抗魚類病原菌的放線菌分離鑒定及其活性研究
2021-09-09 01:07:22馬永紅劉思蓓余子全
激光生物學報 2021年4期
馬永紅,朱 麗,劉思蓓,余子全
(湖南師范大學生命科學學院,淡水魚類發育生物學國家重點實驗室,微生物分子生物學湖南省重點實驗室,長沙 410081)
殺鮭氣單胞菌(Aeromonas salmonicida)是一種嗜冷無動力的氣單胞菌, 廣泛分布于淡水與海水環境中,是一種不能運動、兼性厭氧的革蘭氏陰性菌。其最適生長溫度為22~25℃,35℃以上不生長,是一種條件致病菌,能引起癤瘡病,對魚類的存活有著顯著的影響,曾經給全球水產養殖業造成了巨大的困擾[1]。
癤瘡病是由殺鮭氣單胞菌引起的一種高發性的魚類疾病,影響著世界范圍內的水產養殖,其特點是發病率高,死亡率高[2]。病魚的病癥主要表現為魚體內腹水,體表多處潰爛[3]。在癤瘡病等魚類細菌性疾病的預防與治療中,不規范頻繁地使用化學藥物導致病原體耐藥性增強,水體嚴重污染,從而造成水產疾病難以控制,同時也引發了一系列食品安全問題。因此,利用無毒無污染的微生物及其次級代謝產物作為微生物漁藥來防控水產疾病無疑成為了魚病研究的熱點[4]。
放線菌能產生多種具有獨特作用機制的活性天然產物,同時已有部分研究表明放線菌也能作為一種益生菌用于水產病害的防治。如Babu等[5]在蝦養殖區域的淤泥中分離到多株對白斑綜合征病毒(white spot syndrome virus,WSSV)具有活性的放線菌,并且發現這些放線菌在蝦養殖系統中具有潛在的生物修復作用。拮抗作用是細菌或其他微生物相互作用的方式之一,是指一種微生物在生長過程中通過某種代謝產物或改變生活環境來抑制其他微生物的生長,甚至殺死它們的現象。利用拮抗性能強的菌株抑制病原菌的生長、繁殖或致病性是生物防治的手段之一[6]。疾病防控的重點應該是預防。拮抗菌可以作為抑制病原菌的生物控制劑,能夠減少對化學藥品的使用和依賴,比治療更具經濟效益[7]。因此,本研究主要以殺鮭氣單胞菌為指示菌,從特殊生境下取土樣對該致病菌進行拮抗菌的篩選以及活性研究,為魚類癤瘡病的防治提供新的策略和方法。
1 材料與方法
1.1 鏈霉菌I6分離的材料來源
銀川市賀蘭山平均海拔2 000 m以上,是溫帶草原與荒漠的交界線。該地區植被垂直分布,自然環境較為復雜,土壤類型較多。本研究從該地區山地草甸取土樣,從中分離到鏈霉菌I6。
1.2 鏈霉菌I6的形態學觀察
將無菌蓋玻片以45°左右的角度并排插入高氏固體平板培養基中,插入深度為蓋玻片的1/3左右;用無菌竹簽挑取鏈霉菌I6孢子,接種于蓋玻片與高氏培養基的交界線上,在28℃下培養3 d左右;在相差顯微鏡與掃描電鏡下觀察鏈霉菌I6菌絲與孢子的形態特征以及排列情況等[8]。
1.3 鏈霉菌I6 16S rDNA的擴增測序及系統發育樹的構建
使用細菌基因組DNA快速抽提試劑盒提取鏈霉菌I6的全基因組[9],以基因組DNA為模板,用通用引物擴增目的基因。引物序列:27F(5'-AGAGTTTGATCCTGGCTCAG-3')、1492R(5'-ACGGCTACCTTGTTACGACTT-3')。序列預期長度約為1 500 bp。PCR反應體系:DNA模板2.5 μL(放線菌I6基因組稀釋10倍)、上游引物1.0 μL、下游引物1.0 μL、dNTP Mixture(2.5 mmol/L)4.0 μL、10×Ex Taq Buffer 5.0 μL、Ta-KaRa Ex Taq(5 U/μL)1.0 μL、無菌超純水35.5 μL,總體積50.0 μL。PCR反應條件:94℃預變性5 min,94℃變性30 s,58℃退火30 s,72℃延伸2 min,30個循環,72℃補平10 min,16℃保存。電泳檢測PCR擴增結果[10]。用膠回收試劑盒純化PCR產物,并送至上海生物工程有限公司進行測序[11]。將測序獲得的鏈霉菌I6的16S rRNA測序結果上傳至NCBI(https://www.ncbi.nlm.nih.gov/),利用BLAST和GenBank數據庫中的序列進行同源性分析。根據比對結果,在GenBank中檢索獲得標準菌株的16S rDNA序列,使用MEGA7.0.26軟件并采用鄰接法構建系統發育樹。
1.4 鏈霉菌I6活性產物分布的檢測
刮取適量鏈霉菌I6孢子至AM6培養基,置于28℃、200 r/min搖床發酵5 d;收集發酵液并離心(8 000 r/min,8 min),分別收集鏈霉菌I6的發酵上清液與細菌沉淀。菌體沉淀用無菌水洗3次,之后加入少量AM6培養基重懸、超聲破碎菌體,最后使用無菌濾頭將發酵上清液與沉淀破碎液過濾后進行抑菌試驗,測定抑菌圈的大小。
1.5 鏈霉菌I6發酵條件的優化
為了提高鏈霉菌I6的抗菌活性物質產量,本研究對鏈霉菌I6的發酵條件進行了優化。首先,刮取高氏平板上培養好的鏈霉菌I6孢子至胰蛋白胨大豆肉湯(trypticase soy broth,TSB)培養基中,置于28℃、200 r/min搖床活化2 d,然后再按5%的接種量將種子液分別接入7種不同的發酵培養基(表1)中培養5 d;取出培養后的發酵上清液,以殺鮭氣單胞菌為指示菌,未接菌的發酵培養基做為對照進行抑菌試驗,篩選活性最強的培養基。接下來將鏈霉菌I6接種至AM3-1最適發酵培養基中,置于28℃,200 r/min搖床培養,在培養的第1~12天,每天取發酵上清液(以殺鮭氣單胞菌為指示菌,未接菌的AM3-1培養基做對照)進行1次抑菌試驗,測定不同培養時間對鏈霉菌I6產抗菌物質的影響。試驗每組設3個平行,重復3次。最后,將最適發酵培養基AM3-1的初始pH分別調節至4、5、6、7、8、9、10,以5%的接種量將鏈霉菌I6的種子液分別轉接至不同初始pH的發酵培養基中,置于28℃、200 r/min搖床培養5 d;取各組鏈霉菌I6的發酵上清液,以殺鮭氣單胞菌為指示菌,未接菌的AM3-1培養基做為對照進行抑菌試驗,測定不同的初始pH對鏈霉菌I6產抗菌物質的影響,試驗每組設3個平行,重復3次。

表1 發酵培養基的類型及組成Tab. 1 Type and composition of fermentation medium
1.6 鏈霉菌I6生理生化特征的檢測
根據《鏈霉菌鑒定手冊》中的方法配制各類生理生化培養基,在無菌超凈工作臺內用已滅菌的竹簽挑取少量鏈霉菌I6孢子接種至各類生理生化培養基中,按照該手冊中的檢測方法進行各類生理生化檢測[12]。
1.7 鏈霉菌I6發酵液對殺鮭氣單胞菌的最小抑菌濃度(MIC)測定
通過梯度稀釋法對鏈霉菌I6的發酵上清液進行稀釋,得到不同濃度的稀釋液,并以未接菌的AM3-1培養基作為對照,每個樣品中加入10.0 μL(1×107CFU/mL)殺鮭氣單胞菌的菌液,于30℃過夜培養。觀察菌株的生長情況,以肉眼觀察到沒有殺鮭氣單胞菌生長的發酵液濃度設定為最小抑菌濃度(minimum inhibitory concentration,MIC)。觀察結束后,取每個濃度組的菌懸液100.0 μL至溶菌肉湯(lysogeny broth,LB)固體平板上涂布均勻,30℃過夜培養,觀察平板上殺鮭氣單胞菌的生長情況,平板內無殺鮭氣單胞菌生長的發酵液濃度設定為最小殺菌濃度(minimum bactericidal concentration,MBC)。
1.8 慶大霉素與諾氟沙星標準曲線的繪制及鏈霉菌I6發酵液效價的測定
選用常用來防治魚類病害的慶大霉素與諾氟沙星抗生素作為標準品來衡量鏈霉菌I6發酵液的效價。稱取2 mg慶大霉素與2 mg諾氟沙星抗生素分別溶解于1 mL無菌水中,配成2 mg/mL的慶大霉素母液與2 mg/mL的諾氟沙星母液,并在超凈工作臺內用無菌濾頭過濾,分別配置成1 000.00、500.00、250.00、125.00、62.50、31.25、15.63、7.81 μg/mL的慶大霉素溶液與諾氟沙星溶液。
以殺鮭氣單胞菌作為指示菌,將其涂布在LB平板上吹干;在涂有病原菌的板上打6個直徑為7 mm的孔(圖1a),分為S和K 2組孔,分別在不同的平板上加入50.0 μL不同濃度的慶大霉素與諾氟沙星抗生素以及已過濾的鏈霉菌I6的發酵上清液到S孔中,每個濃度重復3次,K孔中加入相同濃度的抗生素標準品,30℃過夜培養,觀察并計算出各濃度組的抑菌圈直徑的平均值。
將所有平板中K孔的濃度抑菌圈直徑的平均值減去每個平板上K孔的濃度抑菌圈直徑的平均值獲得每個平板的校正值。根據每個平板的校正值對各組濃度慶大霉素與諾氟沙星抑菌圈的直徑及鏈霉菌I6發酵上清液抑菌圈的直徑進行校正,得到校正值。分別以慶大霉素與諾氟沙星濃度的對數值為橫坐標,殺鮭氣單胞菌抑菌圈直徑校正值為縱坐標,繪制慶大霉素與諾氟沙星標準曲線(圖1b、1c)。然后將鏈霉菌I6發酵液校正的抑菌圈直徑數值代入標準品的回歸方程中算出所對應的標準品的濃度,得到發酵液的效價。

圖1 抗生素滴加位置及抗生素對殺鮭氣單胞菌的標準曲線Fig. 1 Antibiotic dropping position and standard curve of antibiotic against Aeromonas salmonicida(a)抗生素的滴加位置;(b)諾氟沙星對殺鮭氣單胞菌的標準曲線;(c)慶大霉素對殺鮭氣單胞菌的標準曲線。(a) Antibiotic dropping position; (b) Standard curve of norfloxacin against Aeromonas salmonicida; (c) Standard curve of gentamicin against Aeromonas salmonicida.
1.9 鏈霉菌I6發酵液對魚類的安全性評估
鏈霉菌I6發酵液對多種魚類病原菌具有抑制作用,尤其對殺鮭氣單胞菌具有非常強的抑制活性。為了將鏈霉菌I6應用到魚類養殖的疾病防控中,本研究對鏈霉菌I6的次級代謝產物進行毒性檢測,通過體外反應評估鏈霉菌I6發酵液對魚類細胞的安全性。將草魚肝臟細胞L8824培養至對數期后,在顯微鏡下觀察草魚肝臟細胞的生長狀態。將生長狀況較好的對數期草魚肝臟細胞的細胞密度稀釋為1×105個/mL,取100.0 μL加入96孔板中,放置在30℃細胞培養箱中培養12 h,隨后分別加入10.0 μL的鏈霉菌I6發酵液、未接種的AM3-1培養基、殺鮭氣單胞菌,置于30℃細胞培養箱培養12 h和24 h后,顯微鏡下觀察草魚肝臟細胞的形態。
2 結果與分析
2.1 鏈霉菌I6的鑒定
鏈霉菌I6的菌落呈圓形扁平狀且較為干燥,中間凸出,氣生菌絲為淡粉色,基內菌絲為淡黃色并伴有黃色可溶性色素產生。用顯微鏡觀察鏈霉菌I6,發現其營養菌絲沒有隔膜、不斷裂,菌絲細長且分支多而短,孢子絲呈螺旋狀。在掃描電鏡下可以看出,鏈霉菌I6孢子絲呈緊密螺旋狀(5~8圈),孢子呈扁平橢圓狀,中央有凹陷,并且孢子表面有褶皺(圖2、3)。

圖2 鏈霉菌I6的培養特征及相差顯微鏡觀察Fig. 2 The phase-contrast microscopy and culture characteristics of Strptomyces sp. I6(a)鏈霉菌I6的培養特征;(b)鏈霉菌I6的相差顯微鏡觀察。(a) Culture characteristics of Strptomyces sp. I6; (b) The phase-contrast microscopy of Strptomyces sp. I6.
2.2 鏈霉菌I6的16S rDNA擴增及系統發育分析
鏈霉菌I6的全基因組DNA提取凝膠電泳圖中條帶清晰。該菌用16S rDNA的通用引物進行PCR擴增獲得長度約為1 500 bp的特異性條帶。將16S rDNA擴增測序結果進行BLAST比對分析后,選擇相似度高的不同物種的同源序列,以FASTA格式下載。使用MEGA7.0.26軟件,運用鄰接法構建鏈霉菌I6的系統發育樹,結果顯示,鏈霉菌I6單獨為一支,與Streptomyces triticistrain NEAU A3和Streptomyces bikiniensisstrain WANG1的親緣關系較近(圖4)。但將鏈霉菌I6的生理生化特征與Streptomyces triticistrain NEAU A3和Streptomyces bikiniensisstrain WANG1的生理生化特征相比較后發現,其生理生化特征與這兩株菌稍有差異[13-14](表2),推測鏈霉菌I6可能為一株新的鏈霉菌。

表2 鏈霉菌I6與菌株Streptomyces tritici strain NEAU A3和Streptomyces bikiniensis strain WANG1的生理生化比較Tab. 2 Physiological and biochemical comparison of Streptomyces sp. I6 and Streptomyces tritici strain NEAU A3 and Streptomyces bikiniensis strain WANG1
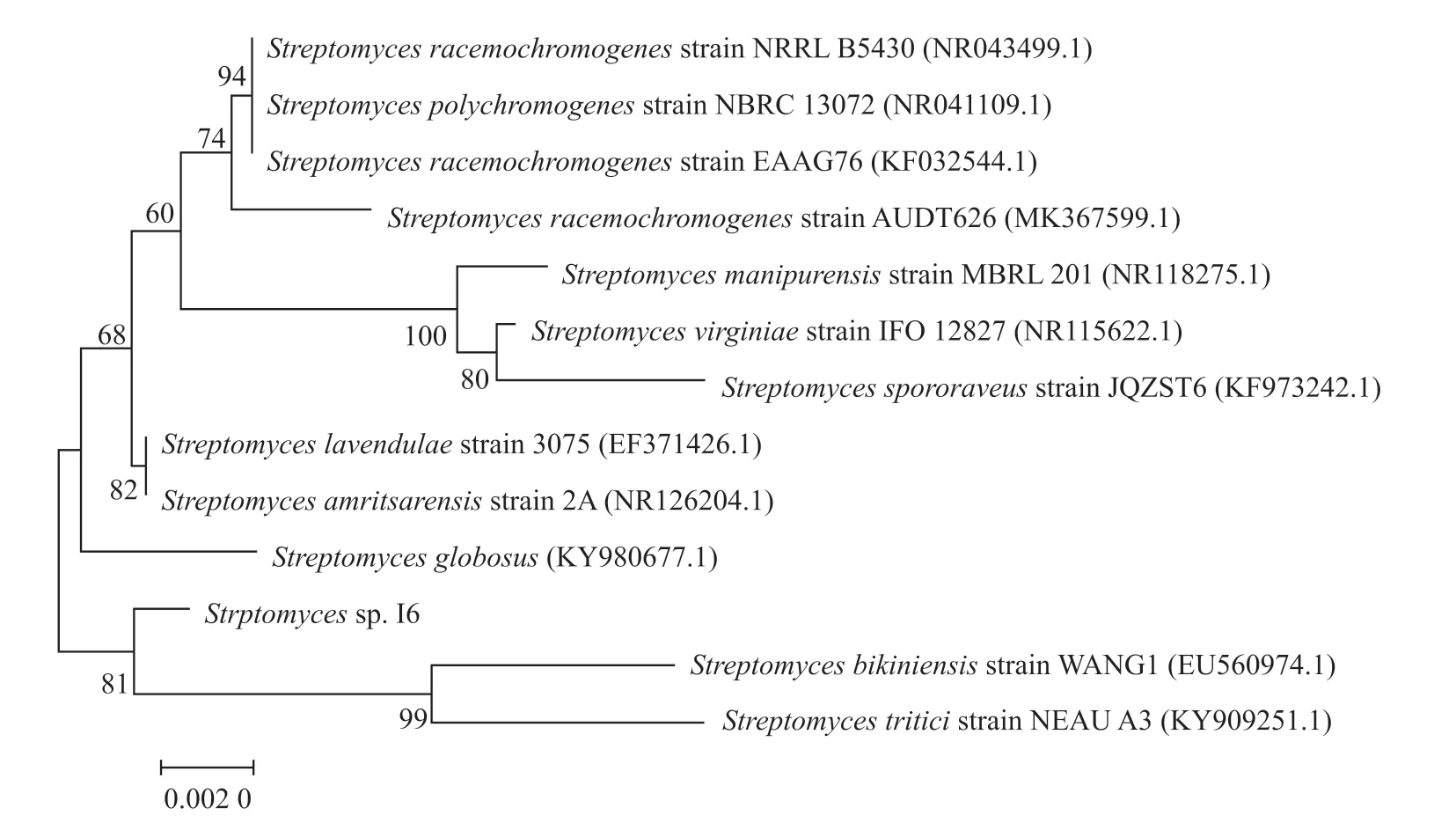
圖4 鏈霉菌I6的系統發育樹Fig. 4 Phylogenetic tree of Strptomyces sp. I6

圖3 鏈霉菌I6掃描電鏡觀察Fig. 3 The scanning electron microscope of Strptomyces sp. I6(a)鏈霉菌I6;(b)鏈霉菌I6孢子。(a) Strptomyces sp. I6; (b) Spore of Strptomyces sp. I6.
2.3 鏈霉菌I6發酵條件的優化
首先將鏈霉菌I6分別接入7種培養基中發酵,取樣檢測發酵上清液對殺鮭氣單胞菌的抑制作用,發現在AM3-1、AM6、AM24-6、AM5以及AM1培養基中能產生抗菌物質,在AM4與S培養基中不能產生抗菌物質,且AM3-1發酵液的抑菌圈直徑比其他培養基抑菌圈直徑大5.0~10.0 mm,其中比AM1培養基大10.0 mm左右,比AM6培養基大5.0 mm左右,故確定AM3-1培養基為鏈霉菌I6的最佳發酵培養基(圖5a)。接下來用最適培養基AM3-1發酵鏈霉菌I6,分別在不同的時間段進行取樣,檢測發酵上清液對殺鮭氣單胞菌的抑制作用。結果表明,活性物質在發酵后開始產生,第5天的發酵液對殺鮭氣單胞菌的抑菌作用最強,抑菌圈直徑達到25.0 mm左右,說明活性物質產量在第5天時達到頂峰,第5天后隨著發酵時間的增加,發酵液中的活性物質的產量稍有降低,但基本保持穩定,故確定鏈霉菌I6的最佳發酵時長為5 d(圖5b)。最后以5%的接種量將鏈霉菌I6的種子液分別轉接至不同初始pH的最適發酵培養基AM3-1中,發酵5 d后,取樣檢測發酵上清液對殺鮭氣單胞菌的抑制作用。結果表明,發酵液初始pH對鏈霉菌I6活性化合物的生產有明顯的影響。當pH為4時,鏈霉菌I6的發酵液內基本無抗菌化合物的產生,抑菌圈直徑為0;隨著發酵液初始pH從4增大到7~8時,抑菌圈直徑從無逐漸增大到25.0 mm左右,表明pH為7~8時活性化合物的產量達到最大;當發酵液初始pH繼續增大時,抑菌圈直徑逐漸變小;當pH為10時,抑菌圈直徑下降到10.0 mm左右,故確定鏈霉菌I6的最適pH為7~8。

圖5 鏈霉菌I6發酵條件的優化Fig. 5 The fermentation conditions of Streptomyces sp. I6 were optimized(a)不同發酵培養基對鏈霉菌I6產活性化合物的影響。字母表示顯著性差異;(b)不同發酵時間對鏈霉菌I6產活性化合物的影響;(c)發酵初始pH對鏈霉菌I6產活性化合物的影響。(a) Effects of different fermentation media on active compounds produced by Streptomyces sp. I6. Letters indicate significant differences; (b) Effects of different fermentation time on active compounds produced by Streptomyces sp. I6; (c) Effects of initial pH of fermentation broth on active compounds produced by Streptomyces sp. I6.
2.4 鏈霉菌I6抗魚類病原菌活性物質的研究
為了探究鏈霉菌I6的抗菌活性物質的分布,本研究分別用發酵上清液與沉淀做了抗殺鮭氣單胞菌的活性試驗,研究表明,鏈霉菌I6的發酵上清液對殺鮭氣單胞菌具有很強的抑菌活性(圖6)。鏈霉菌I6發酵液對殺鮭氣單胞菌的MIC為稀釋16倍后的發酵液(表3)。鏈霉菌I6發酵上清液對殺鮭氣單胞菌的校正抑菌圈直徑為25.3 mm。將此數值代入慶大霉素和諾氟沙星標準品的回歸曲線方程中,得出鏈霉菌I6發酵上清液分別相當于慶大霉素與諾氟沙星為7.67 μg/mL與665.00 μg/mL的效價。接下來評估鏈霉菌I6發酵液對草魚肝臟細胞的安全性,發現殺鮭氣單胞菌與草魚肝臟細胞共孵育12 h和24 h后細胞嚴重變形,所有細胞全部死亡。鏈霉菌I6發酵液與草魚肝臟細胞共孵育12 h和24 h后,草魚肝臟細胞未出現壞死、病變等情況,生長狀況與對照組一致,說明鏈霉菌I6的代謝產物對草魚肝臟細胞無毒性(圖7)。

圖7 鏈霉菌I6發酵上清液對草魚肝臟細胞的影響Fig. 7 Effects of supernatant of Streptomyces sp. I6 on liver cells of grass carpCK1:AM3-1發酵培養基處理;I6:鏈霉菌I6發酵液處理;CK2:殺鮭氣單胞菌處理。CK1: AM3-1 fermentation medium treatment; I6: Streptomyces sp. I6 fermentation broth treatment; CK2: Aeromonas salmonicida treatment.
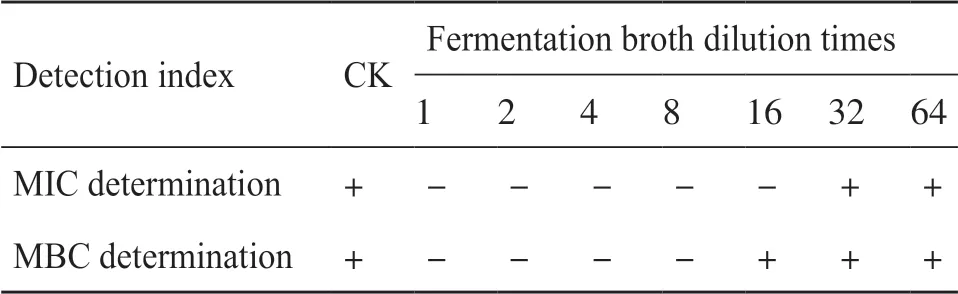
表3 鏈霉菌I6發酵液對殺鮭氣單胞菌MIC與MBC值的檢測Tab. 3 Detection of MIC and MBC values of Streptomyces sp. I6 fermentation broth against Aeromonas salmonicida

圖6 鏈霉菌I6活性物質的分布Fig. 6 Active substance distribution of Streptomyces sp. I61:鏈霉菌I6發酵上清液;2:菌體破碎重懸液;3:AM6培養基。1: Streptomyces sp. I6 fermentation supernatant; 2: Bacterial sedimentation and suspension; 3: AM6 medium.
3 討論
鏈霉菌是微生物的重要類型,廣泛存在于各種生態環境中,在陸地、海洋、空氣塵埃等環境中都有發現。鏈霉菌產生的次生代謝產物更是藥物研發的重要來源,在藥物研發中發揮著重要的作用[15-16]。統計數據表明,以鏈霉菌為代表的放線菌貢獻了一半以上的臨床抗生素藥物及先導物[17-19],鏈霉菌屬微生物占據了微生物代謝產物的39%。目前發現的1 000多種微生物生物活性物質中,由鏈霉菌屬微生物所產生的次生代謝產物中得到的抑菌活性物質約占2/3[20]。據報道,鏈霉菌對魚類病原菌有抑制效果,其中李夢茜等[21]分離的鏈霉菌LFJK-11發酵液對引起羅非魚敗血癥的嗜水氣單胞菌有較強的抑制作用。這與本文中研究的鏈霉菌I6發酵液對引起魚類癤瘡病的病原菌殺鮭氣單胞菌具有很強的抑制活性相類似。
殺鮭氣單胞菌廣泛分布于自然界中,是氣單胞菌中最早被鑒定的致病菌,能引起癤瘡病,對魚類的存活有著顯著的影響。目前常用藥物治療的方法防治殺鮭氣單胞菌。但是隨著藥物的使用,殺鮭氣單胞菌的耐藥性越來越強,甚至出現了可移動的耐藥性基因[22]。也有研究發現,殺鮭氣單胞菌還可以和常見的嗜水氣單胞菌等細菌引發魚類的混合感染。因此,尋求水產防治殺鮭氣單胞菌的益生菌迫在眉睫。本研究中篩到的鏈霉菌I6對殺鮭氣單胞菌有很強的抑制作用,更慶幸的是,鏈霉菌I6發酵液對殺鮭氣單胞菌的MIC分別相當于慶大霉素7.67 μg/mL、諾氟沙星665 μg/mL的效價,且對魚類細胞無毒性,是一種益生菌。這與陸婷巍等[23]的殺鮭氣單胞菌拮抗菌X8安全性測試結果相同。因此鏈霉菌I6的活性產物有望開發成新的微生物制劑應用于水產疾病的防控中,具有潛在的經濟效益和社會效益。但目前研究抗魚類病原菌活性化合物生物合成途徑的報道寥寥無幾。因此,我們將繼續對鏈霉菌I6發酵液中抗殺鮭氣單胞菌活性化合物進行分離鑒定,并根據其分子結構、結合基因編輯和生物信息學技術研究其生物合成途徑。
