海南17?a宿根巴西蕉園土壤微生物特征及土壤pH周年變化特征分析
2021-09-14 14:51:30郭素霞程志號孫佩光孫長君郭剛吳瓊
熱帶作物學報 2021年8期
郭素霞 程志號 孫佩光 孫長君 郭剛 吳瓊


摘 ?要:香蕉是全球性的大宗經濟水果,我國是香蕉生產大國也是消費大國,但產量和品質不能滿足需求,每年仍需大量進口。香蕉枯萎病是影響我國香蕉產能的主要因素。巴西蕉在我國曾廣泛種植,但易感香蕉枯萎病,罕有宿根蕉園。宿根蕉園栽培模式在成本控制、產期調節、環境友好等方面有巨大優勢。前期研究結果顯示,土壤pH以及土壤菌群與香蕉枯萎病的發生有緊密聯系。本文以海南臨高皇桐鎮的17 a宿根巴西蕉園為樣本,對該宿根蕉園、附近輪作蕉園及新開墾蕉園全生育期土壤pH及香蕉枯萎病孢子數量進行動態監測,結合土壤菌群宏基因組測序的方法對土壤中的菌群特征進行了初步探索。結果表明:宿根蕉園土壤pH高于輪作蕉園和新開墾蕉園,其pH周年變化幅度低于輪作蕉園和新開墾蕉園;宿根蕉園土壤中香蕉枯萎病孢子數目低于輪作蕉園和新開墾蕉園;在土壤細菌和真菌的菌群多樣性和豐度方面,宿根蕉園均高于輪作蕉園和新開墾蕉園;宿根蕉園在全營養菌門、農桿菌門、毛霉亞門等11個細菌門級和6個真菌門級分類單位上與輪作和新墾蕉園存在顯著差異;宿根蕉園在微單孢菌、芽胞桿菌、角菌根菌等土壤特征種群方面顯著不同于其他2個蕉園;功能豐度熱圖顯示,宿根蕉園土壤細菌和真菌與輪作蕉園、新開墾蕉園均有顯著差異。代謝通路基因豐度分析方面顯示,輪作蕉園和新墾蕉園土壤細菌在生物合成和次級代謝、轉錄、碳代謝、膜轉運等通路明顯活躍,宿根蕉園土壤真菌在細胞遷移、細胞生長和死亡、信號傳導、核酸代謝、蛋白折疊儲運和降解等通路顯著活躍。本研究結果為闡明香蕉宿根栽培模式的內部機理奠定基礎,也為廢棄蕉園改造提供新的參考依據。
關鍵詞:巴西蕉;宿根蕉園;土壤菌群特征;香蕉枯萎病;pH
Abstract: Banana is one of the most widely planted fruits in the world and is a staple for millions of people worldwide. China is a large banana production country but still needs to import huge amounts of banana to meet the demand. Brazil banana has been widely planted in China but it is very susceptible to Fusarium wilt (Panama disease). Ratoon banana filed (RTF) is common in foreign country and famous for its low cost, easy production period regulation and environmental friendliness. RTF is seldom used in China. In this study, the microbial community structure of 17-year Lingao RTF banana field was characterized through ITS and 16S rRNA gene sequencing and the reclaim banana field (RCF) and banana cane rotation field (BCF) nearby were used as the control. The results showed that the diversity and abundance of soil bacteria and fungi in RTF were significantly higher than that of RCF and BCF. The classification of soil microbial species showed that RTF were significantly different among 11 bacteria and 6 fungal taxonomic units such as Agrobacterium tumefaciens, actinomycetes and so on. RTF soil characteristic microorganism showed significantly different with RCF and BCF in Micromonospora, Bacillus and Ceratobasidium. The functional abundance heat map showed that the soil bacterial activity in RTF was significantly lower than that of RCF and BCF, while the fungal activity was significantly higher. Gene abundance analysis of metabolic pathways showed that soil bacteria in RCF and BCF were significantly more active in biosynthesis and secondary metabolism, transcription, carbon metabolism, membrane transfer and other pathways, while soil fungi in RTF was significantly more active in cell migration, cell growth and death, signal transduction, nucleic acid metabolism, protein folding, storage, transportation and degradation. Through the analysis of soil microbial characteristics of RTF, it would lay a foundation for clarifying the internal mechanism of banana ratoon model, and also provide a new reference for the renew of abandoned banana garden.
Keywords: Brazil banana; ratoon banana filed; soil flora characteristics; Fusarium wilt of banana; pH
香蕉是全球產量最高的熱帶水果,我國是香蕉生產大國也是消費大國,但國內的產量和品質不能滿足需求,約有一半的香蕉需要進口[1]。香蕉枯萎病是土傳性病害,近距離傳播途徑以土壤、水流為主,遠距離傳播通過帶病芽、苗,研究發現其病原菌在無寄主的情況下仍可在土壤中存活30?a,因此極難防治,目前尚無理想的防治藥劑和防治方法[2]。香蕉枯萎病對我國香蕉產業沖擊巨大,導致大量蕉園荒廢,不適宜繼續種植香蕉。對于廢棄蕉園,通過輪作甘蔗[3-5]、辣椒[6-7]、水稻[8-10],或通過套種韭菜[11-13]、花生[14]等,能有效降低土壤香蕉枯萎病孢子數量,實現廢棄蕉園重新復墾種植香蕉。此外通過改種抗病品種,如‘南天黃[15-16]、‘寶島蕉[17]等在一定程度上可以減輕香蕉枯萎病的危害。同時,通過新開墾蕉園的方式,也能有效降低香蕉枯萎病的爆發,但新開墾蕉園的潛力也比較有限。雖然通過輪作、土壤改良、更換抗病品種香蕉等方式,能使部分蕉園恢復種植,但由于成本居高不下、技術難度大、操作繁瑣等原因,限制了相關技術的應用和推廣。基于上述多方面原因,導致國內香蕉種植面積近5?a來持續下滑[18]。據報道,國外存在大量的宿根蕉園,部分宿根蕉園能連作70 a[19]。香蕉宿根種植在種植成本、抗病害、調節產期、產蕉的產量和質量上比國內現有種植模式有非常明顯的優勢[19-20],但目前國內對于香蕉宿根連作模式的研究報道還非常少見。
已有研究表明,蕉園土壤pH和土壤菌群與蕉園香蕉枯萎病發生有非常緊密的關系。對海南23個香蕉園土壤微生物數量與18個土壤因子的相關性研究發現,蕉園土壤5~30 cm土層可培養微生物的區系依次為細菌、放線菌、真菌;有效Fe、有效P、土壤pH分別是影響蕉園土壤真菌數量、放線菌數量和土壤微生物總數的關鍵因子[21]。研究表明,土壤pH分別與枯萎病發病率、病情指數呈極顯著負相關[22],石灰、木薯渣、蔗渣等的施用提高了土壤pH,可以降低尖孢鐮刀菌生理小種的致病力[23]。先前研究發現,雖然不同蕉園土壤理化性質及養分含量存在很大差異,但與香蕉枯萎病的發生無直接相關性;蕉園土壤微生物數量的改變是香蕉枯萎病發病的重要特征,枯萎病危害程度為中度、重度的土壤中細菌和放線菌數量明顯低于危害程度為輕度和無危害的土壤,危害程度為中度、重度的土壤真菌數量明顯高于危害程度為輕度和無危害的土壤[24]。
植物土傳病害的生物防治與土壤微生物群落關系緊密,一定程度上依賴于調控土壤微生物群落及其功能多樣性,土壤微生物群落結構越豐富,物種多樣性越高,對病原菌的綜合抗性能力越強[25]。對于土壤微生物群落特征的研究主要有下面幾種方法:基于可培養微生物形態學和營養特征進行分離培養的方法,如平板稀釋涂布法[26];基于可培養微生物化學成分差異進行分析的方法,如脂肪酸譜圖法等[27];基于土壤微生物DNA的分子生物學技術,如高通量測序法的宏基因組學技術[28]。通過土壤微生物群落特征與土傳病害發生之間的關系進行探索,可以發現一些可能與病害發生相關的微生物類群和相關表征通路,為更好地闡明土壤微生物變化對土傳病害的影響提供了新的思路及方法[29]。
巴西蕉是我國早期引進的香蕉品種,綜合性狀明顯優于其他主推品種,但由于極易感香蕉枯萎病導致種植面積大量萎縮。對海南省臨高縣皇桐鎮的一個面積達4 hm2的17 a宿根巴西蕉園經過多年田間跟蹤調查發現,全蕉園未見真菌型香蕉枯萎病發生,蕉園香蕉品質特別是可溶性糖等的含量顯著高于附近蕉園,蕉園成本也顯著低于附近蕉園。該蕉園是國內難得的研究香蕉宿根模式的樣本。本文以該宿根巴西蕉園、附近輪作蕉園及新開墾蕉園的土壤為材料,通過對土壤全年香蕉枯萎病孢子數目的動態統計,結合土壤菌群宏基因組測序的方法,對3處土壤菌群特征進行分析,探求不同種植模式下土壤菌群與香蕉枯萎病發生的關系,為解析宿根巴西蕉園模式成功運行的原因奠定基礎,也為我國未來香蕉產業發展提供新思路和新方法。
1 ?材料與方法
1.1 ?材料
17 a宿根巴西蕉(Musa acuminata Colla)蕉園位于海南省臨高縣皇桐鎮武維村,以該蕉園附近原種植香蕉,但由于枯萎病爆發,已改種3?a甘蔗的廢棄蕉園(記為輪作蕉園)土樣為對照(CK);武維村17 a宿根巴西蕉園附近新開墾土地種植巴西蕉園(記為新開墾蕉園),土樣記為T1;武維村17 a宿根巴西蕉園(記為宿根蕉園)土樣記為T2。各蕉園香蕉品種均為巴西蕉。
1.2 ?方法
1.2.1 ?土壤取樣 ?海南蕉園田間香蕉枯萎病爆發時間集中在5~10片葉苗期和抽蕾期,蕉園留芽和定植的時間分別為2018年7月和4月,土壤pH和香蕉枯萎病孢子數目檢測每個月取樣1次,土壤菌群特征分析取樣時間為2019年3月夏蕉抽蕾前。取樣點盡量避免路邊和行道及其附近,以減少邊際效應。取樣前刨去表層有機質含量豐富的表層土3~5 cm,按照“S”形采樣路線,隨機選取5個點進行取樣,用采樣器(直徑為2.5 cm)采集0~20 cm耕作層土壤,取其下表層土壤檢測土壤pH、肥力及香蕉枯萎病菌孢子含量,深層土壤檢測土壤菌群[30]。每個取樣點3次重復,各小區采集樣品中隨機取5份土壤樣品破碎混勻,冰盒帶回實驗室后,取出一部分用于香蕉枯萎病菌孢子含量檢測,留取部分微生物菌群分析的樣品于-80?℃冰箱保存備用。
1.2.2 ?土壤pH及香蕉枯萎病孢子數目檢測 ?土壤pH檢測參考國家標準GB 7859—1987;通過平板法對蕉園土壤香蕉枯萎病菌孢子含量進行檢測,具體方法參考景曉輝等[31]、韓寶坤等[32]的文獻。
1.2.3 ?土壤菌群16S rDNA測序和ITS DNA測序 ?土壤樣品采集、制備和貯存參考國家標準GB 7830—2011;通過HiPure Soil DNA Kits(美吉生物,廣州,中國)抽提土壤微生物總DNA含量;利用NanoDrop 2000和1%瓊脂糖凝膠電泳檢測DNA提取質量和濃度。用土壤細菌16S rDNA(V3-V4)基因的通用PCR引物:341F: 5-CCTACGGGNGGCWGCAG-3,806R: 5-GGA CTACHVGGGTATCTAAT-3;ITS rDNA測序(ITS2)基因的通用PCR引物:KYO2F: 5-GATGAAGAACGYAGYRAA-3,ITS4RT: 5- CCTCCGCTTATTGATATGC-3。經過2輪PCR擴增后,使用AM Pure XP Beads對第2輪擴增產物進行純化,用ABI Step One Plus Real-Time PCR System(Life Technologies,美國)定量后,使用NovaSeq 6000的PE250模式進行序列測定。土壤樣品文庫的構建、測序以及數據分析委托廣州基迪奧生物科技有限公司進行。
1.2.4 ?土壤菌群特征分析 ?對原始數據進行過濾、拼接并去除嵌合體后得到最終有效數據。使用QIIME(Version 1.91)對最終有效數據進行聚類,并基于細菌和真菌分類數據庫進行注釋和計算α多樣性指數。使用Muscle(Version 3.8.31)軟件進行β多樣性分析,通過PICRUSt進行群落功能預測。在有效分組存在的情況下,進行組間差異比較及差異檢驗。
2 ?結果與分析
2.1 ?宿根蕉園土壤pH和香蕉枯萎病菌孢子含量
蕉園土壤香蕉枯萎病孢子數目與香蕉枯萎病爆發的幾率以及發生的強度密切相關。本研究檢測了宿根蕉園、輪作蕉園、新開墾蕉園的香蕉完整生長發育周期的土壤香蕉枯萎病孢子數目,3種蕉園的孢子數均在3月、8月、9月、10月相對含量較高(圖1A)。進一步對比分析表明,3種蕉園的孢子數目之間存在極顯著差異:宿根蕉園土壤香蕉尖孢鐮刀菌數目最少,平均為(622±144)個/g土樣,顯著低于輪作蕉園和新墾蕉園;新開墾的蕉園土壤香蕉尖孢鐮刀菌數目高于宿根蕉園,但低于輪作蕉園,為(884±119)個/g土樣;輪作蕉園土壤香蕉枯萎病病菌孢子尖孢鐮刀菌數目含量最高,平均為(1639±198)個/g土樣(圖1B)。
蕉園土壤pH也是影響香蕉枯萎病爆發的重要因素。本研究對宿根蕉園、輪作蕉園、新開墾蕉園的香蕉完整生長發育周期的土壤pH進行了動態監測(圖2A),比較分析發現:宿根蕉園土壤pH最高,平均為5.91±0.13;新開墾蕉園土壤pH最低,平均為5.51±0.12,而輪作蕉園pH介于二者之間,為5.69±0.13(圖2B)。
2.2 ?3種蕉園土壤微生物16S和ITS的OTU分析
OTU(operational taxonomic units)是指為了方便在系統發生學或群體遺傳學研究,人為設定的分類單元。利用effective tags之間的序列相似性關系,可以將不同的tags聚類成OTU。獲得OTU之后,利用相關軟件,根據其豐度和序列信息,能夠逐一開展物種注釋、群落多樣性、組間差異等多種核心分析。為了研究樣品物種的組成多樣性信息,用Uparse軟件對所有樣品的全部effective tags序列聚類,默認提供以97%的一致性將序列聚類成為OTUs結果,并計算出每個OTU在各樣品中的Tags絕對豐度和相對信息。在OTUs構建過程中,對不同樣品的effective tags數據,低頻數的Tags數據和Tags注釋數據等信息進行初步統計。通過對土壤微生物16S和ITS的測序結果進行統計,結果如圖3所示:3種蕉園土壤樣本的檢測序列一致性和豐度均比較高,達到相關要求,可進行下一步分析。通過韋恩聚類分析各蕉園土壤的物種組成與分度進一步發現,各土壤采集點間多態性比較高,其中宿根蕉園土壤多態性最高。
2.3 ?3種蕉園土壤微生物物種的16S和ITS測序分析
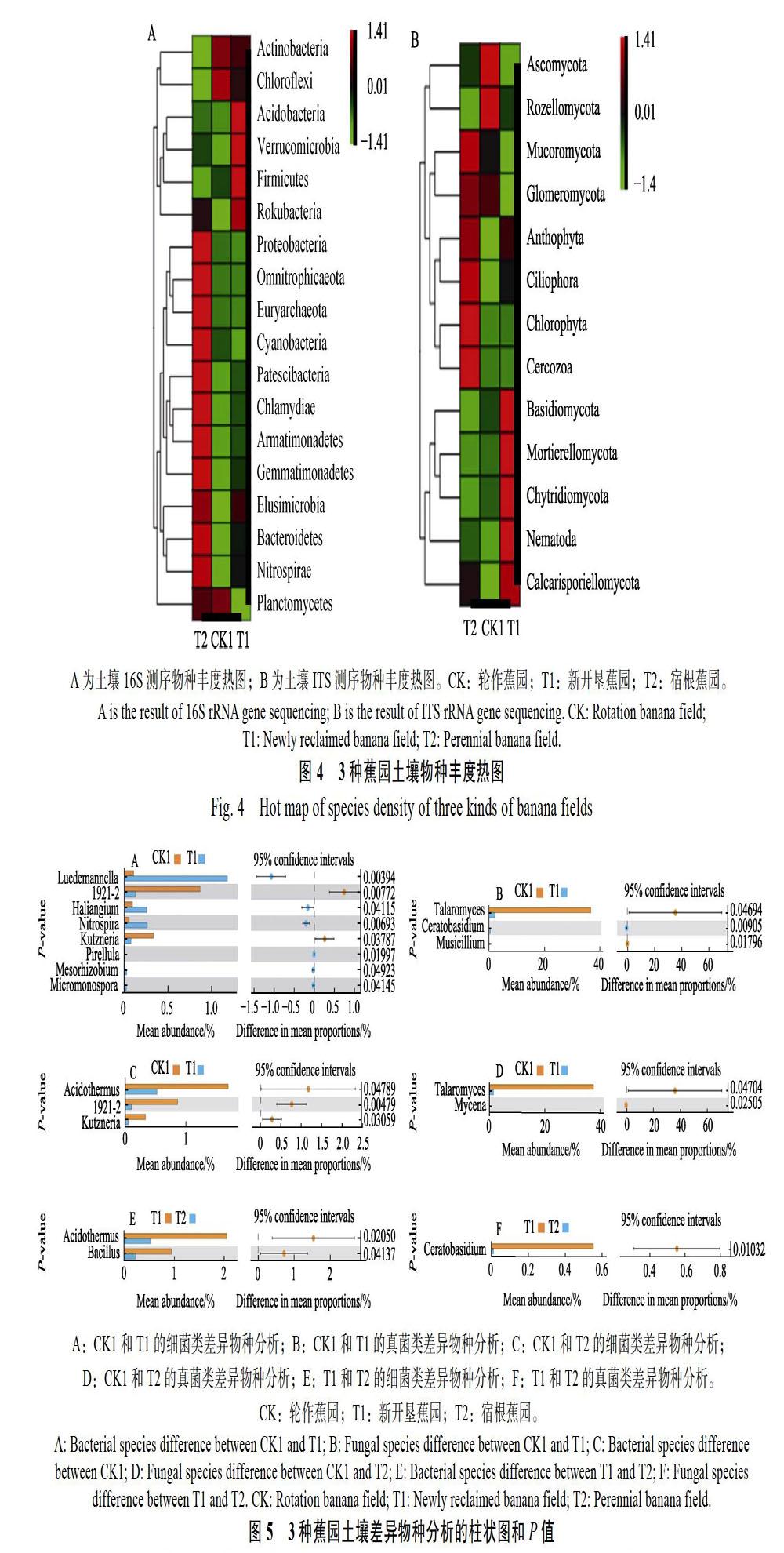
通過物種分類也可以分析不同樣本的物種組成與物種豐度,用于比較不同樣本的物種差異,結合物種分布堆疊圖、物種豐度熱圖和差異物種分析等角度來分析不同樣本的物種差異性。通過16S測序,物種豐度熱圖分析發現,新開墾蕉園與輪作蕉園更接近,宿根蕉園與輪作蕉園和新開墾蕉園均有顯著差異;與新開墾蕉園和宿根蕉園相比,輪作蕉園在放線菌門(Actinobacteria)和浮霉菌門(Planctomycetes)2個門有明顯富集;相對于輪作蕉園和宿根蕉園,新開墾蕉園在酸桿菌門(Acidobacteria)、疣微菌門(Verrucomicrobia)、厚壁菌門(Firmicutes)有明顯富集;與輪作蕉園和新開墾蕉園相比,宿根蕉園在變形菌門(Proteobacteria)、廣古菌門(Euryarchaeota)、藍細菌(Cyanobacteria)等11個門(屬)有明顯富集(圖4A)。通過ITS測序,物種豐度熱圖分析發現,新開墾蕉園與輪作蕉園更接近,宿根蕉園與輪作蕉園和新開墾蕉園均有顯著差異;與新開墾蕉園和宿根蕉園相比,輪作蕉園在子囊菌門(Ascomycota)、羅茲菌門(Rozellomycota)有明顯富集;相對于輪作蕉園和宿根蕉園,新開墾蕉園在擔子菌門(Basidiomycota)、被孢霉門(Mortierellomycota)、壺菌門(Chytridiomycota)、線蟲(Nematoda)、毛霉菌門(Mucoromycota)有明顯富集;與輪作蕉園和新開墾蕉園相比,宿根蕉園在毛霉菌門(Mucoromycota)、球囊菌門(Glomeromycota)、Anthophyta、纖毛門(Ciliophora)、綠藻門(Chlorophyta)、絲足蟲門(Cercozoa)等有明顯富集(圖4B)。
為了進一步分析宿根蕉園、輪作蕉園和新開墾蕉園各點的代表性物種,進行了指示物種分析。通過分析發現,輪作蕉園與新開墾蕉園相比,細菌類方面,小單孢菌屬(Micromonospora)、1921-2、赭黃嗜鹽囊菌(Haliangiu)、硝化螺旋菌(Nitrospira)、庫茨涅爾氏菌(Kutzneria)、小梨形菌屬(Pirellula)、中慢生根瘤菌屬(Mesorhizobium)、小單孢菌(Micromonospora)有顯著差異(圖5A);真核微生物方面,籃狀菌屬(Talaromyces)、角菌根菌屬(Ceratobasidium)、輪枝孢屬(Musicillium)有顯著差異(圖5B)。輪作蕉園與宿根蕉園相比,細菌類方面,熱酸菌屬(Acidothermus)、1921-2、庫茨涅爾氏菌(Kutzneria)存在顯著差異(圖5C);真核微生物方面,籃狀菌屬(Talaromyces)、小菇屬(Mycena)存在顯著差異(圖5D)。新開墾蕉園與宿根蕉園相比,細菌類方面,熱酸菌屬(Acidothermus)、芽胞桿菌(Bacillus)存在明顯差異(圖5E);真核微生物方面,角菌根菌屬(Ceratobasidium)存在顯著差異(圖5F)。上述結果為解析宿根蕉園連作成功的模式奠定了基礎。
2.4 ?蕉園土壤細菌和真菌的基因功能分析
為了進一步分析3種蕉園土壤特征,通過功能分析構建物種與基因的網絡,結合基因的KEGG通路信息來預測整個群落的通路情況。從細菌的功能豐度熱圖(圖6A)可以看出,宿根蕉園組的大多數代謝通路的基因豐度顯著低于輪作蕉園和新開墾蕉園,僅細菌遷移(cell mobility)一項高于另外2種蕉園,而輪作蕉園和新開墾蕉園在酶家族(enzyme families)、生物合成和次級代謝(biosynthesis of other metabolites)、轉錄(transcription)、碳代謝(carbohydrate metabolism)、膜轉運(membrane transport)以及多糖的生物合成與代謝(glycan biosynthesis and metabolism)相關的基因豐度明顯高于輪作蕉園(圖6A)。從真菌的功能豐度熱圖(圖6B)可以看出,宿根蕉園組的大多數代謝通路的基因豐度顯著高于輪作蕉園和新開墾蕉園,而新開墾蕉園又低于輪作蕉園;宿根蕉園土壤真菌從代謝通路上看,在細胞遷移(cell mobility)、細胞生長和死亡(cell growth and death)、信號傳導(signal transduction)、傳染性病害(infectious diseases)、核酸代謝(nucleotide metabolism)、蛋白折疊儲運和降解(folding, sorting and degradation)上有明顯的優勢;輪作蕉園真菌從代謝通路上看,在輔助因子和維生素代謝(metabolism of cofactors and vitamins)、其他次生代謝產物的生物合成(biosynthesis of other secondary metabolites)、氨基酸代謝(amino acid metabolism)、激素代謝方面有明顯的優勢;新開墾蕉園土壤真菌在碳代謝(carbohydrate metabolism)通路上有明顯的富集。上述結果表明,土壤真菌菌群可能是影響香蕉枯萎病數量的關鍵因子。
3 ?討論
海南是典型的熱帶亞熱帶氣候,試驗取材地點臨高皇桐是典型的紅壤,土壤酸性較強。前人研究也表明,蕉園香蕉枯萎病爆發的幾率和發病程度與土壤酸堿度有較大關系,酸性越強,香蕉枯萎病爆發幾率越高[33],蕉園病株又進一步酸化根際土壤[34],一旦陷入該種惡性循環,往往導致蕉園廢棄。宿根蕉園土壤pH顯著高于輪作蕉園和新開墾蕉園,全生育期變異范圍也最小,可能是宿根蕉園能順利運行的主要原因。全生育期蕉園土壤pH動態變化分析發現,pH在雨季下降明顯,相對應的同期土壤香蕉枯萎病孢子數目也明顯上升,表明二者有一定的關聯性。此外,新開墾蕉園和輪作蕉園pH<6,不利于多數細菌和放線菌的生長,而較低的pH更利于香蕉枯萎病菌的生長,孢子相對數目較高;而宿根蕉園pH>6,不利于香蕉枯萎病菌的生長,孢子數相對含量低。
先前研究發現,雖然不同蕉園土壤理化性質及養分含量存在很大差異,但與香蕉枯萎病的發生無直接相關性;蕉園土壤微生物數量的改變是香蕉枯萎病發病的重要特征,枯萎病危害程度為中度、重度的土壤中細菌和放線菌的數量明顯低于危害程度為輕度和無危害的土壤,危害程度為中度、重度的土壤真菌數量明顯高于危害程度為輕度和無危害的土壤[24]。通過對3種蕉園土壤菌群特征分析發現:宿根蕉園土壤的細菌和真菌無論在種類上還是在豐度上均顯著高于輪作蕉園和新開墾蕉園;宿根蕉園土壤細菌菌群活躍程度最低,輪作蕉園低于新墾蕉園;宿根蕉園土壤真菌菌群活躍程度最高,新墾蕉園低于輪作蕉園,表明真菌的活躍程度可能在維持土壤健康和抑制香蕉枯萎病爆發過程中有重要作用。鄧曉等[21]通過分析海南23個香蕉園土壤微生物數量與18個土壤因子的相關性發現,蕉園土壤5~30 cm土層可培養微生物的區系依次為細菌、放線菌、真菌;有效Fe、有效P、土壤pH分別是影響蕉園土壤真菌數量、放線菌數量和土壤微生物總數的關鍵因子。新開墾蕉園和輪作蕉園對于宿根蕉園土壤細菌和真菌的豐度問題可能還需進一步結合其他土壤因子進行分析。通過間作韭菜、假花生和覆蓋稻草處理可顯著提高香蕉根區土壤可培養細菌和放線菌數量,降低尖孢鐮刀菌數量[35]。宿根蕉園由于香蕉莖桿多堆于行間,經分解后還于蕉園,土壤有機質含量明顯高于其他2種模式,部分特征與間作和覆草類似,因此無論是土壤細菌還是真菌的菌群數量和豐度均高于輪作和新開墾蕉園。黃珍等[36]對海南省福山香蕉園香蕉枯萎病發生地和正常種植園的土壤分析發現,正常香蕉種植區的土壤樣品的細菌多樣性較為豐富,其中變形菌門、厚壁菌門、酸桿菌門為主要細菌類群;而香蕉枯萎病發生地土壤以放線菌門和厚壁菌門為主要細菌類群。皇桐、南寶與美臺3個地區蕉園土壤中主要優勢細菌可能為枯草芽胞桿菌、葡萄球菌、反芻真桿菌等[37]。正常香蕉種植區土壤樣品的細菌多樣性較為豐富,以變形菌門、厚壁菌門、酸桿菌門為主要細菌類群;而香蕉枯萎病發生地土壤以放線菌門和厚壁菌門為主[36]。通過土壤特征菌群分析發現,宿根蕉園與新開墾蕉園相比,細菌類方面,熱酸菌屬、芽胞桿菌芽胞桿菌屬存在明顯差異,真核微生物方面,角菌根菌屬存在顯著差異;宿根蕉園與輪作蕉園相比,細菌類方面,熱酸菌屬、1921-2、庫茨涅爾氏菌存在顯著差異,真核微生物方面,籃狀菌屬、小菇屬存在顯著差異。上述研究結果中,熱酸菌屬、芽胞桿菌芽胞桿菌屬等與先前報道相同,同時也發現一些新的菌屬,為未來土壤改良和拮抗菌篩選提供新方向。同時,取材時間、使用化肥和生物肥、種植模式等諸多原因均可能導致菌群特征結果有較大差異。因此相關結果還需進一步的驗證和研究。
參考文獻
胡從九. 淺析世界香蕉市場變化及趨勢[J]. 中國熱帶農業, 2020(6): 39-41, 11.
付 ?崗, 葉云峰, 杜嬋娟, 等. 香蕉枯萎病菌群體多樣性研究進展[J]. 植物檢疫, 2016, 30(2): 1-6.
曾莉莎, 林威鵬, 呂 ?順, 等. 香蕉-甘蔗輪作模式防控香蕉枯萎病的持續效果與土壤微生態機理(Ⅰ)[J]. 中國生態農業學報(中英文), 2019, 27(2): 257-266.
林威鵬, 曾莉莎, 呂 ?順, 等. 香蕉-甘蔗輪作模式防控香蕉枯萎病的持續效果與土壤微生態機理(Ⅱ)[J]. 中國生態農業學報(中英文), 2019, 27(3): 348-357
劉文清, 崔廣娟, 王 ?芳, 等. 香蕉-甘蔗輪作對土壤養分含量及酶活性的影響[J]. 廣東農業科學, 2019, 46(8): 86-96.
劇虹伶. 辣椒-香蕉輪作聯合生物有機肥減輕高發枯萎病蕉園連作障礙機制研究[D]. 海口: 海南大學, 2017.
李夢輝. 辣椒對香蕉枯萎病的防控作用效果[D]. 海口: 海南大學, 2017
耿建建. 稻稈還田淹水聯合水稻輪作對高發枯萎病蕉園土壤修復效應研究[D]. 海口: 海南大學, 2016.
辛 ?侃. 水稻—香蕉輪作并向土壤中添加有機物料防控香蕉枯萎病的研究[D]. 海口: 海南大學, 2014.
辛 ?侃, 趙 ?娜, 鄧小墾, 等. 香蕉-水稻輪作聯合添加有機物料防控香蕉枯萎病研究[J]. 植物保護, 2014, 40(6): 36-41, 52.
黃永紅. 韭菜對香蕉枯萎病的防控效果及其作用機理的研究[D]. 長沙: 湖南農業大學, 2011.
柳 ?影, 丁文娟, 曹 ?群, 等. 套種韭菜配施生物有機肥對香蕉枯萎病及土壤微生物的影響[J]. 農業環境科學學報, 2015, 34(2): 303-309.
黃永紅, 魏岳榮, 李春雨, 等. 韭菜對香蕉枯萎病菌4號生理小種的抑制作用及對香蕉枯萎病的防控研究(英文)[J]. 果樹學報, 2011, 28(5): 857-862, 943.
吳宇佳, 楊 ?春, 雷 ?菲, 等. 套種花生對香蕉園小氣候和土壤理化性質的影響[J]. 福建農業學報, 2020, 35(3): 337-343.
許林兵, 黃秉智, 肖維強. 抗枯萎病香蕉新品種‘南天黃的特征與栽培技術要點[J]. 中國熱帶農業, 2016(4): 24-27.
許林兵, 張錫炎, 李華平, 等. 抗枯萎病香蕉新品種‘南天黃選育[J]. 熱帶作物學報, 2017, 38(6): 998-1004.
王貴花. 香蕉枯萎病菌侵染巴西蕉和寶島蕉差異轉錄組分析[D]. 海口: 海南大學, 2016.
李華平, 李云鋒, 聶燕芳. 香蕉枯萎病的發生及防控研究現狀[J]. 華南農業大學學報, 2019, 40(5): 128-136.
李紹鵬, 陳文河. 厄瓜多爾香蕉產業經營經驗考察報告[J]. 熱帶農業科學, 2000(6): 42-45, 56.
周兆禧, 馬蔚紅, 李敬陽, 等. 海南宿根一代蕉高產栽培管理技術[J]. 中國熱帶農業, 2011(6): 74-76.
鄧 ?曉, 洪 ?葵, 李勤奮, 等. 海南香蕉園土壤微生物與土壤因子的關系[J]. 熱帶作物學報, 2010, 31(4): 530-535.
樊小林, 李 ?進. 堿性肥料調節香蕉園土壤酸度及防控香蕉枯萎病的效果[J]. 植物營養與肥料學報, 2014, 20(4): 938-946.
黃建鳳, 張發寶, 逄玉萬, 等. 酸性土壤改良劑與生防制劑協同防控香蕉枯萎病的效果[J]. 熱帶作物學報, 2017, 38(3): 545-550.
鄧 ?曉, 李勤奮, 侯憲文, 等. 香蕉枯萎病區土壤可培養微生物生態特征[J]. 熱帶作物學報, 2011, 32(2): 283-288.
杜 ?浩, 只佳增, 李宗鍇, 等. 我國土壤微生物菌群構建防控香蕉枯萎病研究進展[J]. 熱帶農業科學, 2020, 40(2): 90-98.
付 ?琳, 阮云澤, 沈宗專, 等. 生物有機肥對連作香蕉根際土壤可培養細菌區系的影響[J]. 南京農業大學學報, 2012, 35(6): 82-88.
齊鴻雁, 薛 ?凱, 張洪勛. 磷脂脂肪酸譜圖分析方法及其在微生物生態學領域的應用[J]. 生態學報, 2003(8): 1576-1582.
孫建波, 鄒良平, 李文彬, 等. 香蕉不同生育期根際土壤細菌群落變化研究[J]. 熱帶作物學報, 2016, 37(6): 1168-1171.
鄧 ?曉, 李勤奮, 武春媛, 等. 健康香蕉(Musa paradisiaca)植株與枯萎病患病植株根區土壤細菌多樣性的比較研究[J]. 生態環境學報, 2015, 24(3): 402-408.
漆艷香, 張 ?欣, 彭 ?軍, 等. 不同抗、感枯萎病香蕉種質根際土壤的微生物數量[J]. 江蘇農業科學, 2019, 47(13): 110-114.
景曉輝, 吳倫英, 區小玲, 等. 一種簡便分離香蕉枯萎病菌的選擇性培養基[J]. 熱帶作物學報, 2009, 30(11): 1671-1673.
韓寶坤, 杜艷華. 非無菌操作下分離尖孢鐮刀菌的培養基[J]. 植物病理學報, 2001(4): 373.
李 ?進, 張立丹, 劉 ?芳, 等. 堿性肥料對香蕉枯萎病發生及土壤微生物群落的影響[J]. 植物營養與肥料學報, 2016, 22(2): 429-436.
楊紹瓊, 張光勇, 孫寅虎, 等. 香蕉枯萎病區感病與健康植株根系附近土壤養分及土壤酶活性差異性研究[J]. 安徽農業科學, 2020, 48(4): 141-145.
王麗霞, 井 ?濤, 殷曉敏, 等. 不同種植模式對香蕉根區土壤養分及可培養微生物數量的影響[J]. 中國南方果樹, 2020, 49(4): 80-86, 89.
黃 ?珍, 譚志瓊, 阮云澤. 香蕉園土壤16S rDNA文庫分析[J]. 熱帶作物學報, 2010, 31(6): 989-993.
陳 ?波, 張錫炎, 黃 ?霄, 等. 香蕉枯萎病區土壤真菌多樣性分析[J]. 江蘇農業科學, 2013, 41(11): 354-357.
責任編輯:謝龍蓮
