CC類趨化因子配體2對(duì)結(jié)核分枝桿菌感染人單核巨噬細(xì)胞THP-1自噬的影響及分子機(jī)制
2021-09-27 02:32:22李秀萍嚴(yán)曉蕓
臨床誤診誤治 2021年9期
李秀萍,王 玲,王 馨,嚴(yán)曉蕓,董 力
結(jié)核病是一種嚴(yán)重的傳染病,由結(jié)核分枝桿菌感染引起,對(duì)人類健康構(gòu)成了巨大威脅。有流行病學(xué)統(tǒng)計(jì)數(shù)據(jù)顯示,近年來(lái)我國(guó)結(jié)核病病例顯著增加[1]。有研究表明,結(jié)核分枝桿菌是一種細(xì)胞內(nèi)病原體,可寄生于宿主巨噬細(xì)胞,通過(guò)調(diào)控炎癥反應(yīng)及吞噬體酸化、成熟等不同機(jī)制調(diào)節(jié)宿主細(xì)胞死亡[2]。巨噬細(xì)胞能夠殺滅微生物,但結(jié)核分枝桿菌仍然能夠克服巨噬細(xì)胞的殺微生物機(jī)制而存活并復(fù)制[3]。自噬促進(jìn)受損細(xì)胞器和錯(cuò)誤折疊蛋白質(zhì)降解,在抗菌防御機(jī)制中起著關(guān)鍵作用。重要的是,自噬既可以調(diào)節(jié)宿主對(duì)分枝桿菌感染的抗性,又可以控制細(xì)胞的存活[4]。CC類趨化因子配體2(CCL2)是一種調(diào)節(jié)單核細(xì)胞和巨噬細(xì)胞向感染部位遷移和滲透的關(guān)鍵趨化因子,其水平與結(jié)核病嚴(yán)重程度相關(guān)[5]。有研究顯示,CD44能夠促進(jìn)結(jié)核分枝桿菌感染巨噬細(xì)胞中CCL2的表達(dá)[6]。還有研究表明,CCL2參與調(diào)節(jié)角質(zhì)形成細(xì)胞的自噬[7]。然而,現(xiàn)臨床尚未確定CCL2對(duì)結(jié)核分枝桿菌感染巨噬細(xì)胞自噬的影響。因此,本研究探討CCL2對(duì)結(jié)核分枝桿菌感染人單核巨噬細(xì)胞THP-1自噬的影響及分子機(jī)制,以期為結(jié)核病的防治提供實(shí)驗(yàn)基礎(chǔ)。
1 材料與方法
1.1細(xì)胞株和主要試劑 結(jié)核分枝桿菌H37Rv株和人單核巨噬細(xì)胞THP-1(美國(guó)ATCC);RPMI 1640培養(yǎng)基和青-鏈霉素雙抗(美國(guó)HyClone);胎牛血清(美國(guó)Gibco);Lipofectmine 2000轉(zhuǎn)染試劑(美國(guó)Invitrogen);TRIzol試劑(北京索萊寶);PrimeScript RT試劑盒和Green Premix Ex Taq Ⅱ(日本TaKaRa);CCL2 siRNA和siRNA control(廣州銳博生物);CCL2、LC3-Ⅰ、LC3-Ⅱ、p62、p65、IKK-α和IκB-α單克隆抗體及辣根過(guò)氧化物酶標(biāo)記的二抗(美國(guó)CST);RIPA裂解液(上海碧云天);SDS-PAGE凝膠制備試劑盒、BCA蛋白濃度測(cè)定試劑盒和ECL化學(xué)發(fā)光試劑盒(賽默飛世爾)。
1.2細(xì)胞培養(yǎng) THP-1細(xì)胞復(fù)蘇后在含10%胎牛血清的RPMI 1640培養(yǎng)液加1%青-鏈霉素雙抗中進(jìn)行培養(yǎng),于37 ℃、5%CO2、飽和濕度的培養(yǎng)箱進(jìn)行培養(yǎng),觀察細(xì)胞生長(zhǎng)狀態(tài)并及時(shí)更換培養(yǎng)液,采用胰蛋白酶消化細(xì)胞并傳代處理,選取生長(zhǎng)狀態(tài)良好的細(xì)胞進(jìn)行后續(xù)實(shí)驗(yàn)。
1.3H37Rv感染THP-1細(xì)胞 將傳代的THP-1細(xì)胞接種到6孔板中,接種密度為1×106/ml,向每孔中加入濃度為150 ng/ml的佛波酯(PMA)(美國(guó)Sigma公司)溶液,37 ℃培養(yǎng)箱誘導(dǎo)培養(yǎng)24 h,去上清,PBS沖洗3次,再向每孔中添加2 ml無(wú)血清無(wú)雙抗的培養(yǎng)液,將H37Rv以MOI=5感染細(xì)胞[8],37 ℃培養(yǎng)箱感染4 h,棄上清,更換成新的培養(yǎng)液,此時(shí)記為感染0 h,測(cè)定CCL2 mRNA和蛋白表達(dá),隨后分別在感染6、12、24和48 h時(shí)測(cè)定CCL2 mRNA和蛋白表達(dá)。
1.4細(xì)胞轉(zhuǎn)染和分組處理 待THP-1細(xì)胞貼壁良好,根據(jù)Lipofectmine 2000轉(zhuǎn)染試劑使用說(shuō)明書分別將CCL2 siRNA或siRNA control轉(zhuǎn)染至THP-1細(xì)胞,轉(zhuǎn)染6 h后更換完全培養(yǎng)液,實(shí)驗(yàn)分為Mock(對(duì)照)組、H37Rv(感染)組、H37Rv+si-NC(降低CCL2感染對(duì)照)組和H37Rv+si-CCL2(降低CCL2感染)組,其中H37Rv+si-NC組和H37Rv+si-CCL2組的THP-1細(xì)胞于轉(zhuǎn)染48 h后,將H37Rv以MOI=5進(jìn)行感染,并在H37Rv+si-CCL2組中添加濃度為150 ng/ml的NF-κB信號(hào)通路激活劑PMA進(jìn)行干預(yù)作為H37Rv+si-CCL2+PMA組,24 h后分別收集各組細(xì)胞進(jìn)行相關(guān)指標(biāo)的檢測(cè)。
1.5RT-PCR檢測(cè)CCL2 mRNA表達(dá)量 使用TRIzol試劑分離待測(cè)各組THP-1細(xì)胞中總RNA,使用PrimeScript RT試劑盒對(duì)RNA進(jìn)行逆轉(zhuǎn)錄,按照使用說(shuō)明書采用Green Premix Ex Taq Ⅱ進(jìn)行RT-PCR檢測(cè)。以GAPDH為內(nèi)參,采用2-ΔΔCt分析CCL2 mRNA表達(dá)量。GAPDH上游引物:5'-GCACCGTCAAGGCTGAGAAC-3',下游引物:5'-TGGTGAAGACGCCAGTGGA-3'。CCL2上游引物:5'-GCTCCGGGCCCAGTATCT-3',下游引物:5' -ACAGGGAAGGTGAAGGGTATGA-3' 。
1.6Western blot檢測(cè)CCL2、LC3-Ⅰ、LC3-Ⅱ、p62、p65、IKK-α和IκB-α蛋白表達(dá)量 將待測(cè)THP-1細(xì)胞在裂解緩沖液中裂解以提取總蛋白。使用BCA蛋白濃度測(cè)定試劑盒檢測(cè)蛋白質(zhì)濃度,隨后將蛋白質(zhì)用15%SDS-PAGE分離,然后轉(zhuǎn)移到PVDF膜上。用5%脫脂牛奶封閉后,將膜與一抗(CCL2、LC3-Ⅰ、LC3-Ⅱ、p62、p65、IKK-α和IκB-α均為1∶1000稀釋)在4 ℃下孵育過(guò)夜,然后將膜與二抗(1∶3000稀釋)一起孵育2 h。使用ECL化學(xué)發(fā)光試劑盒顯影,采集圖像獲得條帶,以GAPDH進(jìn)行標(biāo)定,使用Image J軟件分析目的蛋白相對(duì)表達(dá)水平。
1.7菌落形成單位計(jì)數(shù)分析H37Rv胞內(nèi)存活率 將降低CCL2表達(dá)的THP-1細(xì)胞以2×105/孔接種到6孔板中,貼壁后將H37Rv以MOI=5進(jìn)行感染,6 h后棄上清,PBS沖洗3次,加入含20 μg/ml阿米卡星(山東齊魯制藥有限公司)的培養(yǎng)液,殺死未進(jìn)入細(xì)胞的H37Rv,在第4天以0.1%的Triton X-100裂解細(xì)胞,以10倍梯度稀釋,涂布在7H11平板中,在37 ℃培養(yǎng)箱培養(yǎng)3周,進(jìn)行菌落計(jì)數(shù)分析H37Rv胞內(nèi)存活率。
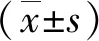
2 結(jié)果
2.1H37Rv感染THP-1細(xì)胞中CCL2表達(dá) H37Rv感染THP-1細(xì)胞不同時(shí)間CCL2 mRNA和CCL2蛋白總體比較差異有統(tǒng)計(jì)學(xué)意義(P<0.01)。H37Rv感染THP-1細(xì)胞6 h CCL2 mRNA和CCL2蛋白開(kāi)始逐漸升高,隨著感染時(shí)間延長(zhǎng)CCL2 mRNA和CCL2蛋白持續(xù)升高,于24 h達(dá)到最高水平,48 h又有所降低。H37Rv感染THP-1細(xì)胞不同時(shí)間CCL2 mRNA和CCL2蛋白兩兩比較差異均有統(tǒng)計(jì)學(xué)意義(P<0.01)。見(jiàn)表1和圖1。后續(xù)實(shí)驗(yàn)選擇感染時(shí)間為24 h。

表1 不同時(shí)間H37Rv感染THP-1細(xì)胞中CCL2表達(dá)比較

圖1 不同時(shí)間H37Rv感染THP-1細(xì)胞中CCL2蛋白表達(dá)
2.2CCL2在H37Rv感染THP-1細(xì)胞中轉(zhuǎn)染效率 Mock組、H37Rv組、H37Rv+si-NC組和H37Rv+si-CCL2組4組H37Rv感染THP-1細(xì)胞中CCL2 mRNA和CCL2蛋白表達(dá)總體比較差異有統(tǒng)計(jì)學(xué)意義(P<0.01)。與Mock組比較,H37Rv感染THP-1細(xì)胞中CCL2 mRNA和CCL2蛋白表達(dá)H37Rv組和H37Rv+si-NC組均升高;與H37Rv組和H37Rv+si-NC組比較,H37Rv感染THP-1細(xì)胞中CCL2 mRNA和CCL2蛋白表達(dá)H37Rv+si-CCL2組降低,差異均有統(tǒng)計(jì)學(xué)意義(P<0.01)。見(jiàn)表2和圖2。
表2 采用不同處理方式4組H37Rv感染THP-1細(xì)胞中CCL2表達(dá)比較

圖2 采用不同處理方式4組H37Rv感染THP-1細(xì)胞中CCL2蛋白表達(dá)
2.3CCL2降低對(duì)H37Rv感染THP-1細(xì)胞自噬影響 Mock組、H37Rv組、H37Rv+si-NC組和H37Rv+si-CCL2組4組H37Rv感染THP-1細(xì)胞中LC3-Ⅰ、LC3-Ⅱ和p62表達(dá)總體比較差異有統(tǒng)計(jì)學(xué)意義(P<0.01)。與Mock組比較,H37Rv組和H37Rv+si-NC組H37Rv感染THP-1細(xì)胞中LC3-Ⅰ和p62表達(dá)升高,LC3-Ⅱ表達(dá)降低;與H37Rv組和H37Rv+si-NC組比較,H37Rv+si-CCL2組H37Rv感染THP-1細(xì)胞中LC3-Ⅰ和p62表達(dá)降低,LC3-Ⅱ表達(dá)升高,差異均有統(tǒng)計(jì)學(xué)意義(P<0.01)。見(jiàn)表3和圖3。

圖3 采用不同處理方式4組H37Rv感染THP-1細(xì)胞中LC3-Ⅰ、LC3-Ⅱ和p62表達(dá)比較
表3 采用不同處理方式4組H37Rv感染THP-1細(xì)胞中LC3-Ⅰ、LC3-Ⅱ和p62表達(dá)比較
2.4CCL2降低對(duì)H37Rv胞內(nèi)存活影響 H37Rv組、H37Rv+si-NC組和H37Rv+si-CCL2組H37Rv胞內(nèi)存活率分別為(100.00±9.22)%、(98.57±9.14)%和(46.88±4.52)%。3組H37Rv胞內(nèi)存活率總體比較差異有統(tǒng)計(jì)學(xué)意義(P<0.01)。H37Rv組和H37Rv+si-NC組H37Rv胞內(nèi)存活率明顯高于H37Rv+si-CCL2組,差異有統(tǒng)計(jì)學(xué)意義(P<0.01)。
2.5CCL2對(duì)H37Rv感染THP-1細(xì)胞中NF-κB信號(hào)通路激活影響 Mock組、H37Rv組、H37Rv+si-NC組和H37Rv+si-CCL2組4組H37Rv感染THP-1細(xì)胞中p65、IKK-α和IκB-α表達(dá)總體比較差異有統(tǒng)計(jì)學(xué)意義(P<0.01)。與Mock組比較,H37Rv感染THP-1細(xì)胞中p65、IKK-α和IκB-α表達(dá)H37Rv組和H37Rv+si-NC組均升高;與H37Rv組和H37Rv+si-NC組比較,H37Rv感染THP-1細(xì)胞中p65、IKK-α和IκB-α表達(dá)H37Rv+si-CCL2組降低,差異均有統(tǒng)計(jì)學(xué)意義(P<0.01)。見(jiàn)表4和圖4。

圖4 采用不同處理方式4組H37Rv感染THP-1細(xì)胞中p65、IKK-α和IκB-α表達(dá)
表4 采用不同處理方式4組H37Rv感染THP-1細(xì)胞中p65、IKK-α和IκB-α表達(dá)比較
2.6PMA逆轉(zhuǎn)CCL2降低對(duì)H37Rv感染THP-1細(xì)胞自噬影響 與H37Rv+si-CCL2組比較,H37Rv+si-CCL2+PMA組H37Rv感染THP-1細(xì)胞中LC3-Ⅰ和p62表達(dá)升高,LC3-Ⅱ表達(dá)降低,差異有統(tǒng)計(jì)學(xué)意義(P<0.01),見(jiàn)表5和圖5。

圖5 H37Rv+si-CCL2組和H37Rv+si-CCL2+PMA組H37Rv感染THP-1細(xì)胞中LC3-Ⅰ、LC3-Ⅱ和p62表達(dá)
表5 H37Rv+si-CCL2組和H37Rv+si-CCL2+PMA組H37Rv感染THP-1細(xì)胞中LC3-Ⅰ、LC3-Ⅱ和p62表達(dá)比較
3 討論
結(jié)核分枝桿菌感染肺部時(shí),將導(dǎo)致結(jié)核病的發(fā)生,結(jié)核病是一種高病死率的傳染病[9]。到目前為止,尚沒(méi)有有效疫苗可以預(yù)防所有的結(jié)核分枝桿菌菌株,當(dāng)前結(jié)核病的藥物治療面臨耐藥性的障礙,故現(xiàn)迫切需要弄清結(jié)核分枝桿菌感染的病理機(jī)制,以便可以確定有效的治療靶標(biāo)和診斷生物標(biāo)志物[10]。在體內(nèi),結(jié)核分枝桿菌具有較長(zhǎng)的潛伏期,這給結(jié)核病診斷和治療帶來(lái)了挑戰(zhàn),而及時(shí)的診斷和治療對(duì)遏制結(jié)核病的傳播具有重要意義。越來(lái)越多的研究表明,結(jié)核分枝桿菌可以改變基因表達(dá),從而導(dǎo)致免疫逃逸[11-12]。以往研究發(fā)現(xiàn),CCL2在肺結(jié)核患者中異常表達(dá),提示它可能在肺結(jié)核發(fā)生和發(fā)展中起關(guān)鍵作用[13]。本研究結(jié)果發(fā)現(xiàn),H37Rv感染THP-1細(xì)胞能夠升高CCL2的表達(dá),降低CCL2的表達(dá)可通過(guò)阻斷NF-κB信號(hào)通路抑制H37Rv感染THP-1細(xì)胞自噬。
人體防御機(jī)制是病原體入侵研究必不可少的內(nèi)容。自噬是許多類型病原體感染的關(guān)鍵防御機(jī)制[14]。自噬是維持細(xì)胞穩(wěn)態(tài)的重要生理過(guò)程,可促進(jìn)細(xì)胞存活,為抵抗微生物入侵的重要防御機(jī)制。然而,在分枝桿菌感染中,分枝桿菌利用自噬以確保其細(xì)胞內(nèi)存活[15-16]。因此,在分枝桿菌感染中發(fā)現(xiàn)自噬過(guò)程背后的主要調(diào)節(jié)劑具有重要意義。本研究探討CCL2對(duì)結(jié)核分枝桿菌感染巨噬細(xì)胞自噬的影響。首先,本研究評(píng)估了CCL2和結(jié)核分枝桿菌之間的關(guān)系,觀察到結(jié)核分枝桿菌以時(shí)間依賴的方式升高CCL2的表達(dá)水平。接著,本研究確認(rèn)由結(jié)核分枝桿菌改變的防御機(jī)制是否與CCL2的異常表達(dá)有關(guān)。然后,本研究探討CCL2表達(dá)降低對(duì)巨噬細(xì)胞自噬及結(jié)核分枝桿菌存活的影響。值得注意的是,本研究發(fā)現(xiàn)CCL2表達(dá)降低增強(qiáng)了正常巨噬細(xì)胞自噬,并抑制結(jié)核分枝桿菌的存活。這一發(fā)現(xiàn)證實(shí)了結(jié)核分枝桿菌對(duì)細(xì)胞自噬的影響是通過(guò)CCL2表達(dá)降低實(shí)現(xiàn)的,這表明CCL2可能為肺結(jié)核提供了新的治療靶點(diǎn)。以往研究顯示,在結(jié)核病患者中NF-κB信號(hào)通路異常活化,且結(jié)核分枝桿菌感染后能夠激活NF-κB信號(hào)通路[17-18]。有研究顯示,大豆凝集素誘導(dǎo)的自噬通過(guò)產(chǎn)生活性氧并通過(guò)NF-κB信號(hào)通路發(fā)揮抗結(jié)核分枝桿菌的作用[19]。本研究結(jié)果同樣發(fā)現(xiàn),結(jié)核分枝桿菌感染的巨噬細(xì)胞中NF-κB信號(hào)通路關(guān)鍵蛋白p65、IKK-α和IκB-α表達(dá)升高,而降低CCL2能夠有效抑制p65、IKK-α和IκB-α的表達(dá)。此外,NF-κB信號(hào)通路激活劑PMA能夠逆轉(zhuǎn)CCL2降低對(duì)結(jié)核分枝桿菌感染巨噬細(xì)胞自噬的抑制作用。以上結(jié)果表明CCL2可能通過(guò)抑制NF-κB信號(hào)通路調(diào)節(jié)結(jié)核分枝桿菌感染巨噬細(xì)胞自噬。
綜上所述,結(jié)核分枝桿菌感染的巨噬細(xì)胞中CCL2表達(dá)升高,降低CCL2能夠激活自噬并抑制結(jié)核分枝桿菌胞內(nèi)存活,其作用機(jī)制與抑制NF-κB信號(hào)通路有關(guān)。CCL2似乎是結(jié)核分枝桿菌感染巨噬細(xì)胞中宿主防御機(jī)制的一部分,本研究結(jié)果有助于進(jìn)一步了解細(xì)胞內(nèi)穩(wěn)態(tài)在防御細(xì)菌感染中的調(diào)控機(jī)制,為結(jié)核病的治療提供實(shí)驗(yàn)基礎(chǔ)。
猜你喜歡
新少年(2022年9期)2022-09-17 07:10:54
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2020年6期)2020-12-16 02:56:41
文苑(2020年12期)2020-04-13 00:54:10
科技傳播(2019年22期)2020-01-14 03:06:54
中學(xué)生數(shù)理化·中考版(2019年12期)2019-09-23 06:23:28
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
