垂釣強度對鯽幼魚易釣性和生長的影響
2021-09-08 07:10:50劉小玲曾令清
水生生物學(xué)報 2021年5期
關(guān)鍵詞:實驗
陳 歡 何 衍 劉小玲 曾令清
(重慶師范大學(xué)生命科學(xué)學(xué)院, 進化生理與行為學(xué)實驗室, 動物生物學(xué)重慶市重點實驗室, 重慶 401331)
全世界淡水、海洋和陸地生態(tài)系統(tǒng)中都有人類活動引起的動物表型反應(yīng)[1]。動物種群內(nèi)個體之間行為和生理上的差異影響個體的生理狀態(tài)和生態(tài)表現(xiàn)[2]。然而, 在特定環(huán)境中行為或生理的最適表型可能會因人類活動而發(fā)生改變, 從而導(dǎo)致受影響動物表型的進化反應(yīng)。例如, 在漁業(yè)(包括休閑漁業(yè))生產(chǎn)中, 對具有特定表型個體的選擇性捕獲可以驅(qū)動魚類表型的進化方向, 即漁業(yè)導(dǎo)致進化(Fisheries induced evolution)[3—5]。這些研究大多集中于生活史特征的改變[6], 如在商業(yè)捕撈下海洋魚類種群的生長速度下降、總生殖產(chǎn)量降低和性成熟年齡降低[4,7]。類似的過程也可能發(fā)生在以休閑垂釣為目標的淡水漁業(yè)中[8,9]。易釣性(Vulnerability to angling)是指魚類種內(nèi)個體間被釣概率在不同時空條件下的穩(wěn)定差異, 即某些個體一直容易被釣, 而某些個體一直難于被釣, 是一個復(fù)雜的遺傳性狀[10]。垂釣(如競技垂釣和休閑垂釣)可能改變野生魚類種群的基因庫, 間接影響種群的生存、生長、繁殖等生活史特征, 并影響垂釣成功率[10]。
當(dāng)今休閑漁業(yè)在世界范圍內(nèi)非常流行[11,12], 還是魚類資源調(diào)查的重要手段[13—15]。全球范圍的休閑漁民數(shù)量相當(dāng)可觀, 而且還在不斷增加[16]。垂釣已成為重要的休閑漁業(yè)活動, 據(jù)統(tǒng)計, 具有被釣經(jīng)歷的魚類數(shù)量約占全球每年總漁獲量的12%[17]。多數(shù)垂釣愛好者每年垂釣次數(shù)超過10次, 多者達到30次以上, 在春季垂釣的人數(shù)較多[18], 這一現(xiàn)狀可能對繁殖季節(jié)魚類的繁殖活動造成負面影響。社會學(xué)習(xí)發(fā)生在包括魚類在內(nèi)的許多動物類群中[19],魚類可能對垂釣過程中人工誘餌及其引發(fā)的脅迫進行社會學(xué)習(xí)。例如, 日本比目魚(Paralichthys olivaceus)通過觀察其他同類個體被捕食經(jīng)歷而成功有效避開捕食者[20]; 先捕后放的社會經(jīng)歷誘導(dǎo)鯉(Cyprinus carpio)產(chǎn)生魚鉤回避行為以降低被捕風(fēng)險, 可能影響魚類的易釣性[21]。雖然魚類易釣性與個體的勇敢性[22,23], 營養(yǎng)狀況和饑餓水平[24], 捕食風(fēng)險[25]和溫度[26]等因素相關(guān), 但在休閑垂釣活動中不同強度的垂釣活動對魚類易釣性影響的相關(guān)研究仍顯匱乏。
鯉科(Cyprinidae)是我國淡水魚類中種數(shù)最多的一個科, 在淡水養(yǎng)殖業(yè)和捕撈業(yè)及水產(chǎn)品動物蛋白供應(yīng)中具有不可取代的作用與地位, 也是內(nèi)陸水域(如長江、黃河)或人工池塘的常見垂釣種類。鯽(Carassius auratus)是一種我國常見的鯉科魚類和經(jīng)濟養(yǎng)殖魚類, 棲息在青藏高原地域以外的各大水系, 具有食性廣、適應(yīng)性強、繁殖力強和抗病力強等特點。研究發(fā)現(xiàn), 鯽幼魚的易釣性存在明顯的個體穩(wěn)定差異[27]; 異育銀鯽(Carassius auratus gibelio)的易釣性明顯強于鯉[28]; 短期饑餓增強草魚(Ctenopharynodon idellus)的易釣性[29], 但降低了異育銀鯽的易釣性[30]。為考察不同垂釣強度對鯉科魚類易釣性和生長的影響, 本研究以鯽幼魚為實驗對象,對3個不同垂釣強度組[高強度組(1d垂釣1次)、中強度組(2d垂釣1次)、低強度組(4d垂釣1次)]進行垂釣活動, 每個垂釣處理組均設(shè)3個平行組, 每個平行組包括40尾鯽幼魚, 每個重復(fù)在垂釣10h后則停止該組的垂釣活動, 記錄成功垂釣每尾魚的時間、序號和電子信息, 計算垂釣比例、單尾被釣時間、平均垂釣序號及其變異系數(shù)(Coefficient of variation,CV)和實驗期間的生長率。本研究結(jié)果不僅可以為魚類易釣性、增養(yǎng)殖和漁業(yè)管理等提供基礎(chǔ)資料,還可以為政府漁業(yè)管理部門的垂釣指南制定提供重要參考依據(jù)。
1 材料與方法
1.1 實驗魚來源及馴化
本研究采用的鯽幼魚來自重慶市北碚區(qū)當(dāng)?shù)氐娜斯し敝臭~場, 購回后將魚放入重慶師范大學(xué)進化生理與行為學(xué)實驗室的3個室內(nèi)循環(huán)控溫水養(yǎng)殖水槽中進行為期4周的室內(nèi)馴化。馴化期間, 前期每日09: 00用商業(yè)鯉科魚類通用飼料(中國通威公司)飽足投喂, 后期每日09:00用活體蚯蚓按1%魚群總體重的量進行投喂, 使魚提前適應(yīng)蚯蚓餌料。這種餌料的馴化原因是基于預(yù)備實驗發(fā)現(xiàn), 即使在商業(yè)飼料中加入黏合劑重新制成人工顆料誘餌, 但在水中仍會出現(xiàn)一定程度的溶解, 導(dǎo)致部分個體攝入溶解了的食物殘渣, 影響魚類的食欲及易釣性, 導(dǎo)致垂釣時間過長, 使得魚群中所有個體的營養(yǎng)狀態(tài)無法達到較好的局部控制。每次投喂5min前, 將養(yǎng)殖水槽的充氧泵和循環(huán)系統(tǒng)關(guān)閉, 以減少水體曝氣和循環(huán)對魚攝食的干擾; 投喂30min后, 用虹吸管將殘餌、殘渣及糞便全部清除, 以維持養(yǎng)殖水體的質(zhì)量, 但每天的維護工作導(dǎo)致養(yǎng)殖水體的部分減少(約馴化水體的10%)。因此, 每隔3天對養(yǎng)殖水槽補充曝氣3d的水體至正常水量。循環(huán)水槽的水溫控制在(26.9±0.1)℃, 溶氧水平控制在7.0 mg/L以上,光周期設(shè)定為14 L﹕10D。為便于區(qū)分不同的鯽幼魚個體, 實驗開始前2周, 對經(jīng)MS-222(80 mg/L)麻醉后的實驗魚進行PIT(Passive integrated transponder)數(shù)字標簽(直徑2 mm×長8 mm, 0.03 g/個)體內(nèi)植入, 個體的電子標簽信息可通過掃描儀檢測獲得。在標簽植入后的2周內(nèi), 未發(fā)現(xiàn)鯽幼魚個體死亡情況。
1.2 實驗設(shè)計
本研究共設(shè)置3種不同的垂釣強度, 即高垂釣強度組(1d垂釣1次)、中垂釣強度組(2d垂釣1次)、低垂釣強度組(4d垂釣1次)。基于預(yù)備實驗, 本研究共進行3個9d的實驗周期。在每個實驗周期, 高強度組在1—9d中每天垂釣1次, 中強度組在第1、第3、第5、第7、第9天中每天垂釣1次, 低強度組在第1、第5和第9天中每天垂釣1次。每個處理組的垂釣活動時間為當(dāng)天08:30—18:30(10h), 在到達規(guī)定時間后, 不管桶中的魚是否全部被成功垂釣, 立即停止該組的垂釣活動。這種方式的設(shè)定目的是通過垂釣在單位時間內(nèi)對魚群施加垂釣脅迫, 并非獲得百分之百的垂釣比例。在馴化結(jié)束后, 篩選出大小相近、體格健康的鯽幼魚360尾作為實驗對象(表 1)。在每次垂釣之前, 將3個處理組的魚群轉(zhuǎn)移至相應(yīng)的垂釣桶中適應(yīng)并禁食48h, 以排空消化道內(nèi)容物并維持實驗魚的一定食欲。在實驗期間, 實驗室維持安靜狀態(tài)以減少聲音對魚的影響, 環(huán)境條件(如水溫、溶氧水平和光周期等)與馴化期間保持一致。

表1 鯽幼魚的形態(tài)學(xué)參數(shù)Tab. 1 The morphology of juvenile crucian carp
1.3 實驗裝置及流程
實驗裝置 本研究采用3個完全相同的藍色垂釣桶(高94 cm×直徑68 cm, 水深80 cm)進行實驗,將3個垂釣桶放置在同一水平線上的位置, 保持相等間距。在桶中加入充分曝氣3d后的自來水, 在每個垂釣桶的底部均勻放置一定數(shù)量的綠色塑料水草及鵝卵石等, 以模擬魚類野外棲息環(huán)境并提高垂釣環(huán)境的富集度(圖 1)。每個垂釣桶均配置一個微循環(huán)系統(tǒng)和一個充氧砂頭。在每個垂釣桶的頂部蓋有不透光和顏色均勻的黃色硬紙板, 在其中間具有一個25 cm×25 cm的正方形垂釣孔。垂釣人員與垂釣桶保持1.2 m的距離, 有助于減少垂釣過程中垂釣人員的操作對實驗魚的潛在影響; 在垂釣桶周圍鋪置一層橡膠發(fā)泡板, 以減小取魚鉤時被釣個體出現(xiàn)滑落而造成的損傷。
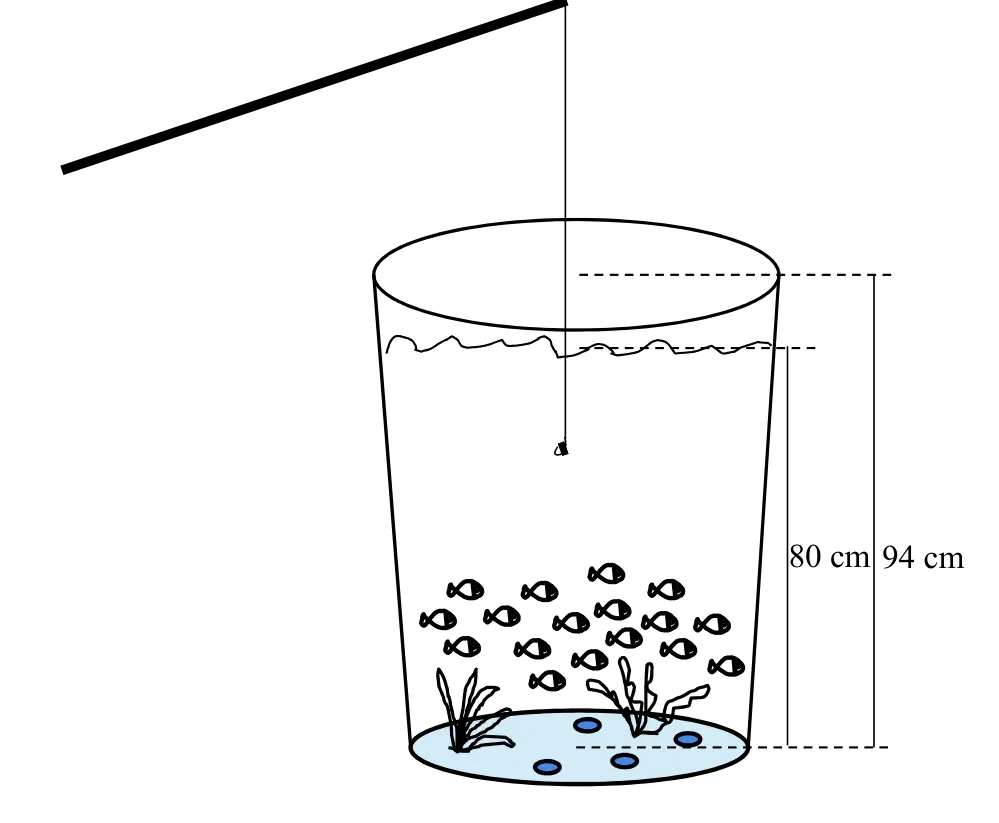
圖1 垂釣裝置的示意圖Fig. 1 Schematic of the angling device
實驗流程 在垂釣前, 檢查每組魚的活躍情況, 測定垂釣桶中水體的溶氧水平和水溫并做好記錄, 準備好完全一致的垂釣漁具和垂釣餌料(長度5 mm的新鮮蚯蚓段塊)及用于暫養(yǎng)被釣個體的塑料水桶(30 L)。在垂釣期間, 關(guān)閉充氣泵和微循環(huán)系統(tǒng), 將裝有魚鉤的誘餌輕輕沉入垂釣桶中部的空間位置(距離水面40 cm), 通過觀察浮標來判斷魚的吃餌情況, 當(dāng)魚因被釣而劇烈逃逸拉動浮漂時, 則迅速將魚釣出并快速小心取出魚鉤, 用掃描儀獲取被釣個體的電子標簽信息, 隨后將魚置于含有阿莫西林水體的暫養(yǎng)水桶中恢復(fù)2h, 以減少損傷部位的感染機率, 最后記錄該尾魚的被垂釣時間和被釣序號。在垂釣后, 將成功垂釣起來的所有個體全部放回垂釣桶, 每日18:40對每個強度組投喂0.5%魚群總體重的新鮮蚯蚓, 隨后用虹吸管清除殘渣和糞便以維持垂釣桶的水質(zhì), 并補充適當(dāng)曝氣后的自來水以維持正常水位。
1.4 實驗參數(shù)
易釣性 本研究采用垂釣比例、單尾被釣時間、平均垂釣序號及其變異系數(shù)來評價魚類易釣性[27—30]。垂釣于當(dāng)天8:30開始, 從魚餌放入垂釣桶中開始至第1尾魚釣出水面的時間為垂釣第1尾魚的所需時間, 其余釣出的每尾魚計時方法同上,垂釣活動直至18:30。魚類易釣性的實驗參數(shù)如下所示:
(1)垂釣比例(%)是指在10h垂釣時間內(nèi)成功垂釣起來的鯽幼魚的個體數(shù)量占單個魚群數(shù)量(n=40)的比例。若該比例越大, 則這個魚群的易釣性就越強。
(2)單尾被釣時間(min)是從指上一尾魚的被釣時間到下一尾魚被釣時間的時間間隔[28—30]。若1個魚群的單尾被釣時間的平均值越小, 則這個魚群的易釣性就越強。
(3)平均垂釣序號是指單尾魚在1個實驗周期中所有被釣序號的平均值[28—30]。每次垂釣活動中對被釣個體的先后順序依次從1至最后一尾編號(如40), 因此在每次垂釣中釣出的每尾魚具有唯一的序號。若單尾魚的平均垂釣序號越小, 則該尾魚的易釣性就越強。
(4)平均垂釣序號CV是指單尾魚在1個實驗周期中所有被釣序號的變異系數(shù), 評價被釣個體的垂釣序號的變異程度[28—30]。若單尾魚的平均垂釣序號CV的數(shù)值越小, 則該尾魚在此實驗周期中的易釣性越穩(wěn)定, 即具有較強的穩(wěn)定性, 反之成立。
生長率 由于高強度垂釣實驗可能對實驗魚產(chǎn)生更強的脅迫作用從而對其生長率產(chǎn)生更為顯著的影響, 故本研究采用實驗魚體重的特定生長率(Specific growth rate,SGR)評價不同垂釣強度組在每個實驗周期的體重變化情況, 即單位時間內(nèi)魚體重自然對數(shù)的增長值, 其計算公式如下所示:

式中,SGR是實驗魚體重的特定增長率(%/d),W0為實驗魚在0時的初始體重(g),W9為實驗魚在第9天時的最終體重(g), 9為實驗時間(d)。
1.5 數(shù)據(jù)分析
所有數(shù)據(jù)先用Excel2010進行初步整理, 然后再用軟件SPSS 19.0進行統(tǒng)計分析。(1)用雙因素方差分析(Two-way ANOVA)考察垂釣強度和時間對鯽幼魚垂釣比例和單尾被釣時間的影響, 再用單因素方差分析進行多重比較。(2)用單因素方差分析考察垂釣強度對特定生長率、平均垂釣序號及其C V的影響, 若組間存在顯著性影響, 再采用Duncan進行多重比較。(3)用Pearson相關(guān)性分析考察垂釣比例與單尾被釣時間、平均垂釣序號與其CV之間的相關(guān)性。所有數(shù)據(jù)均以平均值±標準誤(Mean±SE)表示, 顯著性水平定在P<0.05。
2 結(jié)果
2.1 垂釣強度對垂釣比例和單尾被釣時間的影響
本研究發(fā)現(xiàn), 垂釣強度和垂釣時間對鯽幼魚垂釣比例和單尾被釣時間均有影響, 即3個垂釣強度組的垂釣比例均呈現(xiàn)下降的變化趨勢, 單尾被釣時間呈現(xiàn)增加的變化趨勢(圖 2和圖 3)。第1天, 高強度組、中強度組和低強度組之間的垂釣比例均超過了95%, 并且無顯著性差異(圖 2;P=0.667), 說明實驗開始時3個垂釣強度組的易釣性維持相當(dāng)。第5天, 中強度組與低強度組間的垂釣比例無顯著性差異, 但均明顯大于高強度組的垂釣比例(圖 2;P<0.05)。第9天, 中強度組和低強度組間的垂釣比例仍無差異, 但明顯高于高強度組; 相比較于第5天, 中強度組和低強度組的垂釣比例與高強度組之間的差異存在明顯擴大的變化趨勢(圖 2;P<0.05), 表明在第9天中, 中強度組和低強度組的易釣性比高強度組更強。
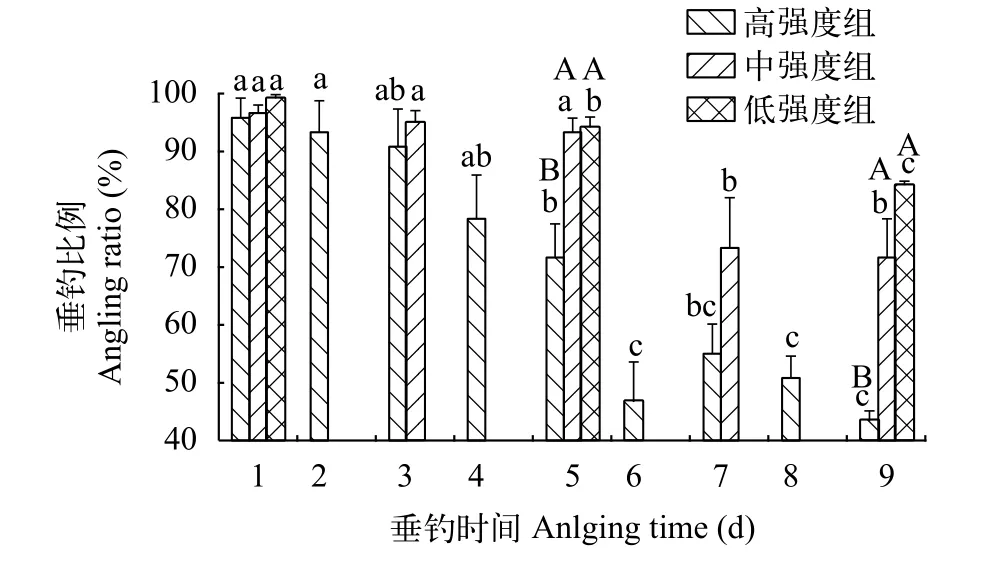
圖2 不同垂釣強度和不同垂釣時間下鯽幼魚的垂釣比例Fig. 2 Angling ratio of the juvenile crucian carp under the conditions of different angling intensities and different angling times
本研究發(fā)現(xiàn), 高強度組和中強度組的單尾被釣時間隨著垂釣時間的延長而明顯增加, 但低強度的單尾被釣時間無顯著變化(圖 3)。第1天, 高強度組、中強度組和低強度組的單尾被釣時間均為5min左右, 并且無顯著性差異(圖 3;P=0.590), 說明實驗開始時3個垂釣強度組的易釣性無差異。第5天, 3個強度組的單尾被釣時間仍無顯著性差異,但相比較于第1天, 僅高強度組的單尾被釣時間顯著增加到3倍以上(圖 3;P<0.05)。第9天, 3個強度組的單尾被釣時間之間均存在顯著性差異, 高強度組最大, 中強度組次之, 低強度組最小(圖 3;P<0.05)。
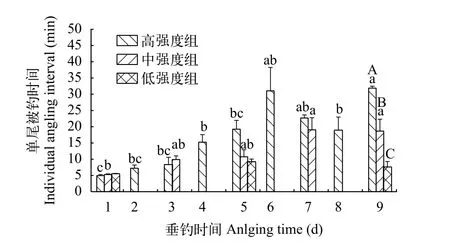
圖3 不同垂釣強度和不同垂釣時間下鯽幼魚單的尾被釣時間Fig. 3 Individual angling interval of the juvenile crucian carp under the conditions of different angling intensities and different angling times
本研究發(fā)現(xiàn), 高強度組和中強度組的垂釣比例與單尾被釣時間存在負相關(guān), 即在中高垂釣強度條件下, 實驗魚的易釣性不僅明顯降低, 而且其每尾魚被釣的時間間隔也逐漸增加(圖 4;P<0.05)。然而, 低強度組的垂釣比例與單尾被釣時間無顯著相關(guān)性(圖 4;P=0.402)。

圖4 不同垂釣強度組鯽幼魚垂釣比例與單尾被釣時間的相關(guān)性Fig. 4 Relationships between angling ratio and individual angling interval of the three angling intensity treatments in juvenile crucian carp
2.2 垂釣強度對平均垂釣序號及其CV的影響
本研究發(fā)現(xiàn), 除低強度組外, 垂釣強度對高、中強度組的平均垂釣序號及其CV均有影響(圖 5),即2個垂釣強度組的平均垂釣序號隨著被釣次數(shù)的增加均呈現(xiàn)減小的變化趨勢, 暗示在連續(xù)垂釣過程中經(jīng)常被釣個體的易釣性較強并且維持較為穩(wěn)定。高強度組和中強度組平均垂釣序號的CV隨著被釣次數(shù)的增多均呈增大的變化趨勢(圖 6), 說明個體的被釣次數(shù)越多, 其平均垂釣序號越不穩(wěn)定。此外, 本研究還發(fā)現(xiàn), 3個強度組的平均垂釣序號與其CV均呈負相關(guān), 即平均垂釣序號越大, 其變異系數(shù)越小(圖 7;P<0.05)。
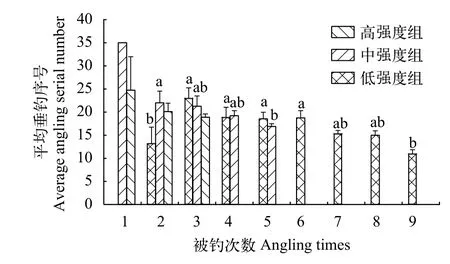
圖5 不同垂釣強度和不同垂釣時間下鯽幼魚的平均垂釣序號Fig. 5 Average angling serial number of the juvenile crucian carp under the conditions of different angling intensities and different angling times
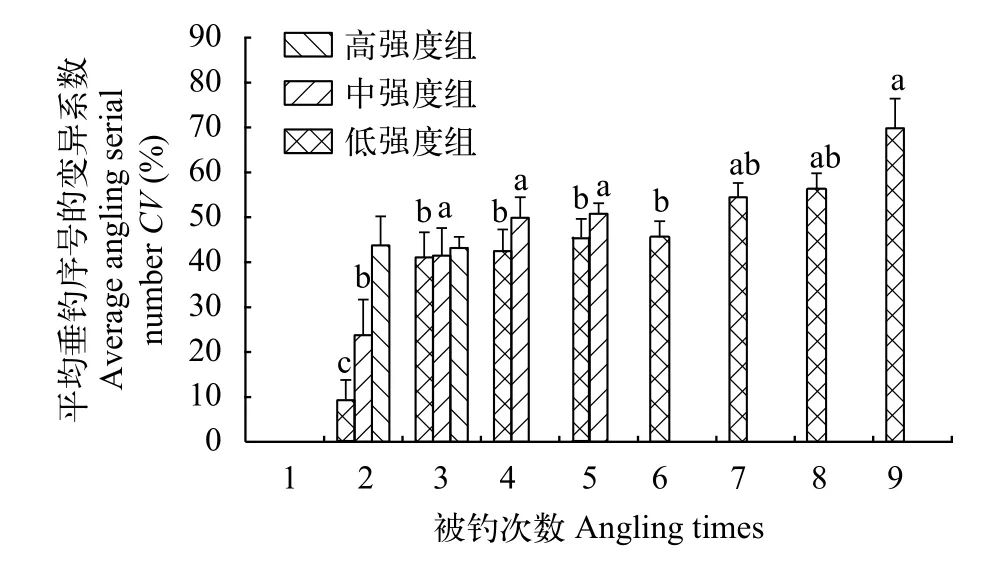
圖6 不同垂釣強度和不同垂釣時間下鯽幼魚平均垂釣序號的變異系數(shù)Fig. 6 Coefficient variation of average angling serial number of the juvenile crucian carp under the conditions of different angling intensities and different angling times
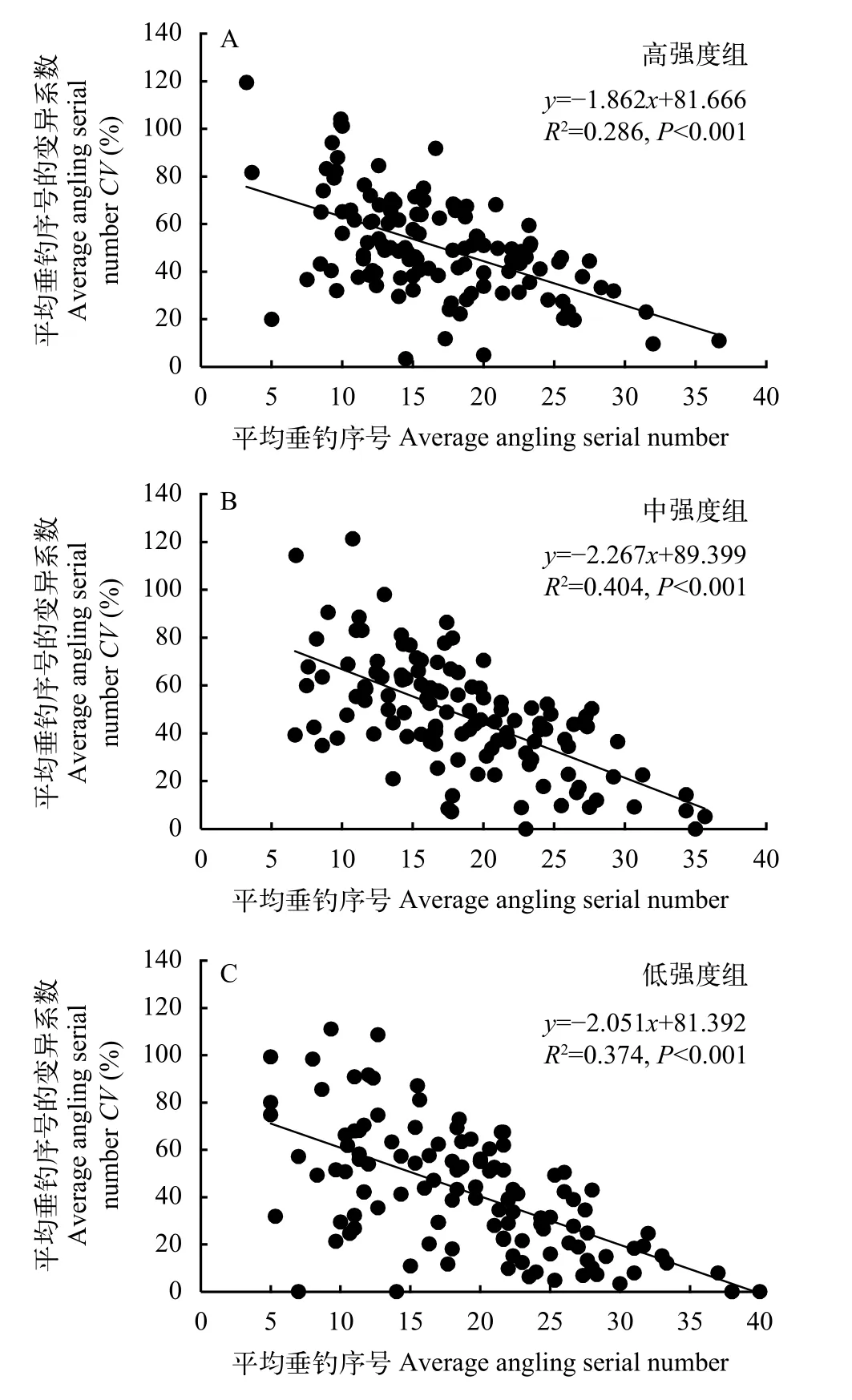
圖7 鯽幼魚平均垂釣序號與其變異系數(shù)的相關(guān)性Fig. 7 Correlations between average angling serial number and its coefficient variation in juvenile crucian carp
2.3 垂釣強度對特定生長率的影響
本研究發(fā)現(xiàn), 3個垂釣強度的平均特定生長率均出現(xiàn)負值情況, 但3個組之間無顯著差異(圖 8)。進一步分析, 中強度組和低強度組在不同被釣次數(shù)的特定生長率無顯著差異, 但高強度組在不同被釣次數(shù)的體重特定生長率具有差異, 即隨著被釣次數(shù)的增多特定生長率呈先上升后下降的變化規(guī)律(圖 9)。
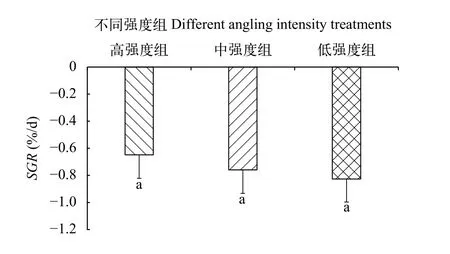
圖8 不同垂釣強度下鯽幼魚的特定體重生長率Fig. 8 Specific growth rate of the juvenile crucian carp under different angling intensities
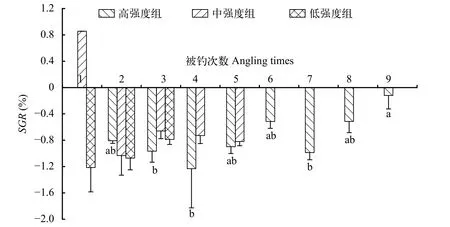
圖9 不同垂釣頻次下鯽幼魚的特定體重生長率Fig. 9 Specific growth rate of the juvenile crucian carp under different angling frequencies
3 討論
3.1 垂釣強度對鯽幼魚易釣性及其穩(wěn)定性的影響
本研究發(fā)現(xiàn), 隨著垂釣次數(shù)的增加, 3個垂釣強度組鯽幼魚的易釣性均出現(xiàn)降低的現(xiàn)象, 即垂釣比例下降、單尾被釣時間增加, 導(dǎo)致垂釣比例與單尾被釣時間呈負相關(guān), 表明無論何種垂釣強度, 重復(fù)性的垂釣均會降低鯽幼魚的易釣性。出現(xiàn)該現(xiàn)象的可能原因是在反復(fù)的垂釣活動中鯽對人工誘餌的學(xué)習(xí)能力使其產(chǎn)生了垂釣記憶, 個體再次面對暗藏魚鉤的人工誘餌時可能表現(xiàn)更加謹慎的鉤回避行為。在垂釣過程中, 被成功垂釣的鯽個體遭受被魚鉤扎住及離開水體暴露于空氣等嚴重環(huán)境脅迫,而未被成功垂釣的鯽個體也獲取有關(guān)垂釣脅迫中的社會信息(如同類個體被釣和人工誘料的突然移動造成的水體動蕩等), 當(dāng)這些魚類再次面臨相似的垂釣情境時可能會表現(xiàn)得較為謹慎而降低易釣性。研究發(fā)現(xiàn), 魚類個體在其經(jīng)歷被捕獲和釋放的脅迫經(jīng)歷中獲得有效的經(jīng)歷學(xué)習(xí)[31—35], 如鯉能夠通過該經(jīng)歷學(xué)習(xí)能力而產(chǎn)生明顯的鉤回避行為, 降低魚類的易釣性[21,36—38]。此外, 魚類還會觀察其他同種個體的被捕獲過程而進行社會學(xué)習(xí)[19], 如日本比目魚(Paralichthys olivaceus)成功地學(xué)會了通過觀察其他同種個體被捕食過程而有效地回避捕食者,有助于提高學(xué)習(xí)者的生存適合度[20]。因此, 在垂釣過程中那些未被釣個體也可能通過社會學(xué)習(xí)而產(chǎn)生鉤回避行為, 進而降低了這些未被釣個體的易釣性。
本研究發(fā)現(xiàn), 相比較于中強度組和低強度組,高垂釣強度導(dǎo)致鯽幼魚易釣性的下降程度更大。出現(xiàn)這種現(xiàn)象的可能原因是當(dāng)對鯽進行高強度垂釣時, 鯽幼魚經(jīng)歷了更多次的先被釣后釋放過程,這種更為豐富的個體脅迫經(jīng)驗明顯增強了高強度組個體的經(jīng)驗學(xué)習(xí)效果。此外, 在高強度垂釣過程中鯽幼魚觀察同種其他個體的被垂釣整個過程, 使觀察個體產(chǎn)生了更有成效的鉤回避行為。相比較于高強度組, 雖然中低垂釣強度也導(dǎo)致鯽產(chǎn)生一定的對垂釣活動的社會學(xué)習(xí)和經(jīng)驗學(xué)習(xí), 但由此產(chǎn)生的鉤回避行為次數(shù)可能相對較少。因此, 中低垂釣強度組可能仍然表現(xiàn)相對積極的攝食行為, 導(dǎo)致中低強度組鯽個體仍維持較高的易釣性, 而高強度組鯽幼魚的易釣性明顯下降。
魚類的易釣性作為一種復(fù)雜的遺傳性狀, 在種間和種內(nèi)都存在個體差異[10], 如在虹鱒(Oncorhynchus mykiss)中的這種差異表現(xiàn)為有些個體可以多次被捕獲, 呈現(xiàn)出較高的易釣性, 而有些個體盡管經(jīng)過反復(fù)的垂釣努力, 仍然未能被捕獲, 表現(xiàn)為較低的易釣性[39]。有關(guān)鯽幼魚重復(fù)垂釣的研究發(fā)現(xiàn), 每天都被釣的鯽幼魚個體的比例占7.5%, 表明該種魚的前期被釣經(jīng)歷并不影響這些個體后期再次被釣, 而17.5%鯽幼魚個體在實驗過程中從未被釣[27], 上述結(jié)果說明不同易釣性的鯽幼魚個體在整體實驗過程中保持較高的不易釣性。本研究也發(fā)現(xiàn), 在高中低垂釣強度組中每天都被釣的鯽幼魚個體的比例分別為13.3%、51.6%和79.2%, 表明在垂釣強度越低的環(huán)境條件下實驗魚全部被釣的比例反而越高。另外, 本研究還發(fā)現(xiàn), 在所有垂釣強度條件下, 被釣次數(shù)越多的個體的平均垂釣序號越小,但其CV卻越大, 表明易釣性越高的個體, 其穩(wěn)定性相對較差。出現(xiàn)這種現(xiàn)象的原因可能是相較于中低易釣性個體, 高易釣性的鯽幼魚個體更為活躍和勇敢, 可能增加魚類個體與漁具的接觸概率[40], 從而使其在垂釣實驗前期表現(xiàn)為更加積極頻繁地攝取人工誘餌, 同時提高了其被捕獲的概率, 導(dǎo)致在垂釣過程中高易釣性的鯽個體經(jīng)歷了更多次被捕獲和釋放的過程, 并由此累積了更豐富的人工誘餌的識別經(jīng)驗而表現(xiàn)出更為謹慎的攝餌行為, 在垂釣實驗后期中這些表型個體易釣性發(fā)生波動的可能性更高, 而發(fā)生易釣性更不穩(wěn)定的現(xiàn)象。
3.2 垂釣強度對鯽幼魚生長率的影響
研究發(fā)現(xiàn), 具有較高能量代謝率和體型較大(如體重和體長)的魚類個體更易被釣[27,28,41,42], 并且這些高易釣性的魚類個體卻具有較低的生長率[43],這種現(xiàn)象可能是因為易釣性高的個體具有較高的能量代謝強度, 在行為上表現(xiàn)為更為勇敢、敢于探索等, 在垂釣過程中頻繁靠近餌料而消耗的能量也相對較多, 降低了生長率。然而, 本研究發(fā)現(xiàn)3個垂釣強度組中不同易釣性個體的平均生長率均出現(xiàn)負生長, 表明原先設(shè)定的每天餌料投放比例低于理論估計值。此外, 3個垂釣強度組之間的生長率無明顯差異, 說明不同垂釣強度對鯽幼魚生長造成的負面影響并無差異; 相比較于中低強度組, 只有高強度組個體的生長率隨著被釣次數(shù)的增加呈現(xiàn)下降的變化趨勢, 表明易釣性越高的魚類個體, 其負生長率反而較低。出現(xiàn)這種現(xiàn)象的原因可能是在該組的垂釣過程中, 高易釣性個體積極覓食, 雖然會消耗更多能量但是也可在此過程中有更多的機會成功獲取餌料, 或在每天晚上餌料投喂過程中因競爭能力強而獲取更多食物, 使得其體重損失減少,導(dǎo)致其生長率的絕對值小于中低易釣性個體。
綜上所述, 隨著垂釣強度的增加, 鯽幼魚的易釣性將降低, 即高強度的垂釣會降低鯽幼魚的易釣性。然而, 易釣性越高的個體卻具有較低的易釣穩(wěn)定性, 說明高強度的垂釣還會導(dǎo)致鯽幼魚的易釣性表現(xiàn)出較低的穩(wěn)定性。因此, 對于不論休閑漁業(yè)(如野外或競技垂釣)或經(jīng)濟漁業(yè)而言, 對目標魚類的垂釣強度不宜設(shè)定過強, 否則會導(dǎo)致這些魚類的易釣性出現(xiàn)較大程度下降, 不僅影響垂釣效率和垂釣者的滿意度, 也可能影響種群表型的可塑性。此外, 雖然不同垂釣強度組之間鯽幼魚的生長率并無差異, 但均呈現(xiàn)負生長現(xiàn)象, 表明被垂釣魚塘中魚類個體的日常能量消耗明顯大于非垂釣魚塘, 建議適當(dāng)增加投餌量以保障被垂釣魚塘中魚類個體較高的營養(yǎng)需求。
猜你喜歡
作文·小學(xué)低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學(xué)生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學(xué)低年級(2024年2期)2024-04-29 00:00:00
作文·小學(xué)低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學(xué)生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
