 棄上清, 加入1 mL 0.9% NaCl 重懸, 取150 μL 涂平板, 30 °C 倒置培養 4 d, 觀察菌落直徑及顏色; 構建 PGADT7-MYB44 重組質粒作為獵物進行誘餌和獵物的共轉化,依次從自激活、共轉化實驗組和共轉化陰陽性對照組涂布的平板 DDO 上挑單菌落, 分別接種于 5 mL DDO 液體培養基, 30 °C 250 r/min 培養16 h, 取300 μL 菌液于50 mL DDO 中, 30 °C 250 r/min 培養8—12 h 至OD600值為0.4—0.6; 各取10 μL 菌液點種到平板DDO、TDO/3AT (15 mmol/L)、QDO 上, 30 °C培養箱培養3 d, 觀察菌斑生長情況, 若涂布在平板DDO 上長菌則共轉化成功, 在平板 TDO/3AT(15 mmol/L)上長菌而在QDO 上不長菌為陽性結果,在平板TDO/3AT (15mmol/L)上不長菌為陰性結果。
2 結果
2.1 UpMYB44 ORF 區全長序列的克隆與序列分析
利用RT-PCR 以滸苔cDNA 為模板, PCR 擴增相應片段, 測序得到UpMYB44的ORF 區全長序列, 序列全長為1 437 bp, 共編碼478 個氨基酸。序列上傳至Genbank 數據庫, 登錄號為MW174238。
2.2 UpMYB44 生物信息學分析結果
通過 NCBI 與其他物種的 MYB 蛋白進行BLASTn 比對, 結果顯示UpMYB44 蛋白與另一種綠藻小球藻的MYB44 蛋白有較高的同源性, 氨基酸的一致性為50.00%, 與高等植物番木瓜、荷花、木槿、牽牛花、柿樹和向日葵的氨基酸相似率也在45.00%—50.00%之間, 用DNAMAN 將UpMYB44 氨基酸序列與上述物種進行比對, 發現了較為保守的序列區域(圖1), 推測與這些MYB 類轉錄因子有相似的生物學功能。利用Expays 在線網站預測結果顯示UpMYB44 蛋白含有 478 個氨基酸, 分子量為52.49 kDa, 等電點為6.15; 利用CDD 數據庫分析顯示 UpMYB44 含有兩個 SANT 結構域, 是典型的R2R3-MYB(圖2); 利用ProtScale 工具對UpMYB44蛋白進行親水性/疏水性分析, 結果顯示UpMYB44為親水性蛋白; 在SOPMA 在線網站預測蛋白二級結構, UpMYB44 蛋白二級結構中含有42.68%的α-螺旋結構、8.16%的延伸連結構、9.62%的β-轉角結構、39.54%的無規則卷曲結構(圖3)。利用Swiss-model網站對UpMYB44 蛋白進行3D 結構預測, 發現與其蛋白二級結構結果類似, 具有較多的α-螺旋結構和無規則卷曲結構(圖4)。
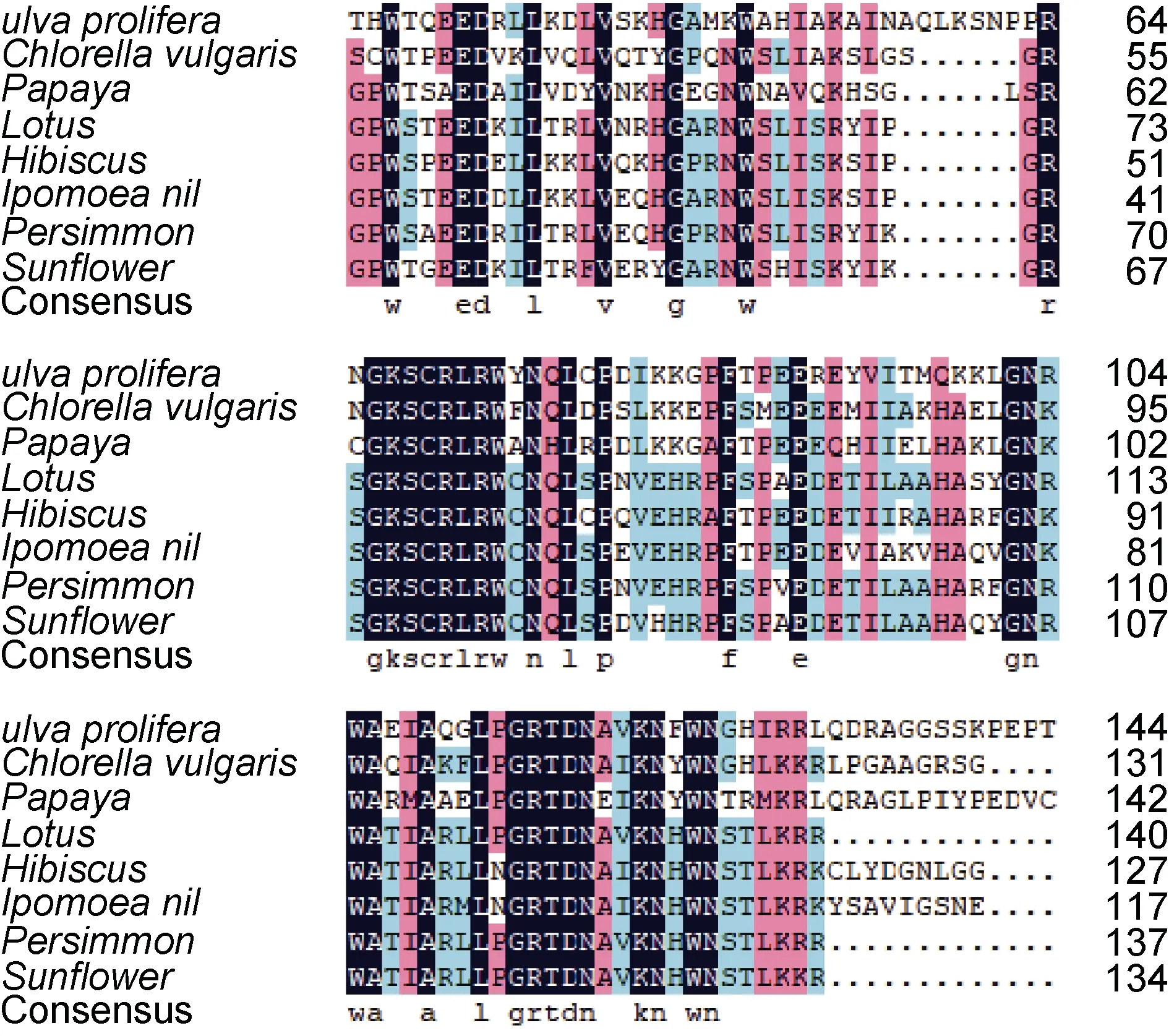
圖1 UpMYB44 與其他物種MYB 類轉錄因子氨基酸序列的多重比對Fig.1 Multiple alignment of the amino acid sequence of UpMYB44 and MYB transcription factors from other species

圖2 UpMYB44 蛋白保守結構域預測Fig.2 Prediction of conservative domain of UpMYB44 protein

圖3 UpMYB44 二級結構預測Fig.3 Prediction of secondary structure of UpMYB44
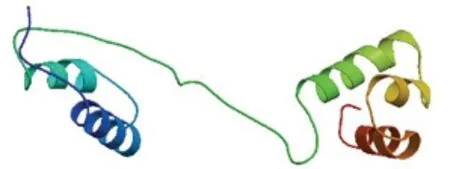
圖4 UpMYB44 蛋白三維結構預測模型Fig.4 Predicted 3D structure model of UpMYB44 protein
2.3 pET-32a+UpMYB44 融合蛋白的可溶性分析和純化
結果表明, UpMYB44 融合蛋白主要以可溶性蛋白的形式存在于上清中, 但是在包涵體沉淀中也有一定量的表達(圖5)。選擇上清中可溶性蛋白進行后續蛋白純化實驗, 通過4 次洗滌和3 次洗脫, 獲得了大小約為65 kD (含標簽)的純度較好的UpMYB44 融合蛋白(圖6)。
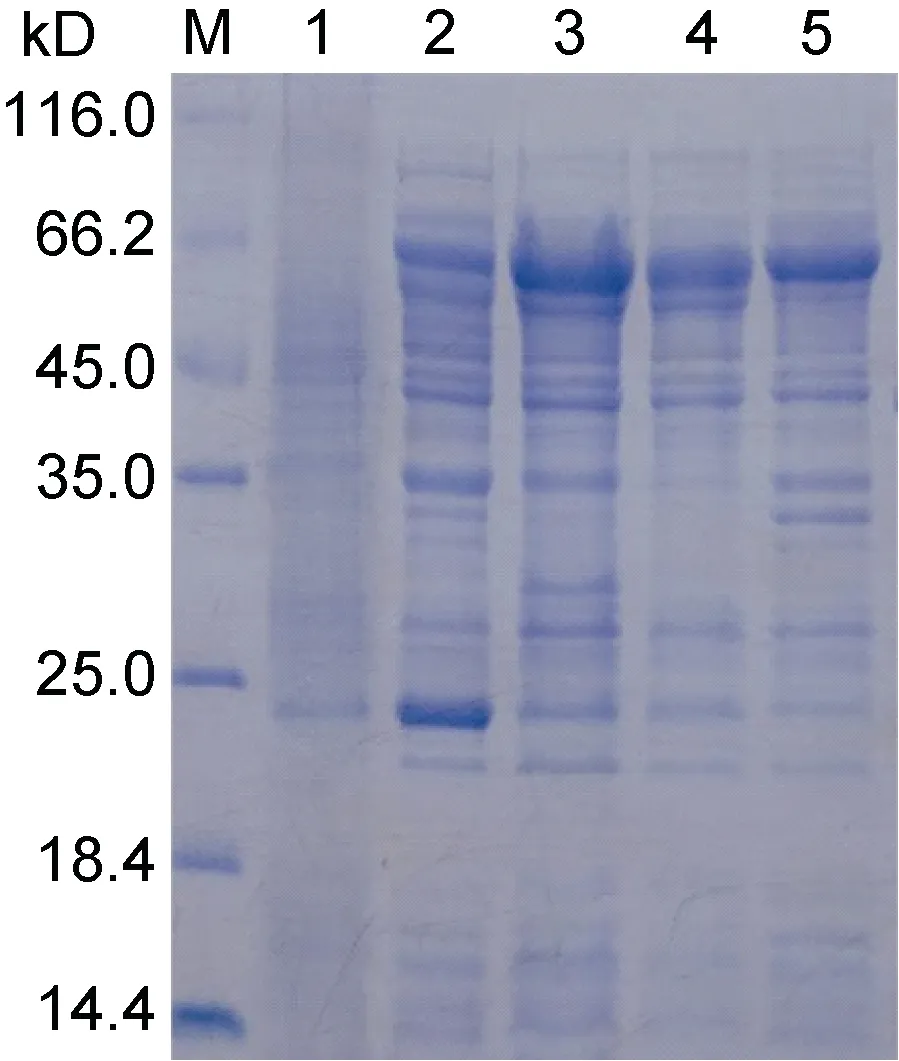
圖5 UpMYB44 蛋白表達鑒定SDS-PAGE 分析Fig.5 Identification of UpMYB44 protein expression by SDS-PAGE analysis
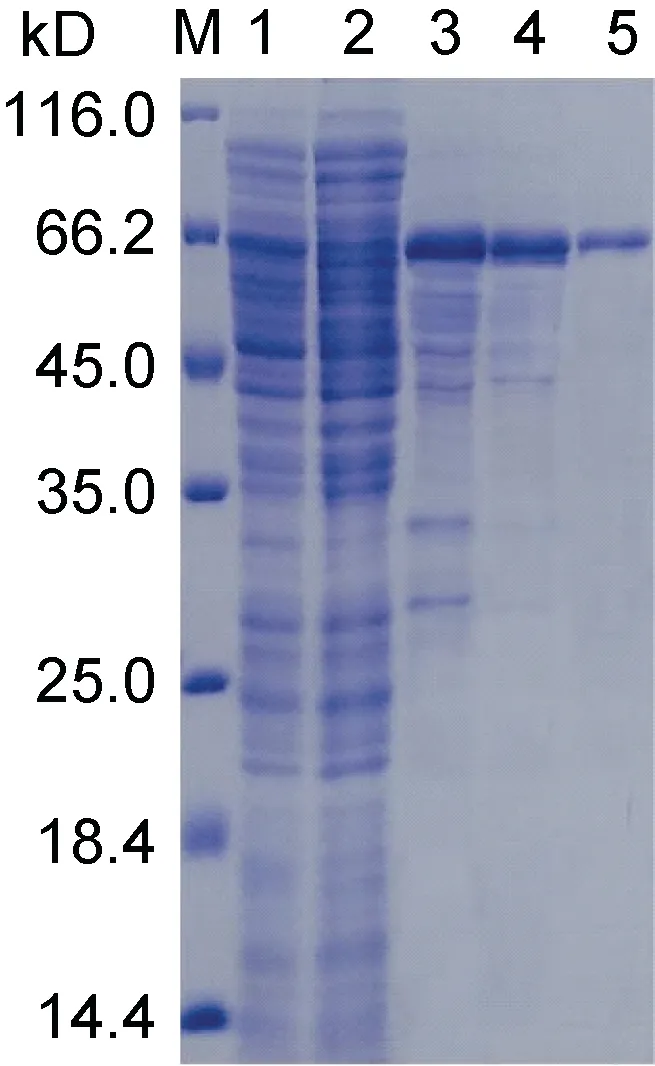
圖6 UpMYB44 蛋白純化SDS-PAGE 分析Fig.6 Purification of UpMYB44 protein by SDS-PAGE analysis
2.4 UpMYB44 蛋白的定位
在對照組空載pC131-YFP 中, 黃色熒光在細胞膜、細胞核和細胞質中都有分布, 而融合表達載體pC131-UpMYB44-YFP 的細胞中, 黃色熒光僅在細胞核中被發現, 而在細胞膜和細胞質中都沒有熒光發現, 并且黃色熒光和核定位的marker 蛋白的紅色熒光發生重合。結果顯示UpMYB44 蛋白僅定位在細胞核上, 這一點符合MYB 類轉錄因子的特點(圖7)。

圖7 UpMYB44 蛋白的亞細胞定位Fig.7 Subcellular localization of the UpMYB44 protein
2.5 UpMYB44 在不同光照和鹽度處理下的表達模式分析
UpMYB44在三種不同光照和鹽度處理下的表達量發生了顯著變化, 在光照組中, 18 μmol photons/(m2·s)處理的低光組中UpMYB44的表達量最高, 隨著光照強度的增加,UpMYB44的表達量逐漸降低( 圖 8),UpMYB44的 表 達 量 在 216 μmol photons/(m2·s)處理的高光組中最低; 在鹽度組中, 隨著鹽度的增加,UpMYB44的表達量逐漸升高,UpMYB44的表達量在鹽度40 處理的高鹽組中最高,在鹽度12 處理的低鹽組中最低(圖9)。
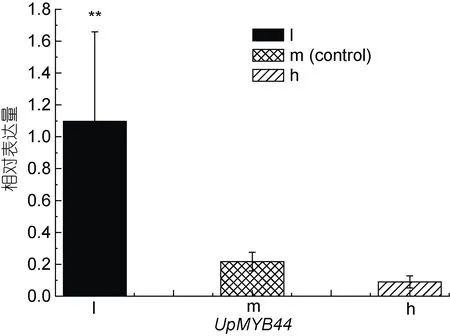
圖8 UpMYB44 在三種光照強度條件下的表達分析Fig.8 Expression analysis of UpMYB44 under three lights stresses

圖9 UpMYB44 在三種鹽度強度條件下的表達分析Fig.9 Expression analysis of UpMYB44 under three salinities stresses
2.6 UpMYB44 和UpCPP5 蛋白存在互作關系
MYB 類轉錄因子通常需要與其他蛋白互作進行發揮其功能。利用STRING 在線網站預測UpMYB44的互作蛋白, 根據生物信息學預測結果發現UpMYB44 和另一種調控植物生長發育和響應非生物脅迫中起重要作用的轉錄因子UpCPP 存在互作關系,以缺失蛋白UpCPP5 為誘餌, 通過酵母雙雜交點對點驗證, 結果表明重組誘餌質粒PGBKT7-CPP5 對酵母細胞無毒性且3-氨基-1,2,4 三唑(3AT, 15mmol/L)能抑制激活Y2H Gold 酵母報告基因, 酵母雙雜交的結果顯示, 將獵物質粒 PGADT7-MYB44 與誘餌質粒PGBKT7-CPP5 共轉化酵母細胞: 對照結果符合預期,說明該系統可用于雙雜交驗證; 實驗組涂布DDO 平板能生長說明誘餌和獵物共轉化成功, 涂布TDO/3AT (15mmol/L)板能生長, QDO 平板不長, 說明激活報告基因HIS3 的表達, 說明UpMYB44 蛋白和UpCPP5 蛋白之間存在相互作用(圖10)。

圖10 UpMYB44 與UpCPP5 互作蛋白驗證Fig.10 Verification of UpMYB44 interacting with UpCPP5
3 討論
研究發現在植物中MYB類轉錄因子參與了多種生物和非生物脅迫的響應過程(Leaet al, 2007)。超表達玉米中的ZmMYB3R可以增加其對干旱和鹽度的耐受性(Yinet al, 2017), 過量表達煙草的NtMYB15可以促進其適應低溫脅迫(Wuet al, 2019), 在模式生物擬南芥中超表達AtMYB74同樣具有增強其鹽耐受性的功能(Xuet al, 2015), 而且MYB 類轉錄因子常常需要與其他蛋白互作來發揮功能(Kasparet al, 1999)。
本研究以受關注度較高的引發“綠潮”災害的滸苔為研究對象, 初步分析了MYB類家族成員UpMYB44的生物學功能。結果表明UpMYB44有兩個MYB結構域, 屬于典型的R2R3-MYB 轉錄因子, 研究表明該類轉錄因子參與了棉花和月季對高鹽脅迫的響應(Zhaoet al, 2019b, 包穎等, 2020)。鹽度和光照是影響藻類正常生長和生理生化指標的重要的兩個環境因子(Araiet al, 1991), 特別是對滸苔而言, 鹽度和光照的改變會引起其氧化應激反應(Luoet al,2011), 以及影響其體內的總類胡蘿卜素含量(Heet al,2020)。在本研究中,UpMYB44在高鹽條件下的表達量發生了顯著提高, 說明該轉錄因子參與了滸苔的對高鹽脅迫的響應過程, 可能對提高滸苔適應惡劣的外界環境的能力有一定幫助, 最終有利于其快速增殖。R2R3-MYB轉錄因子也會對光照的改變作出響應, 在早期的研究中發現紅色紫蘇中存在一種光誘導的Myb-p1基因, 其在花青素的形成過程中起到了決定性作用(Gonget al, 1999), 在馬鈴薯中發現的StR2R3-MYB1就是一種光響應MYB類轉錄因子, 基于對光處理后基因表達特性分析的結果表明該轉錄因子可能受到光信號和環境脅迫的誘導(秦玉芝等,2015)。在滸苔中UpMYB44的表達量同樣受到了不同光照強度的影響, 且隨著光照的逐漸增強其表達量呈下降趨勢, 這和一般應激反應的表現有差異, 推測夏季滸苔在漂浮狀態下的高光照強度脅迫會影響其正常的生長狀態, 導致部分基因的表達出現下調, 說明過高的光強對其增殖反而不利。UpMYB44基因在滸苔受到光照和鹽度脅迫時會發生響應, 對夏季抵御海面嚴酷的環境脅迫起到一定作用。對MYB類轉錄因子互作模式的研究也已經開展, 比如在蘋果中MdMYB23 蛋白通過結合MdBT2 蛋白來抑制原花色素的積累(Anet al, 2018), 通過鑒定與AtMAPK3P 相互作用的MYB 轉錄因子成員, 增強了我們對擬南芥中MAPK 相互作用蛋白網絡的認識, 也證實了蛋白質與蛋白質的相互作用對于生物過程至關重要(Giriet al, 2014)。
在本研究中, 通過酵母雙雜交系統證明了UpMYB44 蛋白和一種CPP 蛋白具有互作, 通過和Genbank 中的序列進行比對, 我們將其命名為UpCPP5。CPP 蛋白又稱為tesmin/TSO1-like 蛋白, 在植物中廣泛分布, 該家族在植物生長發育和細胞分裂控制中起重要作用(Yanget al, 2008), TSO1 參與了擬南芥花發育的細胞分裂過程(Liuet al, 1997), 如果該基因輕度的突變會影響擬南芥胚珠發育, 而強突變對所有花卉組織都有影響(Andersenet al, 2007),因此, 猜測這兩類轉錄因子的相互作用影響了滸苔的細胞分裂過程, 這個研究方向正好和目前滸苔如何引發“綠潮”災害的研究熱點相互關聯。本實驗的研究材料滸苔是一種能夠快速生長的災害綠藻, 只要外界條件合適, 其生長速率相當驚人(Hiraokaet al,2008), 每年夏季由于其暴發性生長對我國沿海城市造成了巨大的環境污染(Yuet al, 2017; Heet al, 2019),對近海的生態系統也造成了嚴重的破壞(Quet al,2020)。根據之前滸苔基因組的研究, 我們推測滸苔的快速生長和細胞的快速分裂密切相關, 但是滸苔增殖最終重要的是營養增殖, 因此后續可以通過深入研究滸苔中MYB 家族和CPP 家族的互作是否會影響滸苔的生長發育和細胞分裂, 并結合滸苔營養增殖方面的研究為進一步了解環境脅迫調控網絡提供依據, 更重要的是為研究滸苔的快速繁殖機制提供了全新的思路。
4 結論
本研究在本實驗室已有的研究基礎上, 克隆了1個R2R3-MYB轉錄因子基因——UpMYB44的ORF 區全長序列, 其氨基酸序列與GenBank 中已有的小球藻MYB 蛋白具有相似性, 我們對其進行了生物學功能驗證, 在異于滸苔最適環境的光照和鹽度的培養條件下,UpMYB44的表達量會發生顯著變化, 證明其參與了滸苔響應光照和鹽度壓力的過程, 可能在鹽度和光照壓力應答中具有關鍵作用。實驗證明UpMYB44 與CPP 類轉錄因子UpCPP5 存在較強的互作, 推測二者的結合可能參與了滸苔的生長發育和細胞分裂過程, 但其具體的作用機制還需要進一步研究。
