傳染性胰臟壞死病毒VP3抗血清的制備及應用
2021-09-17 09:55:14陳桂花賀文斌徐黎明趙景壯任廣明邵軼智段凱越盧彤巖
大連海洋大學學報 2021年4期
關鍵詞:檢測
陳桂花,賀文斌,徐黎明,趙景壯,任廣明,4,邵軼智,段凱越,盧彤巖,4*
(1.上海海洋大學 水產科學國家級實驗教學示范中心,上海 201306;2.中國水產科學研究院黑龍江水產研究所,黑龍江 哈爾濱 150070;3.上海海洋大學 農業農村部漁業動植物病原庫,上海 201306;4.中國水產科學研究院長江水產研究所,湖北 武漢 430223)
傳染性胰臟壞死病毒(infectious pancreas necrosis virus,IPNV)屬于雙RNA病毒科Birnaviridae水生雙RNA病毒屬Aquabirnavirus,是引起傳染性胰腺壞死病(infectious pancreatic necrosis,IPN)的病原體[1],其主要危害鮭鱒幼魚,致死率高達70%以上[2],同時也可使成年鮭鱒及許多其他魚類發生嚴重的急性感染,此外,在一些軟體動物和甲殼動物中也分離到了IPNV[3]。傳染性胰臟壞死病于20世紀50年代在美國首次暴發,目前,該病已在歐洲、亞洲和非洲廣泛流行[4]。1986年在中國山西省某虹鱒Oncorhynchusmykiss漁場首次暴發傳染性胰腺壞死病(IPN)[5],隨后在甘肅[6]、山東[7]、遼寧[8]、云南[9]和四川[10]等省份接連暴發了該疫情,患病虹鱒死亡率高達90%,給鮭鱒養殖業帶來了巨大的經濟損失。
IPNV基因組由A、B兩個線性dsRNA分子組成,A鏈編碼非結構蛋白VP5和pVP2-VP4-VP3多聚蛋白前體[11],其中,VP4作為病毒蛋白酶將多聚蛋白切開釋放結構蛋白VP2和VP3[12];B鏈編碼聚合酶蛋白VP1[13]。許多學者已對VP2蛋白的結構和功能進行了研究,并首先確定了VP2蛋白是IPNV的主要保護性抗原[14-16]。Tarrab等[17]利用抗VP3單克隆抗體檢測到病毒粒子表面有部分VP3蛋白存在;Park等[18]研究發現,VP3蛋白也存在中和抗原表位;Moon等[19]用純化后的VP3和VP2蛋白分別免疫虹鱒,也證明VP3蛋白和VP2蛋白一樣,具有中和抗原表位,且具有較好的免疫原性。因此,VP3蛋白也是IPNV重要的保護性抗原。
盡管IPNV在中國流行多年,但是目前中國仍無有效防控傳染性胰臟壞死病的藥物,對待該病的防控方法主要以切斷病原傳播為主,故而對IPNV的全面監測尤為重要。目前,全球IPNV可分為6個基因型(Genogroup Ⅰ~genogroup Ⅵ),共包含9個血清型[20-21],不同基因型IPNV毒株的基因序列差異較大,核酸檢測方法在IPNV檢測中具有一定的局限性,較難實現使用一對引物覆蓋所有基因型的IPNV毒株。因此,極有必要豐富IPNV的檢測方法,為IPNV的有效監測提供保障。本研究中,選取IPNV毒株ChRtm213[9],利用大腸桿菌對其VP3基因進行體外表達及純化,同時制備鼠抗血清,并成功將該血清應用到中國不同地區Genogroup Ⅰ和Genogroup Ⅴ型IPNV分離株的免疫學鑒定,旨在為IPNV的監測提供保障。
1 材料與方法
1.1 材料
本研究中所用IPNV和傳染性造血器官壞死病毒(IHNV)均由黑龍江省水產動物疾病與免疫技術重點實驗室在中國養殖虹鱒流行病學調查過程中分離保存,其中,IPNV-QH、IPNV-LN和IPNV-GS的基因型為Genogroup Ⅴ,ChRtm213[9]、IPNV-JL和IPNV-HLJ的基因型為Genogroup Ⅰ。
病毒性出血性敗血癥病毒(VHSV)毒株購自中國典型培養物保藏中心。大鱗大麻哈魚胚胎細胞(chinook salmon embryo cells,CHSE-214)由中國水產科學研究院長江水產研究所魚類病害研究室曾令兵教授惠贈;大腸桿菌DH5α和Rosetta菌株及pET-32a表達載體均由本課題組保存。
pMD19-T simple載體、T4 DNA連接酶、限制性內切酶、PrimeScriptTMOne Step RT-PCR Kit Ver 2.0均購自寶生物工程(大連)有限公司;PCR產物純化及膠回收試劑盒、質粒提取試劑盒購自Omega公司;HRP標記的羊抗鼠IgG和FITC標記的羊抗鼠IgG購自Abcam公司;其他化學試劑均為分析純。
1.2 方法
1.2.1 引物設計 根據IPNV ChRtm213毒株VP3基因序列(NCBI登錄號:KX234591),利用Primer Premier 5.0 軟件設計擴增IPNVVP3基因的引物。上游引物VP3-F帶有BamH Ⅰ酶切位點:5′ GGATCCATGGACGCAGAACTGCAAGGGCTGC 3′;下游引物VP3-R帶有SalⅠ酶切位點:5′ GTCGACCACTTCTCCGTCATCGCCGGAG 3′。引物均由哈爾濱博仕生物公司合成。
1.2.2 IPNVVP3 基因的克隆及表達載體構建 以實驗室保存的IPNV(ChRtm213)RNA為模板,以VP3-F和VP3-R為引物,利用一步法RT-PCR試劑盒克隆獲得VP3 ORF,膠回收純化后與pMD19-T simple連接,菌液經PCR和測序鑒定后,將測序正確的質粒命名為pMD19-T-VP3。將pMD19-T-VP3和表達載體pET-32a分別利用BamH Ⅰ和SalⅠ限制性內切酶消化并回收,將使用T4 DNA連接酶連接后的產物轉化至大腸桿菌DH5α中,涂布于含氨芐西林的瓊脂糖固體培養基,于37 ℃下過夜培養后挑取單菌落,進行菌液PCR鑒定,將鑒定正確的菌液擴大培養并提取質粒,對質粒進行BamH Ⅰ和SalⅠ雙酶切鑒定,鑒定正確的重組質粒命名為pET-32a-VP3。
1.2.3 IPNV VP3 蛋白的表達與純化 將重組pET-32a-VP3質粒轉入大腸桿菌Rosetta中,于37 ℃下以100 r/min振蕩培養。當菌液OD600 nm值達到0.3~0.4時,加入IPTG誘導劑(終濃度為0.25 mmol/L)繼續培養。誘導后5 h內每小時收集1 mL菌液,同時收集未誘導菌液作為陰性對照。經超聲破碎后,將全菌、上清及沉淀樣本用質量分數12%的SDS-PAGE凝膠進行電泳,以分析VP3蛋白的表達情況。
超聲破碎后的上清液使用鎳離子親和層析柱純化,依次使用含有10、20、30、40 mmol/L咪唑的洗滌液洗滌層析柱以去除雜蛋白,然后利用含有250 mmol/L咪唑的洗脫液收集目的蛋白,放入PBS溶液中(pH 8.0)透析24 h。透析結束后,于4 ℃下以12 000 r/min離心5 min,收獲的上清液即為純化的VP3蛋白,用紫外分光光度計測定純化后蛋白的濃度。利用質量分數12%的SDS-PAGE電泳分析VP3蛋白的純化情況。
1.2.4 IPNV VP3 蛋白抗血清的制備 將純化后的VP3蛋白無菌過濾,采用0.5 mg/kg的免疫劑量,根據文獻[14]中的方法對Balb/c小鼠進行皮下多點注射。將重組VP3蛋白與弗氏完全佐劑等體積混合,乳化后免疫小鼠;免疫10 d后進行第2次免疫,將重組VP3蛋白與弗氏不完全佐劑等體積混合,完全乳化后免疫小鼠;第2次免疫10 d后進行第3次免疫,免疫方式與第2次相同;第3次免疫10 d后,通過眼球取血的方式收集免疫小鼠血液,進行血清分離,同時以PBS替代抗原免疫的小鼠血液制備的抗血清作為陰性對照。
1.2.5 IPNV VP3 蛋白抗血清效價檢測 用IPNV病毒懸液和重組VP3蛋白分別進行包被,根據文獻[14]中的酶聯免疫吸附法(ELISA)檢測抗血清蛋白效價。將“1.2.4節”中制備的鼠抗血清進行梯度稀釋(體積比分別為1∶500、1∶1 000、1∶2 000、1∶4 000、1∶8 000、1∶16 000、1∶32 000、1∶64 000、1∶128 000),與包被的酶標板于37 ℃下孵育1 h。然后與HRP標記的羊抗鼠IgG(1∶10 000,稀釋)于37 ℃下孵育1 h,用PBS洗去未被吸附的二抗,最后加入TMB底物液進行顯色,置于37 ℃暗箱中反應5 min,每孔加入50 μL終止液。終止反應后于490 nm下測定OD值。
陰性對照為免疫PBS與佐劑的鼠血清,空白對照為PBS。當OD490 nm所讀數值符合(陽性孔-空白孔)/(陰性孔-空白孔)>2.1時,判定樣本為陽性,陽性血清最大稀釋倍數即為抗血清效價[14]。
1.2.6 抗血清的應用 將所制備的抗血清應用到中國不同地區分離到的不同基因型的IPNV分離株的間接免疫熒光抗體法(IFAT)檢測中。在CHSE-214單層細胞爬片上接種本實驗室分離的6株IPNV毒株和IHNV、VHSV,在接毒后72 h取出接毒細胞爬片。將制備好的接毒細胞爬片用體積分數為4%的多聚甲醛固定,用PBS(pH 7.0)緩沖液洗滌3次,用體積分數為0.3%的BSA染色溶液于37 ℃下封閉1 h,再用PBS緩沖液洗滌,加入500倍稀釋的鼠抗VP3蛋白血清作為一抗,37 ℃下孵育1 h,再用PBS緩沖液洗滌,將FITC標記的羊抗鼠IgG抗體稀釋500倍后作為二抗,37 ℃下避光孵育1 h[22]。最后用PBS緩沖液洗滌后置于熒光顯微鏡下觀察。設置陰性血清孵育的細胞作為陰性對照。
2 結果與分析
2.1 VP3 基因的克隆及表達載體的構建
以IPNV ChRtm213 RNA為模板,VP3-F和VP3-R為引物擴增VP3 ORF,經10 g/L瓊脂糖凝膠電泳,結果顯示,獲得了符合目片段大小的PCR產物(711 bp)(圖1)。利用BamH Ⅰ和SalⅠ對重組質粒pET-32a-VP3進行酶切鑒定,結果獲得與VP3基因和表達載體pET-32a大小相符的特異性條帶(圖2),說明已成功構建了用于VP3原核表達的重組質粒。
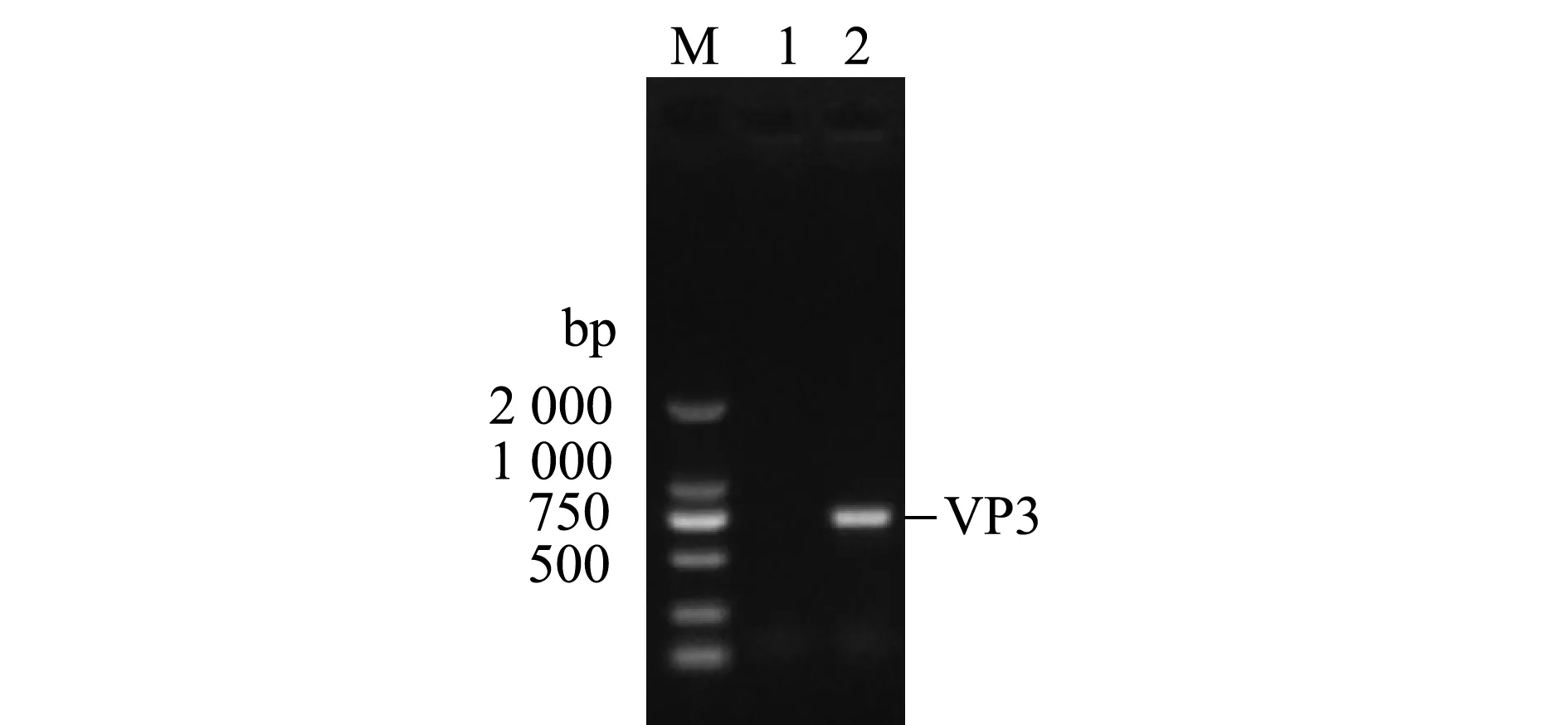
M—DL2000 DNA分子量標準;1—陰性對照;2—VP3基因的PCR產物。
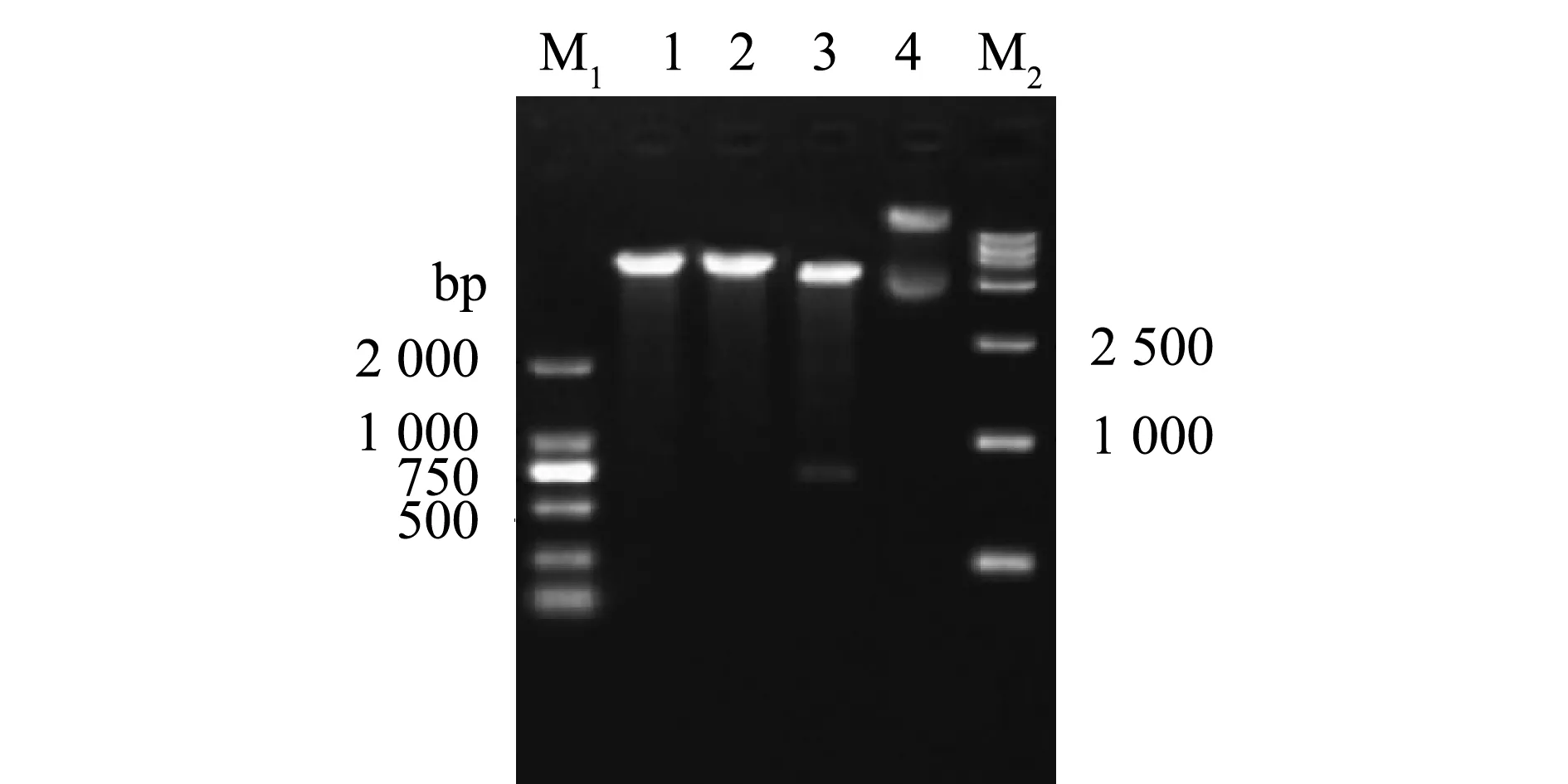
M1—DL2000 DNA分子量標準;M2—DL15000 DNA分子量標準;1—BamH Ⅰ單酶切; 2—Sal Ⅰ單酶切;3—BamH Ⅰ和Sal Ⅰ雙酶切;4—pET-32a-VP3質粒。
2.2 VP3 蛋白的誘導表達
將陽性pET-32a-VP3重組質粒轉入大腸桿菌Rosetta進行誘導表達,對表達產物進行SDS-PAGE分析,結果顯示,VP3蛋白成功獲得表達,出現與目的條帶大小相符的特異性蛋白條帶(相對分子質量約為45 000),且隨著誘導時間的延長,表達量逐漸上升(圖3),在誘導后3 h表達量顯著上升。此外,本研究中還發現,菌體破碎離心后的沉淀和上清液中均存在目的蛋白(圖4),說明本研究中所表達的VP3蛋白以包涵體和可溶性蛋白兩種形式存在。
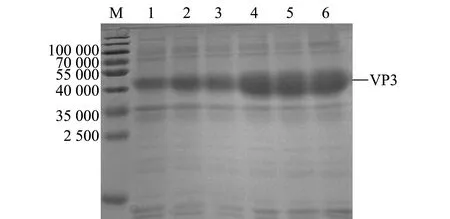
M—蛋白分子量標準;1—未誘導菌體;2~6—誘導1、2、3、4、5 h后菌體。
2.3 VP3 蛋白的純化
將超聲破碎后所得上清經鎳離子親和層析柱純化和PBS(pH 8.0)透析24 h后,對樣品進行SDS-PAGE凝膠電泳分析,結果顯示,純化的VP3蛋白與目的條帶大小相符且為單一條帶,可用于VP3蛋白鼠抗血清的制備(圖5)。

M—蛋白分子量標準;1—誘導前破碎菌體上清;2~8—誘導4 h破碎菌體上清;9—純化后的VP3蛋白。
2.4 IPNV VP3 蛋白鼠抗血清效價的檢測
利用本研究中所制備的抗血清對包被在酶標板上的IPNV和重組VP3蛋白進行酶聯免疫吸附法檢測。根據判定標準可知,該抗血清與IPNV的反應效價為16 000,與重組VP3蛋白的反應效價為32 000(圖6)。這表明,本研究中所制備的抗血清不僅能與重組VP3蛋白發生特異性反應,還能與IPNV病毒粒子中的VP3蛋白發生特異性反應。證明本研究中所制備的重組VP3蛋白具有較好的免疫原性,所制備的抗VP3蛋白的鼠血清可以應用到IPNV的免疫學檢測。
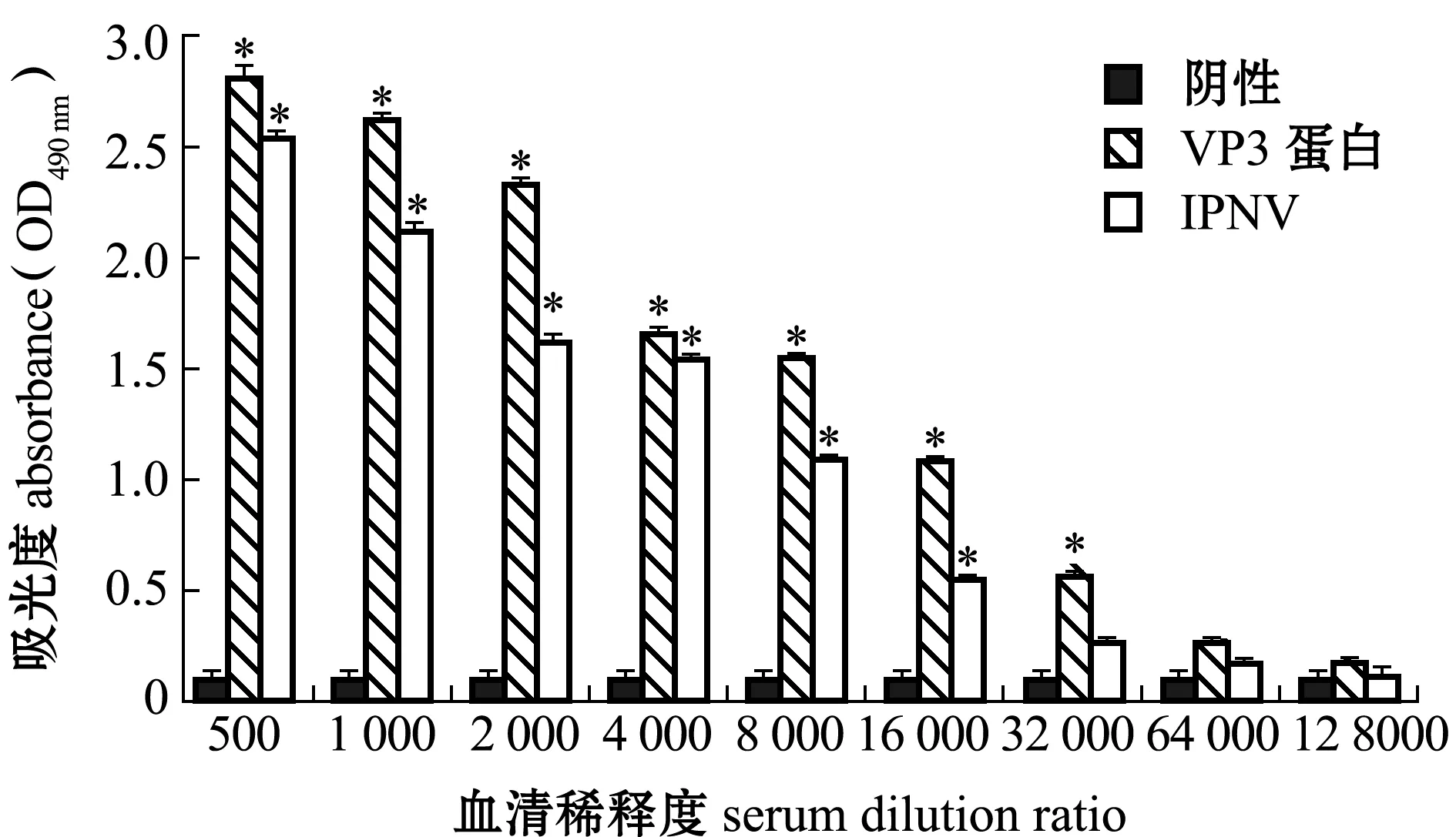
*表示與陰性對照組有顯著性差異(P<0.05)。
2.5 抗血清的應用
將制備的鼠抗血清應用到中國不同地區IPNV分離株的間接免疫熒光檢測中,結果顯示,抗血清與Genogroup Ⅰ型(IPNV-JL、IPNV-HLJ、ChRtm213)和Genogroup Ⅴ 型(IPNV-GS、IPNV-QH、IPNV-LN)的IPNV分離株均能發生特異性反應,呈現出特異性綠色熒光,而不能與IHNV和VHSV發生反應,無特異性熒光顯色,陰性鼠血清孵育的病毒均無任何熒光(圖7)。這表明,本研究中制備的鼠抗血清能夠特異性識別不同地區基因型為Genogroup Ⅰ和Genogroup Ⅴ的IPNV分離株,且與其他鮭鱒常見病毒IHNV和VHSV均無交叉反應,可用于現行IPNV的監測。

圖7 VP3蛋白鼠抗血清的間接免疫熒光抗體檢測結果
3 討論
傳染性胰臟壞死病是鮭鱒魚類最為嚴重的傳染性疾病之一。根據毒株、宿主和環境因素的不同,傳染性胰臟壞死病可造成患魚10%~90%的死亡率[2,23],目前,中國尚無有效防控傳染性胰臟壞死病的藥物,該病的防控主要依賴切斷病原的傳播與擴散。由于IPNV血清型種類較多,且不同血清型毒株間免疫交叉反應程度不同。因此,針對中國現行IPNV的免疫學檢測方法的建立對傳染性胰臟壞死病的監測尤為重要。
3.1 中國現行IPNV的基因型
中國首次暴發傳染性胰臟壞死病是在20世紀80年代,至今已有30余年。由于缺乏有效的防控藥物與監管措施,IPNV在中國養殖環境中快速傳播與擴散。流行病學調查顯示,近年來,在中國東北、西北、西南等虹鱒主養區均有IPNV檢出,給當地虹鱒養殖業造成了巨大的經濟損失。從中國養殖環境中分離到的IPNV主要包含Genogroup Ⅰ型[9-10]和Genogroup Ⅴ型兩種基因型,這兩種基因型分別屬于不同的血清型。本實驗室近年在虹鱒流行病學調查過程中獲得大量的Genogroup Ⅰ型IPNV分離株,因此,本研究中采用了課題組于2013年從中國某虹鱒養殖場分離的Genogroup Ⅰ型毒株制備了抗血清。為了檢驗該抗血清是否能應用到中國現存的兩種基因型IPNV的監測,本研究中應用間接免疫熒光抗體法對抗血清識別IPNV分離株的能力進行了檢驗,結果發現,所制備的抗血清不但能夠特異性識別Genogroup Ⅰ型IPNV毒株,而且能夠特異性識別Genogroup Ⅴ型IPNV,并且該血清不與鮭鱒常見病毒IHNV和VHSV存在交叉反應。本研究中所制備的抗血清在IPNV的監測中將會發揮重要作用。
3.2 VP3蛋白在IPNV免疫學檢測中的應用
VP2和VP3蛋白均為IPNV的主要結構蛋白,而VP2蛋白更多地被用于IPN的檢測及免疫預防研究[24-28]。有研究表明,VP3和VP2蛋白同樣具有抗原表位和中和抗體表位[17-18],且具有良好的免疫原性和反應原性[29]。因此,目前逐漸有學者開始利用VP3蛋白作為目標蛋白建立IPNV檢測方法,如Espinoza等[30]和Milne等[25]利用VP3單克隆抗體分別建立了IPNV免疫印跡法和RT-PCR-ELISA法,均能實現IPNV的高效檢測。此前,本課題組已制備了IPNV VP2蛋白的抗血清[14],且在本研究中表達了IPNV VP3蛋白,制備了VP3抗血清,不但豐富了IPNV免疫學檢測手段,還可為VP3蛋白結構和功能的深入研究提供物質基礎。
3.3 本研究中所制備的抗血清的應用價值
此前中國學者曾體外表達了Genogroup Ⅴ型IPNV的VP3蛋白[24],制備了單克隆抗體[31],成功將該抗體應用到Genogroup Ⅴ型IPNV的免疫學檢測中,但并未驗證其是否可應用于Genogroup Ⅰ型IPNV的檢測。通過文獻查閱可知,中國最初流行的IPNV為Genogroup Ⅴ型,Genogroup Ⅰ型的IPNV是近年分離鑒定的新毒株[9-10]。針對該現狀,本研究中選擇基因型為Genogroup Ⅰ的IPNV分離株,利用大腸桿菌Rosetta進行VP3蛋白的表達,經過鎳離子親和層析柱純化,獲得大量可溶性VP3蛋白,并成功制備了能夠特異識別中國不同地區IPNV分離株的抗血清,該抗血清既可識別基因型為Genogroup Ⅰ的IPNV毒株,還可識別基因型為Genogroup Ⅴ的IPNV毒株,具有重要的現實意義。
4 結論
1)本研究中利用大腸桿菌Rosetta體外表達制備了Genogroup Ⅰ型IPNV ChRtm213毒株的重組VP3蛋白。
2)利用重組VP3蛋白制備了鼠抗血清,該血清不但能夠特異性識別Genogroup Ⅰ型IPNV毒株,還能識別Genogroup Ⅴ型IPNV毒株,可用于IPNV的監測。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
