牦牛KAT8基因在組織及卵泡發育過程中的表達
2021-09-10 05:46:08熊顯榮馬鴻程閔星宇
中國獸醫學報 2021年8期
海 卓,熊顯榮,馬鴻程,張 賀,閔星宇,李 鍵*
(1.西南民族大學 畜牧獸醫學院,四川 成都 610041;2.西南民族大學 動物科學國家民委重點實驗室,四川 成都 610041;3.青藏高原動物遺傳育種資源保護與利用教育部重點實驗室,四川 成都 610041)
遺傳是一種在不改變堿基遺傳序列情況下的修飾遺傳,主要包括組蛋白修飾、DNA甲基化修飾、染色質結構修飾及復雜三維基因組排列[1-2]。染色質、染色體是真核生物遺傳物質處于有絲分裂及減數分裂特定階段中的特定形式,其基本單位是由組蛋白H2A、H2B、H3和H4構成的八聚體與DNA緊密纏繞形成的核小體[3]。DNA甲基化修飾在胚胎腎臟輸尿管分支的形成、胎盤組織的發育皆具有重要作用[4-5]。近幾年組蛋白修飾逐漸被人們所熟知,它是一種由特定相關因子作用于組蛋白特定殘基位點的遺傳修飾,甲基化、乙酰化、磷酸化、糖基化、泛素化等是其主要的修飾途徑[6]。組蛋白乙酰化修飾通過組蛋白乙酰化酶(HATs)與去乙酰化酶(HDACs)調節基因轉錄后的遺傳過程,進而影響細胞的分裂分化[7]。目前發現參與修飾的HATs約有15個,根據其結構域的同源性至少可分為3個家族:GCN5與其相關的N-端乙酰基轉移蛋白、環磷腺苷效應元件結合蛋白(CBP)及其類似蛋白(p300、MoZ、YBF2、SAS3、SAS2)和MYST(Tip60)。目前組蛋白乙酰化在動植物相關基因的研究主要集中于MyST家族[8],MyST家族有組蛋白乙酰基轉移酶Tip60(KAT5)、KAT6A、KAT6B、KAT7和KAT8,且這些家族成員在組蛋白氨基酸殘基的作用位點并不相同[9-10]。
組蛋白乙酰化酶8(KAT8)又稱為MYST1或MOF,除包含C-端HAT結構域外還含有MYST結構域(存在CoA、染色質2個結合位點及C2HC型鋅指蛋白)[11]。KAT8主要作用于組蛋白H4的賴氨酸K16殘基位點(H4K16)[12]。KAT8最初發現于果蠅體內,參與X染色體遺傳物質平衡的劑量補償過程,并被命名為MOF[13]。有研究表明,KAT8是一種潛在的雌激素受體α(estrogen receptor α,ECα)抑制因子,用RNAi技術干擾其表達可促進細胞增殖[14]。KAT8與天冬氨酸重復蛋白5(aspartic acid repeat-containing protein 5,WDR 5)共同作用于雄激素受體(androgen receptor,AR)靶基因上,從而激活前列腺癌細胞中雄激素依賴基因,且敲除KAT8基因能夠顯著降低AR靶基因的表達,進而影響雄性生殖系統[15]。在人胚腎293 T細胞中,過表達KAT8不僅能夠促進H4K16發生乙酰化,還致使多種組蛋白和非組蛋白發生丙酰化修飾[16]。研究發現,KAT8基因也介導雌性卵巢發育及卵母細胞減數分裂調控[17]。植入前胚胎會因母體KTA8缺失而出現囊胚形成率降低及單個囊胚中總細胞數降低的現象,說明KAT8在雌性生殖過程中發揮著不可替代的作用[18]。
牦牛(Bosgrunniens)是一種生活在高海拔、強紫外線且低氧環境中的高原地區優勢物種,是高原地區人民生活保障的必需家畜,具有“全能家畜”、“高原之舟”的美稱。與黃牛的繁殖情況不同,牦牛是季節性發情,通常1年1胎或3年2胎,產犢率低下[19],這嚴重阻礙了牦牛產業的發展。KAT8作為一種重要的染色質組蛋白修飾因子,在牦牛中的表達水平及作用機制人們尚不清楚。為此,本研究利用RT-PCR技術對牦牛KAT8基因進行克隆和生物信息學分析,對其組織表達譜及該基因在卵母細胞成熟過程中的時序表達規律進行探究,并通過免疫組織化學方法對KAT8在不同發育時期卵泡中的定位進行研究,旨在分析KAT8理化性質及在牦牛生殖過程中的表達規律,為牦牛育種提供理論基礎。
1 材料與方法
1.1 樣品采集在秋季選擇3頭3~5周歲未妊娠的母牦牛,屠宰后立即采集心臟、胃、腎臟、肝臟、肺臟、小腸、脾臟、肌肉、子宮及卵巢組織,利用凍存管收集后置于液氮中暫做保存。采集10頭3~5周歲母牦牛卵巢,利用無菌生理鹽水沖洗干凈,隨機分為2份,一份放入4%多聚甲醛(PFA)中進行固定;另一份放入含有雙抗(青霉素、鏈霉素)的30℃生理鹽水中暫存,以上樣品于3 h內帶回實驗室。
1.2 主要試劑TRIzol RNA提取試劑購自Invitrogen公司;核酸助沉劑購自北京百泰克神武技術有限公司;PrimeScrptTMRT Reagent Kit 反轉錄試劑盒、pMD19-T載體均為TaKaRa公司產品;ChamQ Universal SYBR qPCR Master Mix試劑盒、2×Phanta?Max Master Mix(Dye Plus)由南京諾唯贊生物科技股份有限公司提供;DNA膠回收試劑盒購自康寧生命科學有限公司;DEPC、DNA Marker、感受態細胞DH5α均購自天根生化科技有限公司;Rabbit Anti-MYST1/KAT8 antibody由北京博奧森生物技術有限公司提供。
1.3 卵巢獲取及卵母細胞收集在無菌環境下,收集卵巢表面直徑為2 ~10 mm卵泡中的卵泡液,轉移至直徑為90 mm的培養皿中,在體視顯微鏡下選取顆粒細胞包裹致密、細胞無變形退化的卵丘-卵母細胞復合體(cumulus-oocyte complexes,COCs)50~70顆,用OM液清洗2~3次后于透明質酸酶中脫去顆粒細胞,用PBS洗滌3次,獲得生發泡時期(GV期)卵母細胞,放入1.5 mL EP管中備用。分別收集體外培養的第1次減數分裂中期(MⅠ)和第2次減數分裂中期(MⅡ)卵母細胞,用上述方法處理后備用。
1.4 不同時期卵母細胞及各組織cDNA的獲得采用TRIrizol法提取各樣本總RNA,用核酸濃度測定儀檢測總RNA濃度;選取D260/D280值為1.8~2.0的RNA樣本,用PrimeScrptTMRT Reagent Kit 反轉錄試劑盒反轉錄獲得各組織及不同時期卵母細胞的cDNA,收取D260/D280值為1.8~2.0的cDNA樣本,-20℃暫存,備用。
1.5 KAT8基因克隆引物和熒光定量引物的設計與合成根據GenBank中登陸的牛KAT8序列 (GenBank登錄號:XM_027528321.1),利用Primer Premier 5.0設計KAT8分段擴增引物KAT8-1和KAT8-2,其擴增片段長度預期為996,942 bp,KAT8全長預期為1 938 bp。利用NCBI中的Primer-BLAST設計KAT8基因實時熒光定量PCR引物及β-actin定量引物(GenBank登錄號:XM_027538848.1)。引物均送由生工生物工程(上海)股份有限公司合成(表1)。

表1 KAT8 PCR引物及產物長度
1.6 牦牛KAT8基因的克隆及測序以牦牛卵巢cDNA為模板,用KAT8-1和KAT8-2分段擴增KAT8基因,PCR反應體系均為2×Phanta?Max Master Mix (Dye Plus) 25.0 μL,上、下游引物(10 μmol/L)各1.0 μL,cDNA 2.0 μL(0.5 μg/L),ddH2O 補足至50 μL。PCR擴增程序均為95℃預變性 3 min;95℃變性15 s,58℃退火15 s,72℃延伸45 s,35個循環;72℃延伸5 min。分段擴增產物經2%瓊脂糖凝膠電泳檢測后,在紫外光下切取目的條帶,用膠回收試劑盒回收目的片段。將回收的2段目的片段分別與pMD19-T載體連接,連接體系為5 μL:DNA片段 2 μL,pMD19-T載體0.5 μL,solutionⅠ 2.5 μL,16℃連接過夜。將連接產物分別轉入大腸桿菌(E.coli)DH5a感受態細胞中,并將其接種于LB液體培養基中增菌2 h后,4℃環境下8 000 r/min離心15 min收集菌液。將所獲菌體分別涂布于含有100 mg/L氨芐青霉素(Amp)的選擇培養基中,于37℃恒溫培養箱中倒置培養過夜。用無菌接種環分別挑取目的菌落,接種含Amp的液體培養基中,放在搖床上過夜培養。將獲得的菌體送由生工生物工程(上海)股份有限公司測序,利用DNAMAN將目的序列拼接,獲得KAT8基因。
1.7 牦牛KAT8生物學信息學分析利用BLAST在線系統比對物種間KAT8核苷酸序列相似性,通過MAGA 7.0軟件構建其系統進化樹。將序列輸入NCBI-ORF finder(https://www.ncbi.nlm.nih.gov/orffinder)中獲得氨基酸序列。通過ExPASy在線軟件中的ProtParam (http://web.expasy.org/protparam)、ProtScale(http://web.expasy.org/protscale/)程序及Npsa-Prabi (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPS)和SWISS_MODEL預測KAT8蛋白質的理化性質、親疏水性以及該蛋白質的結構。
1.8 KAT8 mRNA的組織表達譜分析以上述反轉錄的各組織cDNA為模板,β-actin為內參基因(表1),進行實時熒光定量分析檢測KAT8 mRNA在各組織相對表達量。熒光定量PCR總反應體系為15 μL:ChamQ Universal SYBR qPCR Master Mix 7.5 μL,上、下游引物(10 μmol/L)各0.5 μL,cDNA (0.5 μg/L) 1 μL,ddH2O 5.5 μL。熒光定量反應條件:94℃ 預變性3 min;94℃ 15 s,60℃ 30 s,72℃ 35 s, 39個循環;95℃ 15 s,60℃ 1 min。每個組織檢測樣本設3個重復組。采用2-△△Ct法計算表達量。
1.9 KAT8在卵母細胞成熟及卵泡發育過程中表達規律分析以1.3中獲得的GV期、MⅠ期、MⅡ期卵母細胞cDNA為模板,利用RT-qPCR技術檢測KAT8在卵母細胞成熟過程中的表達情況,反應體系及條件參照1.8。每個樣本設置3個重復組。采用2-△△Ct法計算表達量。
采用免疫組織化學染色法定位分析KAT8在卵巢中的表達規律。將1.2中已固定的卵巢組織用PBS沖洗干凈后進行石蠟切片,按照Rabbit Anti-MYST1/KAT8 antibody說明書稀釋(1∶100),4℃濕潤條件下孵育過夜。TBST反復清洗3次,每次5 min,滴加過氧化物偶聯鼠兔雙標二抗,室溫孵育30 min。TBST清洗3次,每次5 min,避光環境下DAB顯色液處理15 min;PBS清洗2次,每次5 min,蘇木精復染3 min后,進行藍化、脫水及中性樹脂封片鏡檢。
1.10 生物統計學分析采用SPSS 22.0統計軟件的單因素方差分析(ONE-WAY ANOVA)法分析各個組織及不同時期卵母細胞中KAT8相對表達量的差異性。免疫組織切片利用激光共聚焦進行觀察拍照,隨機選取各發育時期的卵泡。
2 結果
2.1 牦牛KAT8基因的克隆及序列分析以卵巢cDNA為模板,利用分段擴增技術擴增到約996,942 bp片段(圖1)。分段擴增產物測序后,通過DNAMAN拼接獲得1 938 bp 的KAT8基因,其CDS全長為1 377 bp,共編碼458個氨基酸。
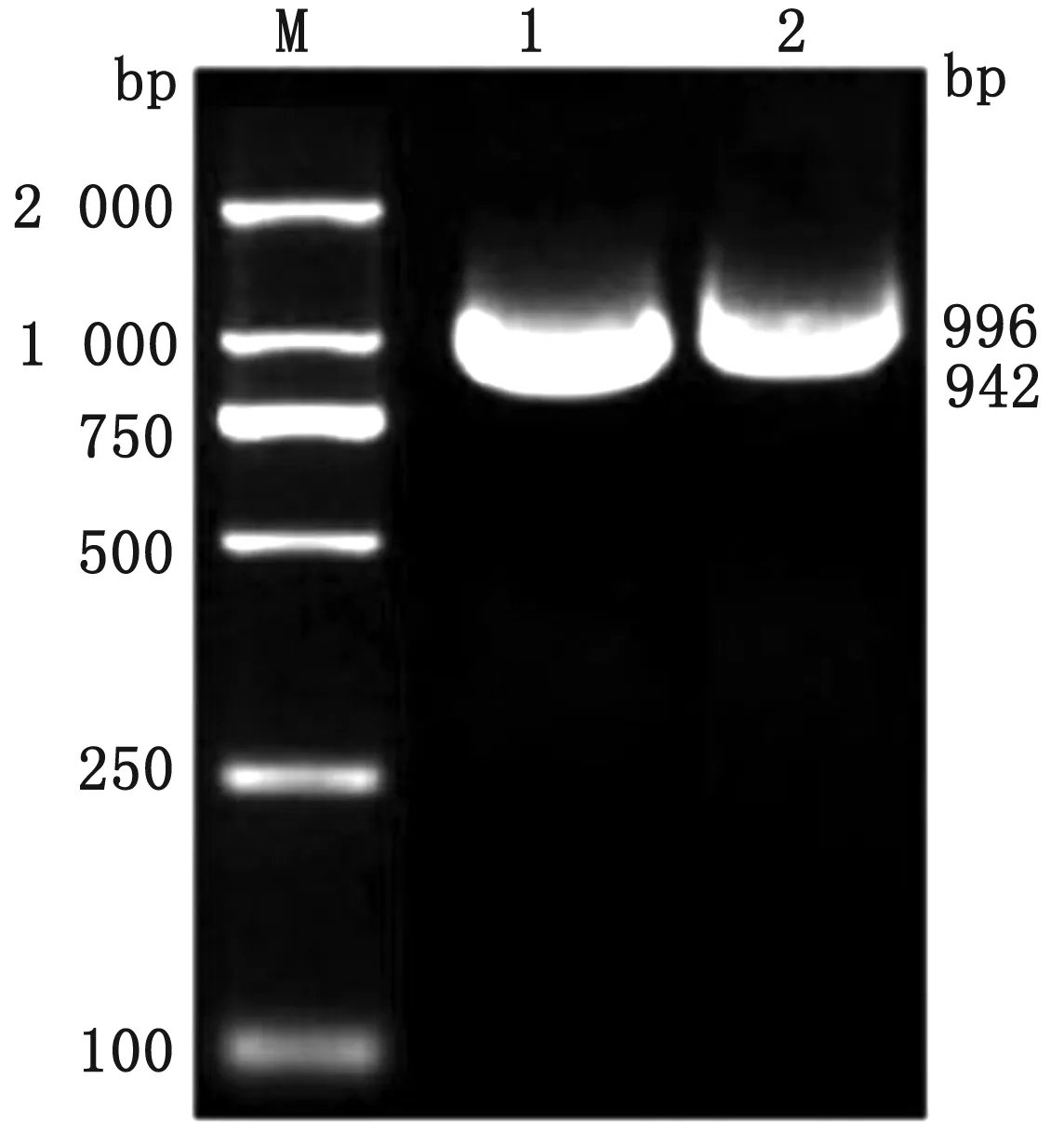
M.DL2000 DNA Marker;1,2.KAT8-2和KAT8-1擴增產物
2.2 牦牛KAT8蛋白相關生物信息學分析通過BLAST比對核苷酸發現牦牛KAT8基因與黃牛、野牛、綿羊和山羊的相似性較高,分別為99.64%,99.27%,98.33%,98.04%;與小鼠、馬和家兔核苷酸相似性較低,分別為88.67%,90.46%,91.65%(圖2A)。利用MAGA 7.0構建系統進化樹,結果顯示(圖2B),牦牛與黃牛、野牛、山羊和綿羊親源關系最近,且與核苷酸相似性比對結果一致;說明KAT8在物種進化過程中高度保守。通過ExPASy在線工具預測KAT8蛋白質相對分子質量為52.44 kDa,分子式為C2366H3671N633O690S13,共有原子數7 373個,其理論等電點為8.48,脂肪指數為77.90,不穩定指數為41.26,推測該蛋白質為不穩定蛋白,且在哺乳動物中該氨基酸預估半衰期為30 h。KAT8蛋白中含量較高的氨基酸有賴氨酸(Lys)、谷氨酸(Glu)和亮氨酸(Leu),分別為9.4%,8.3%,8.1%;含量較低的氨基酸有半胱氨酸(Cys)、色氨酸(Trp)和甲硫氨酸(Met),分別為2.0%,2.0%,0.9%;帶負電荷殘基的天冬氨酸和谷氨酸(Asp+Glu)有59個,帶正電荷殘基的精氨酸、賴氨酸和組氨酸(Arg+Lys+His)有77個。蛋白質親疏水性分析,異亮氨酸(ILe)親水性4.500,精氨酸(Arg)親水性-4.500,且該蛋白質中大多數氨基酸殘基親水性數值小于零,最終總親疏水性平均值-0.584,因此推測KAT8為親水性蛋白質。蛋白質二級結構預測結果顯示,牦牛KAT8蛋白中α螺旋占37.25%,延伸鏈占17.43%,無規則卷曲占45.10%,無β-轉角(圖3)。SWISS_MODEL預測三級結構證實,KAT8蛋白三級結構含有ɑ螺旋、無規則卷曲和延伸鏈,且結果顯示延伸鏈主要位于ɑ螺旋和無規則卷曲結構之間(圖4)。
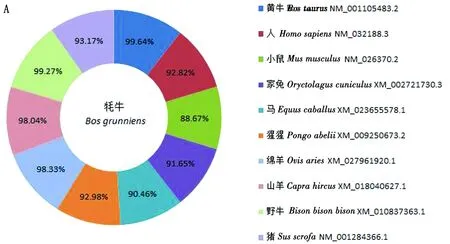
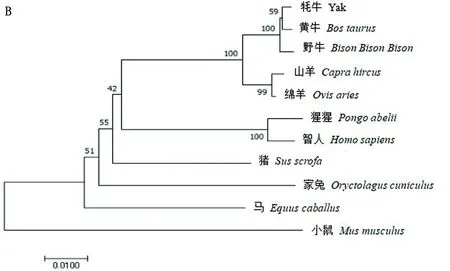
A.牦牛KAT8核苷酸序列相似性分析;B.不同物種間牦牛KAT8蛋白系統進化樹分析
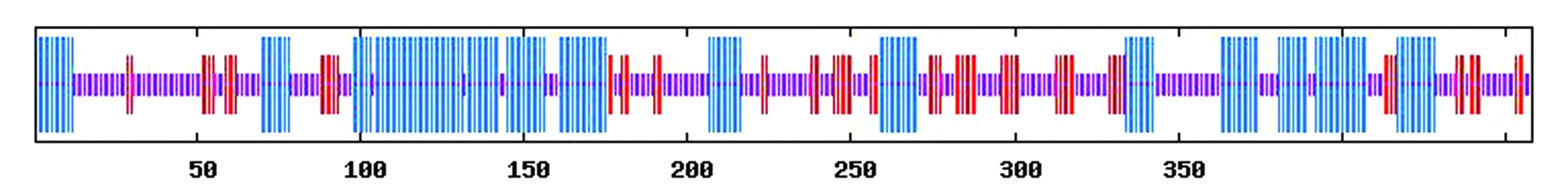
紫色指示無規則卷曲(Cc);藍色指示α螺旋(Hh);紅色指示延伸鏈(Ee)
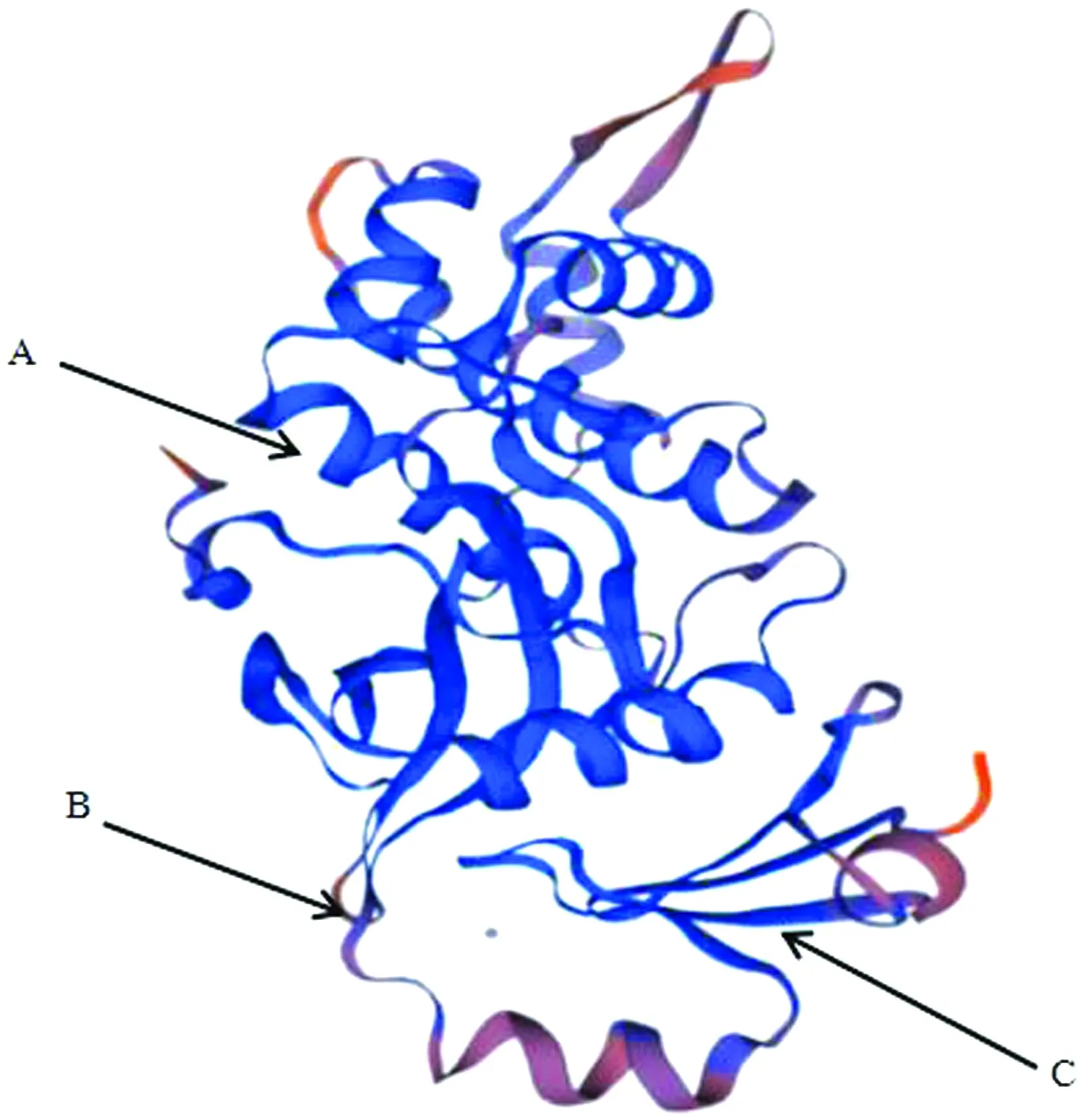
A.α螺旋;B.無規則卷曲;C.延伸鏈
2.3 牦牛KAT8 mRNA組織表達譜分析以β-actin為內參基因,利用RT-PCR技術分析各組織中KAT8 mRNA相對表達量,并繪制組織表達譜(圖5)。結果顯示,KAT8在牦牛各組織中廣譜表達,在肌肉、心臟和卵巢中相對表達量較高,極顯著高于其他組織(P<0.01);在肺臟、脾臟和胃中相對表達量較低,其中肌肉和心臟無顯著性差異,但顯著高于卵巢中KAT8 mRNA表達量(P<0.05)。
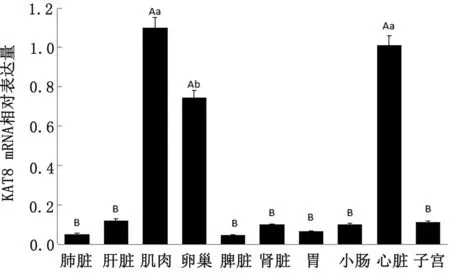
注:相同字母代表差異不顯著(P>0.05);不同小寫字母表示差異顯著(P<0.05);不同大寫字母表示差異極顯著(P<0.01)
2.4 KAT8在牦牛卵泡發育過程中的表達規律及定位分析采用免疫組織化學方法檢測,結果表明KAT8在牦牛各發育時期卵泡中均有表達,表達產物呈褐色或棕褐色(圖6)。由圖6F可知KAT8主要定位于顆粒細胞,而卵泡膜細胞中相對較少。
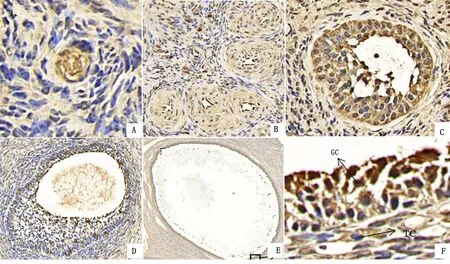
A.原始卵泡(100×);B.初級卵泡(80×);C.次級卵泡(80×);D.三級卵泡(50×);E.優勢卵泡(50×);F.局部放大(100×)。GC.顆粒細胞;TC.膜細胞
2.5 牦牛KAT8 mRNA在卵母細胞成熟過程中表達規律分析以β-actin作為內參基因,利用RT-PCR方法分析KAT8 mRNA在卵母細胞成熟過程中的時序表達規律。結果顯示,在卵母細胞減數分裂各時期KAT8均有表達,生發泡期(GV)相對表達量較高,MⅠ期次之,MⅡ期最低,但無顯著性差異(P>0.05)(圖7)。
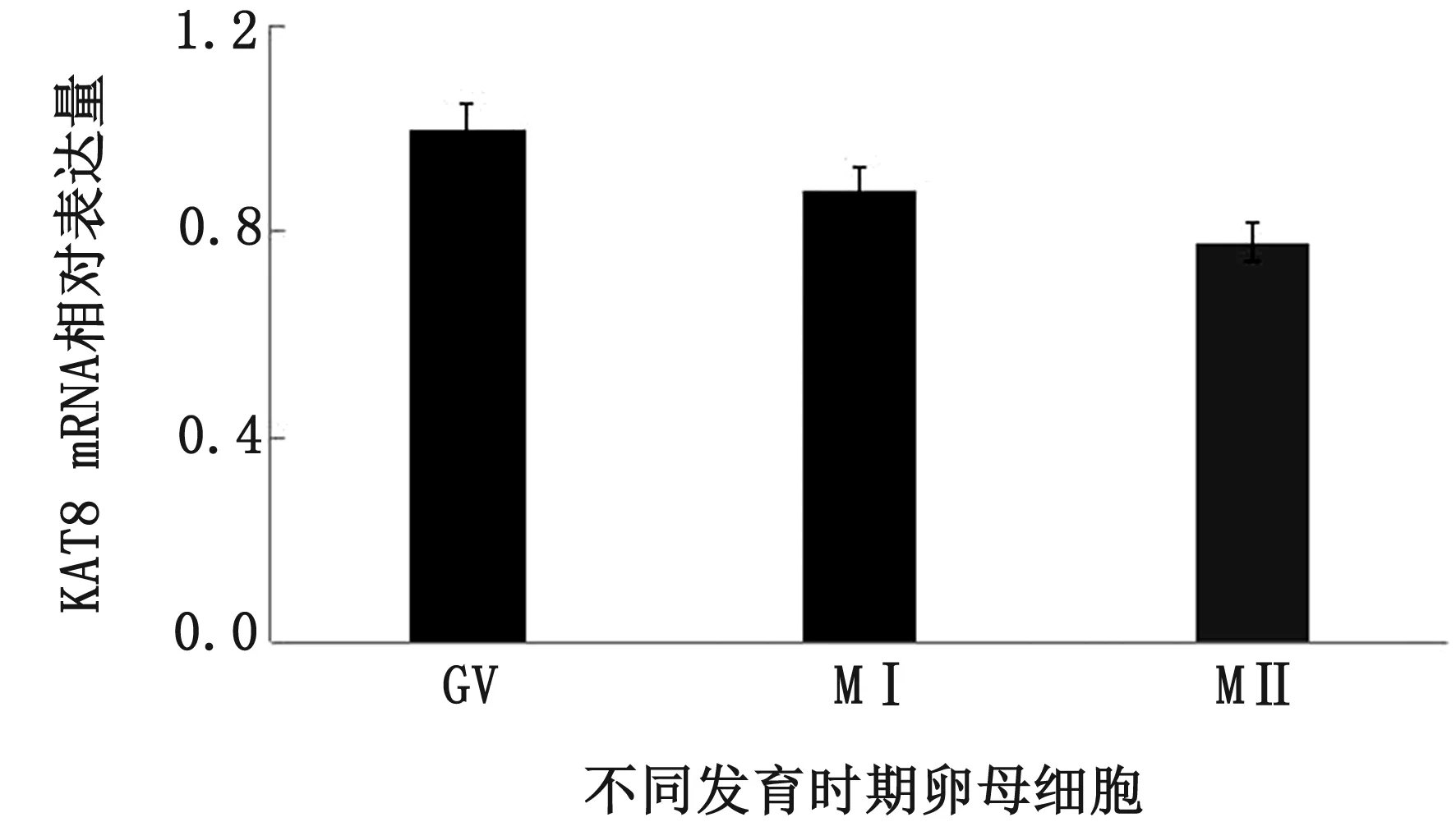
圖7 牦牛卵母細胞成熟過程中KAT8 mRNA表達規律
3 討論
目前,有關KAT8的研究主要集中在免疫及造血功能方面[20-21],然而關于其在卵巢、卵泡發育過程中的表達及作用的研究較少[17]。因此,探討KAT8在牦牛生殖過程中的表達模式及分子機制尤為重要。本試驗利用RT-PCR技術成功克隆出牦牛KAT8基因CDS序列。通過核苷酸相似性分析發現,牦牛、黃牛和野牛KAT8核苷酸序列高度吻合。綜上表明,KAT8基因在物種進化過程中高度保守。
研究可知,KAT8在動物不同組織中的作用不盡相同[19-22],在牦牛各個組織中的相對表達量及在生殖器官及生殖細胞發育過程中的調節機制仍未可知。本研究利用RT-qPCR技術檢測牦牛各組織中KAT8 mRNA相對表達量,結果表明,KAT8在牦牛各組織中廣譜表達,其中肌肉、卵巢和心臟中表達量較高。有報道已證實KAT8(MOF)對抑制心肌肥大具有關鍵性作用,與過氧化氫酶和錳超氧化物歧化酶(MnSOD)具有協同作用,可阻斷心臟橫向動脈縮窄誘導的活性氧基因(ROS)及其下游的c-Raf-MEK-ERK通路,從而抑制心肌肥大[8]。因此,推測牦牛耐低氧特性與其心臟中KAT8 mRNA高表達有關,具體作用機制有待進一步研究。YIN等[17]利用基因編輯技術敲除小鼠卵巢KAT8基因,探究母體卵巢KAT8表達對后續生殖過程的影響,結果發現,敲除鼠卵巢形態與野生型小鼠卵巢形態差異較大,體脂率降低77.4%,且出現了卵泡減少及卵泡表面直徑減小的現象。有研究發現,在人正常卵巢組織中KAT8 mRNA在相對表達量顯著高于卵巢癌組織[23]。本研究證實,KAT8 mRNA在牦牛卵巢中表達量較高。綜上結果表明,KAT8參與卵巢發育,其缺失會嚴重影響卵巢的正常發育。
為進一步探究KAT8在牦牛卵母細胞減數分裂進程的表達規律,本研究利用RT-qPCR技術檢測KAT8 mRNA在牦牛不同時期卵母細胞中的相對表達量,發現GV、MⅠ和MⅡ期卵母細胞表達水平呈下降趨勢,這與YIN等[17]對小鼠卵母細胞的研究結果一致。有研究證實,在卵母細胞成熟過程中組蛋白殘基逐漸進行乙酰化修飾,GV期乙酰化水平最高,隨后降低,同時去乙酰化酶水平隨之升高[22]。結合組蛋白去乙酰化酶1(HDAC1)及組蛋白去乙酰化酶8(HDAC8)在GV期的表達結果推測卵母細胞GV期較高的乙酰化水平抑制去乙酰化修飾作用[19,24]。有研究表明,KAT8在小鼠卵母細胞不同時期中的表達水平存在顯著性差異,GV期顯著高于MⅠ和MⅡ期[17],但本研究中牦牛3個時期卵母細胞表達水平無顯著性差異。本試驗對核苷酸序列相似性分析發現,牦牛KAT8與小鼠相似性最低,推測該基因在牦牛和小鼠中出現的差異可能與物種特異性及牦牛特殊生存環境有關,具體原因有待進行深入研究。
最后,本試驗采用免疫組化對KAT8在牦牛卵巢不同發育時期卵泡的表達定位進行探究,發現KAT8在卵泡各發育期均有表達,且主要定位于卵泡顆粒細胞中。由人正常卵巢及患有癌癥的卵巢組織免疫組化結果可知,KAT8在卵巢多種細胞中均表達,但在不同發育時期的正常及患癌卵泡顆粒細胞中的表達量相對其他細胞明顯升高[23]。目前KAT8在經濟動物生殖過程中的研究尚未見報道。研究KAT8在牦牛不同發育時期卵泡中的表達規律,對進一步探究該基因對雌性牦牛生殖相關的作用機制提供理論依據。
