基于線粒體Cyt b基因的長江刀鱭群體遺傳結構分析
2021-09-08 01:19:14楊彥平許萌原馬鳳嬌郭文君王銀平
江西農業學報 2021年8期
楊彥平,許萌原,馬鳳嬌,郭文君,3,代 培,姜 敏,王銀平,劉 凱**
(1.農業農村部 長江下游漁業資源環境科學觀測實驗站/中國水產科學研究院 淡水漁業研究中心, 江蘇 無錫 214081;2.南京農業大學 無錫漁業學院,江蘇 無錫 214081;3.上海海洋大學 水產科學國家級實驗教學示范中心,上海 201306)
刀鱭(Coilianasus)是一種小型洄游性魚類,隸屬于鯡形目(Clupeiformes)、鳀科(Engraulidae)、鱭屬(Coilia),主要分布于中國沿海及長江中下游地區[1]。刀鱭為典型的江海洄游性魚類,但其有多個生態型,除洄游型群體外,還有陸封型群體湖鱭(C.nasustaihuensis)和定居性群體短頜鱭(C.brachygnathus)[2];洄游型刀鱭每年2月開始在河口區陸續集群,然后溯河而上洄游;定居型刀鱭一般分布于長江下游及附屬湖泊[3]。刀鱭肉質鮮美,被列為“長江三鮮”之首,深受廣大群眾喜愛,具有重要的經濟價值,是我國長江下游及河口區主要經濟魚類之一[4-5]。自20世紀70年代以來,水環境污染加劇,江湖聯系阻斷等導致刀鱭生存環境惡化,適宜繁殖場喪失,加之持續高強度捕撈導致其資源急劇衰退,種群結構逐漸趨于低齡化和小型化[1,6-7]。鄭飛等[8]通過對1973~2009年不同時期的長江刀鱭洄游群體的體長、體重、年齡結構進行綜合比較,得出結論:長江洄游型刀鱭群體無論是體長、體重,還是年齡結構、優勢年齡組,在總體上已呈現出逐漸變小的趨勢。張敏瑩等[9]統計的長江下游刀鱭汛期捕撈量由1995年的1425.00 t銳減至2002年的285.52 t。各方面數據均顯示,長江刀鱭資源被過度利用,種質資源保護和種群恢復刻不容緩。種群遺傳多樣性不僅是形成生物多樣性的基礎,也是物種進化潛能的保證。針對重要物種開展種群遺傳結構研究、了解物種恢復潛力,有助于制定針對性的保護措施[5,10]。
目前,馬春艷等[4,11-13]分別用RAPD、AFLP、線粒體DNA D-loop全序列分子標記和微衛星標記等技術對長江刀鱭遺傳多樣性水平進行了研究,發現長江刀鱭遺傳多樣性水平較為豐富。張媛等[14]利用同工酶、RAPD-PCR和ISSR-PCR等標記技術對長江口刀鱭種群遺傳結構進行了研究,發現長江口刀鱭個體之間遺傳變異水平較高,沒有出現明顯的群體分化。
細胞色素b (Cytochrome b,Cytb)基因是一種線粒體蛋白質編碼基因,約由1140個堿基組成,目前是mtDNA中唯一一個結構和功能被了解的較為清楚的蛋白編碼基因,該基因進化速度適中而且容易使用通用引物擴增和測序,其變異程度足以闡明種間的系統發育關系,又有一定保守性,可進行種上階元水平的研究,適用于一些親緣關系較近群體的遺傳結構研究[15],被認為是了解種質資源狀況和群體遺傳學的理想工具,近年來已被廣泛應用于魚類分子系統學和遺傳多樣性、遺傳結構、物種品系鑒定等方面的研究,尤其在近緣物種間的親緣關系研究中更為有效,也是魚類遺傳學研究中最為常用的目標序列[15-18]。但基于Cytb基因序列研究長江刀鱭種群遺傳結構的報道并不多見[19]。本研究在長江禁漁退捕的背景下,基于Cytb基因序列對長江4個區段刀鱭群體進行種群遺傳結構分析,初步探索了禁漁對長江刀鱭群體遺傳結構的影響,以期為長江刀鱭種質資源養護及后續管理提供科學支撐。
1 材料與方法
1.1 實驗材料
2019年4~6月在長江下游安徽段安慶 (AQ,30°28′30″~30°29′23″ N,116°58′50″~116°59′56″ E)、銅陵-當涂 (TD,31°0′5″~31°32′54″ N、117°46′23″~118°22′14″ E)、江蘇段南京-鎮江 (NZ,32°11′22″~32°14′39″ N,118°57′26″~119°40′42″ E)、長江口崇明 (CM,31°31′08″~31°30′16″ N,121°29′50″~121°40′32″ E)4個調查區段(圖1),使用流刺網和拋定刺網采集刀鱭樣本,每個區段均為雌雄各20尾。取樣本背鰭或胸鰭,固定于無水乙醇中,常溫保存備用。部分樣本以血細胞(-20 ℃保存)為實驗材料。
1.2 基因組DNA的提取
DNA提取所用試劑盒:TaKaRa MiniBEST Universal Genomic DNA Extraction Kit Ver 5.0。提取過程主要包括組織裂解、分離蛋白、洗脫和DNA收集,具體操作步驟參照試劑盒說明書。
1.3 DNA檢測
提取的DNA通過1%瓊脂糖凝膠電泳進行質量檢測。
1.4 PCR擴增與測序
PCR反應體系:引物各2 μL、2×Taq Master Mix(內含Taq、buffer、dNTPs等) 25 μL,以及模板DNA 2 μL,加滅菌雙蒸水至50 μL。擴增mtDNA控制區序列的引物序列為L14724 (5′-GACTTGAAAAACCACCGTTG-3′ )和H15915 (5′-CTCCGATCTCCGGATTACAAGAC-3′ )[20]。
PCR反應條件為:94 ℃預變性5 min;然后進行94 ℃變性30 s,55 ℃退火30 s和72 ℃延伸30 s,共進行33個循環;最后72 ℃延伸7 min。
進行PCR擴增后,將擴增所得的DNA進行測序。測序后判斷擴增DNA與NCBI中已有刀鱭物種是否同源:所得序列在NCBI進行blast比對,query coverage值為100%,說明擴增所得序列能夠被已有刀鱭序列完全覆蓋;E值為0,表示隨機分配可能性為0,進一步說明序列基本上完全匹配;indentity值為99%,表示該物種與其相似度為99%。綜上,可確定擴增所得序列是刀鱭的Cytb基因序列。
1.5 數據處理與分析
測序所得線粒體Cytb基因序列經clustal X軟件進行編輯、排序和手動校對。采用MEGA 3.1軟件計算序列堿基的組成、變異位點數、群體間和群體內的遺傳距離,并采用鄰接法(Neighbour-joining method,NJ)構建中間分化的系統樹。利用DnaSP 5.0軟件計算序列的單倍型數(H)、群體的單倍型多樣性 (Hd) 、群體核苷酸多樣性 (π)和平均核苷酸差異數(k)。應用Arlequin 3.1軟件計算群體間遺傳分化指數(F-statistics,Fst),并進行分子方差分析(AMOVA)和中性檢驗。
2 結果與分析
2.1 刀鱭群體遺傳多樣性
擴增的160條刀鱭線粒體Cytb基因序列經排序、校對和剪輯后,獲得160條長度為790 bp的序列片段。在所有序列中,A、T、G和C 4種堿基平均含量分別為27.7%、31.3%、14.3%和26.7%,G的含量最低;變異位點34個,約占所測序列片段的4.3%。在160個個體中共檢測到31個單倍型(表1),在所有單倍型中,7個單倍型為共享單倍型,其他24個單倍型為群體特有單倍型;共享單倍型中Hap2為所有群體共享,Hap4為銅陵-當涂(TD)、南京-鎮江(NZ)、崇明(CM)區段3個群體共享;除Hap2和Hap4外,其他單倍型在群體中出現頻率較低,均不超過2次。應用Arlequin 3.1軟件分析群體不變位點(ii)789個,轉換位點/顛換位點(R)的值為5.5。
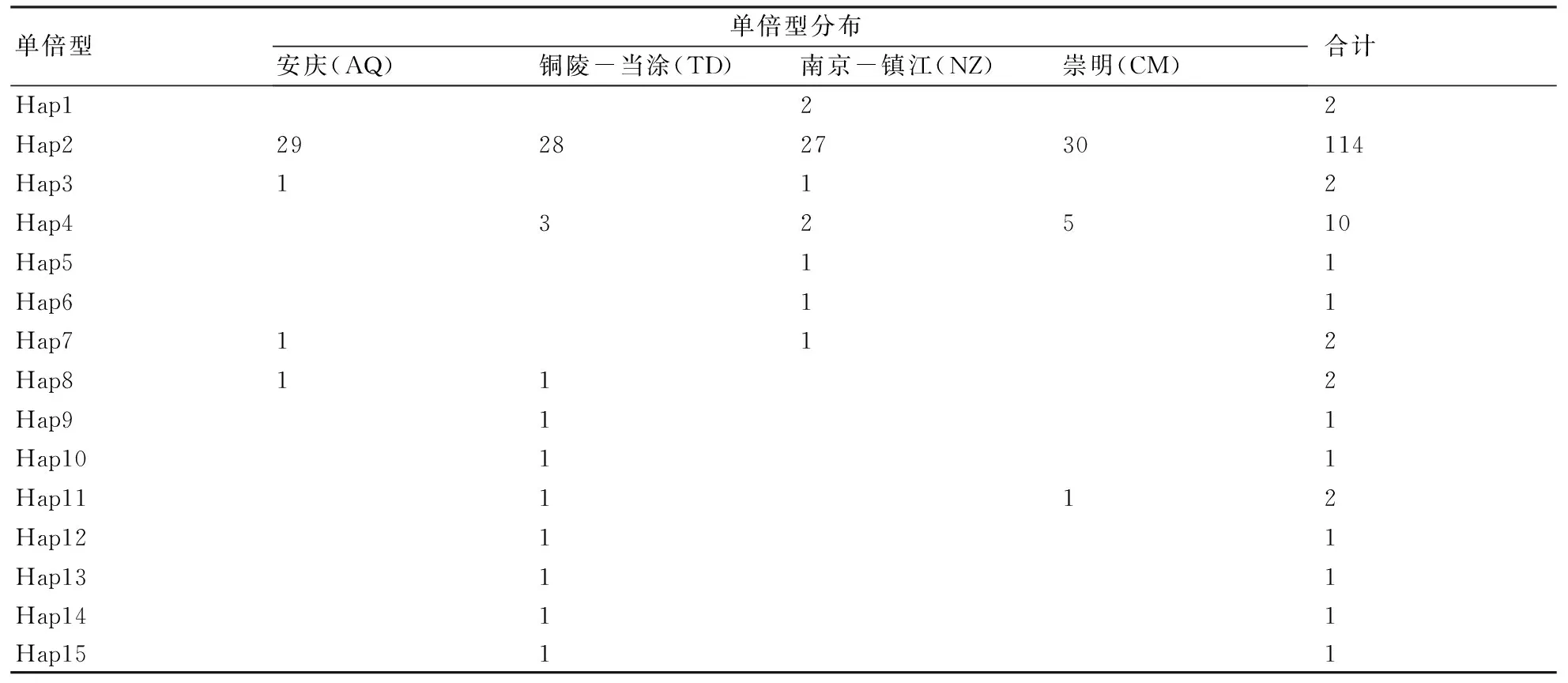
表1 長江刀鱭群體Cyt b序列單倍型分布特征
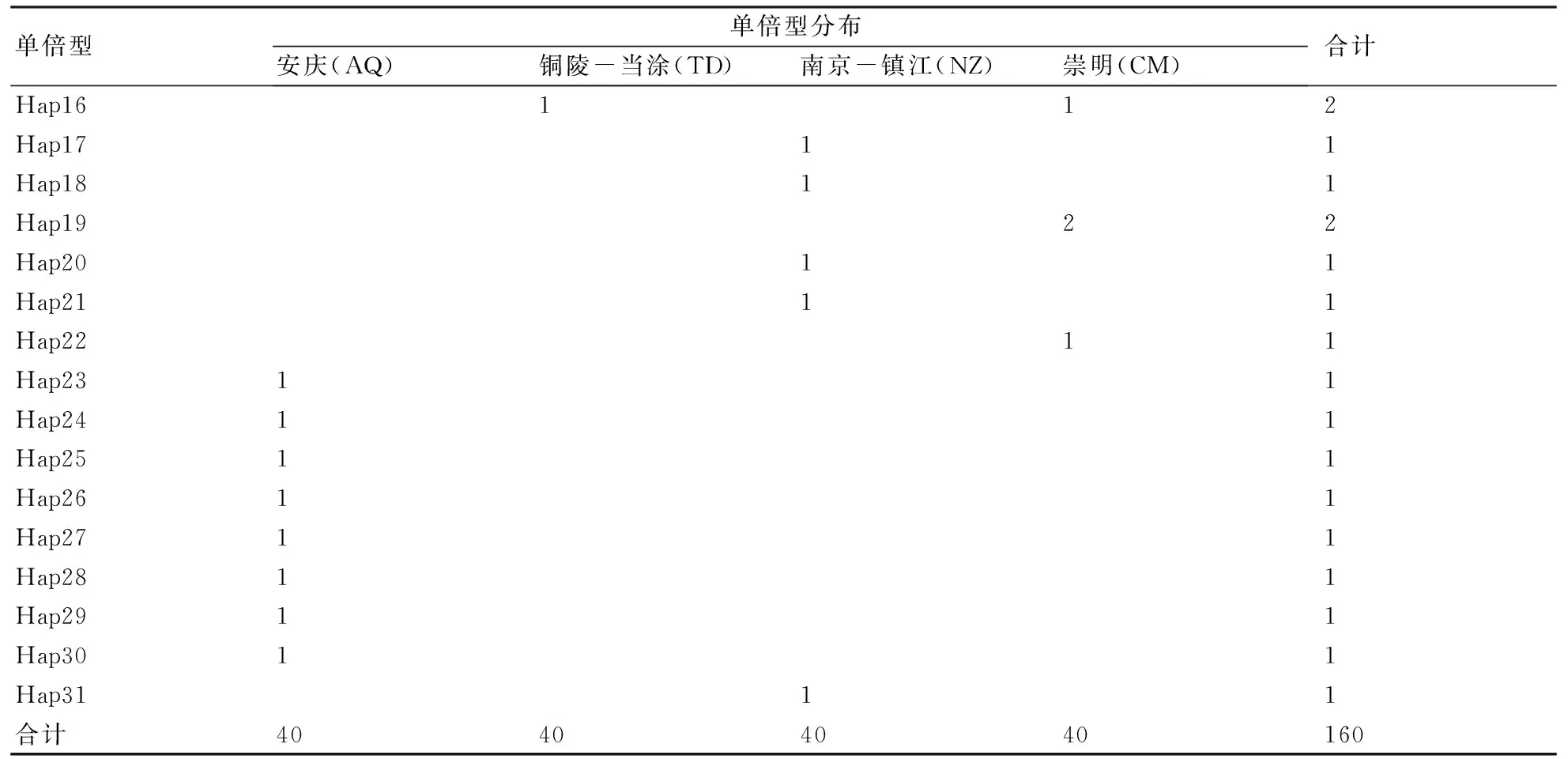
續表1:
160尾樣本分析顯示,單倍型多樣性指數為0.490±0.050,核苷酸多樣性指數為0.00078±0.00010(表2)。就單倍型結果而言,南京-鎮江(NZ)區段群體的單倍型多樣性指數較高,為0.547±0.096,崇明(CM)區段群體較低,為0.428±0.093;就序列而言,銅陵-當涂(TD)區段群體的核苷酸多樣性指數最高,為0.00094±0.00025,崇明(CM)區段群體最低,為0.00066±0.00018。就性別而言(表3),刀鱭雌性群體的單倍型多樣性指數和核苷酸多樣性指數均高于雄性群體,雌性群體與雄性群體相比,單倍型多樣性高出近55%,核苷酸多樣性高出65%。
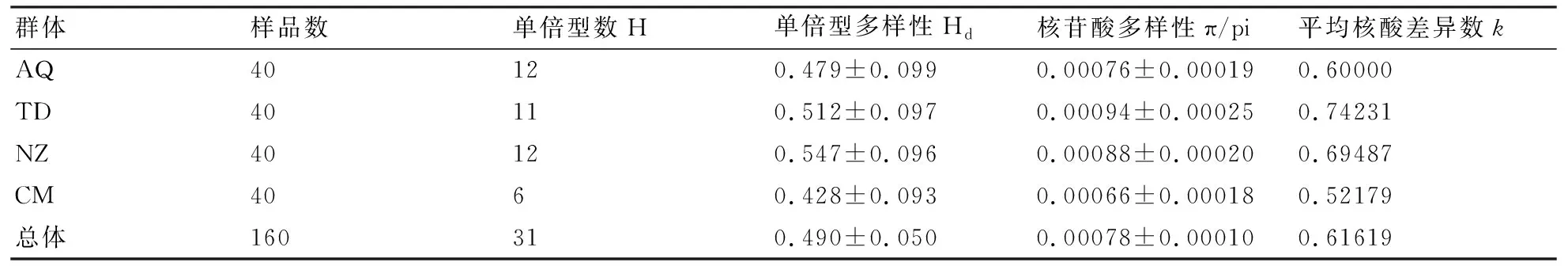
表2 長江刀鱭群體的樣品信息和遺傳多樣性參數
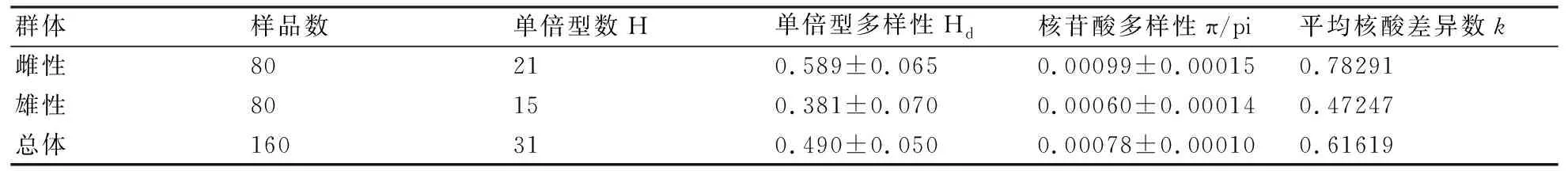
表3 長江刀鱭雌雄群體的遺傳多樣性參數
2.2 刀鱭群體遺傳結構
從長江刀鱭4個地理群體的遺傳距離(表4)來看,群體內的遺傳距離介于0.00060~0.00089,平均值為0.0007825;4個群體間的遺傳距離介于0.00070~0.00088,平均值為0.000785。此結果表明,長江刀鱭群體間的遺傳距離與群體內的遺傳距離非常接近,基本處于同一水平,無明顯遺傳分化。AMOVA結果顯示,長江刀鱭群體內遺傳變異為99.68%,群體間遺傳變異為0.32%(表5),整體遺傳分化系數(Fst)為0.00316,變異來源差異不顯著(P=0.20332),說明遺傳變異主要來自群體內。
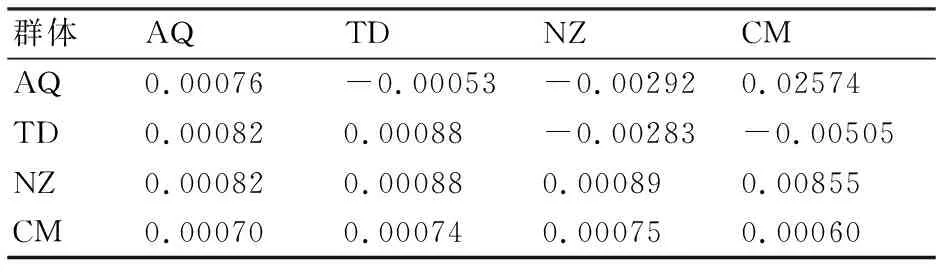
表4 長江刀鱭群體內遺傳距離(對角線)及遺傳分化系數(Fst)(上對角線)和兩兩群體間遺傳距離(下對角線)
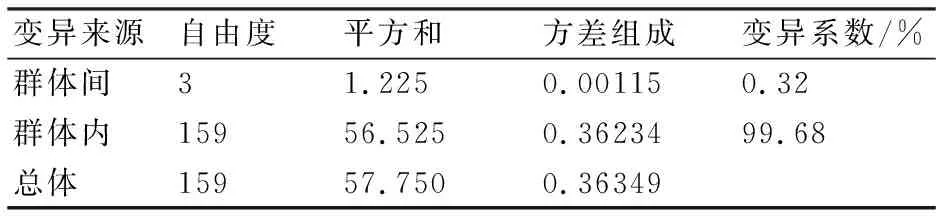
表5 基于Cyt b序列的長江刀鱭群體變異組成分析
在用鄰接法構建的系統進化樹中(圖2),單倍型Hap1~Hap13、Hap15~Hap25、Hap27~Hap31(n=158)聚為一支,單倍型Hap14和Hap26(n=2)聚為另一支,單倍型Hap14是蕪湖的雄性個體,單倍型Hap26是安慶的雌性個體。
2.3 刀鱭群體歷史動態
整體而言,Tijima’sD值和Fu’sFs值分別為-2.1543775和-8.6882225,均為負值且統計檢驗均達顯著水平(P<0.05)(表6)。4個長江刀鱭群體中性檢驗Tijima’sD值介于-2.41066~-1.52331,平均為-0.53859,Fu’sFs值介于-11.12077~-3.74634,平均為-2.17206,兩項指標差異均顯著(P<0.05)。
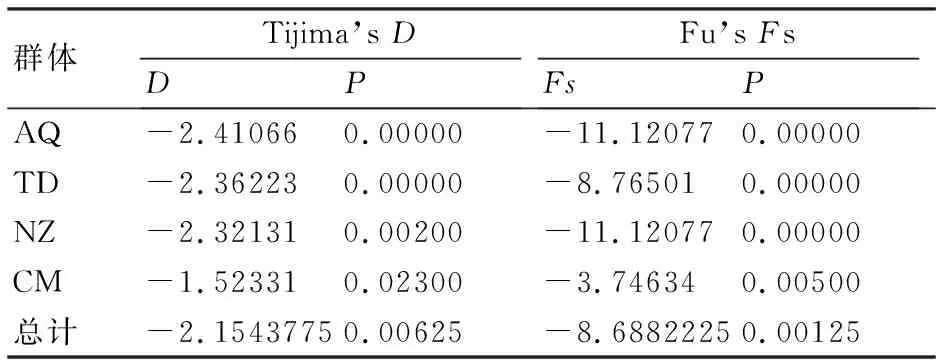
表6 長江刀鱭4個群體的中性檢驗結果
3 討論
3.1 刀鱭群體遺傳多樣性及歷史動態分析
遺傳多樣性亦稱為基因多樣性,從廣義上而言是指地球上生物所含遺傳信息總和。從狹義上而言,是指一個群體內不同個體的遺傳變異總和[21]。物種遺傳多樣性的高低與物種的生存能力、適應能力和進化潛力呈正相關,遺傳多樣性越高,物種對環境的適應能力就越強,蘊含著比較大的進化潛力和豐富的育種及遺傳改良能力,也越容易擴大其生存范圍;低水平的遺傳多樣性預示著適應能力較弱,有害隱形基因表達增加及經濟性狀衰退,最終導致物種退化。本研究獲得的基因序列中A+T的含量(59%)高于C+G的含量(41%),堿基組成不均一,具有較大偏向性,這與張燕萍等[4]、唐文喬等[22]的研究結果一致。單倍型多樣性和核苷酸多樣性是衡量物種遺傳多樣性的兩個重要指標[23]。馬春艷等[24]通過Shannon多樣性指數和Nei’s基因多樣性指數說明刀鱭群體具有較高的遺傳多樣性水平。本研究中上述兩項指標均高于2012年孫超[12]報道的日本刀鱭群體(Hd=0.400,π=0.000369),低于2008年楊金權等[5]報道的長江口刀鱭群體(Hd=0.9983,π=0.0262),呈現相對較低的遺傳多樣性水平。根據Grant等[25]提出了單倍型多樣性和核苷酸多樣性高低分別以0.5和0.005為臨界值,將種群進化模式分為了4個標準:單倍型多樣性和核苷酸多樣性均大于等于臨界值、單倍型多樣性和核苷酸多樣性均小于臨界值、單倍型多樣性大于等于臨界值且核苷酸多樣性小于臨界值、單倍型多樣性小于臨界值且核苷酸多樣性大于等于臨界值,并針對4個標準的出現分別作出了種群歷史狀態解釋。本研究中的刀鱭群體單倍型多樣性接近臨界水平(0.49≈0.5),核苷酸多樣性水平極低于臨界水平(0.00078<0.005),核苷酸多樣性積累所需時間比單倍型所需時間長[26],說明刀鱭可能是由一個較小的群體經過歷史擴張而來,擴張后的進化時間尚未達到累積核苷酸變異所需要的時間。根據Grant等[25]提出的種群進化模式,可推測本研究中刀鱭群體曾經歷過瓶頸效應和種群擴張。近年來的研究逐漸證實了這一點,加之人類活動的擾動,導致長江刀鱭群體遺傳多樣性水平表現出下降趨勢[29]。
3.2 刀鱭群體遺傳分化程度分析
種群間的遺傳距離以及種群分化指數是衡量種群多態程度的一個重要指標,值越大多態性程度就越高[28]。有研究表明,Cytb基因在種群間的基因遺傳距離一般水平范圍為0.005 - 0.015[23],本研究中的結果也介于該范圍,這說明長江刀鱭群體間遺傳距離極小,群體間基因交流頻繁,主要是因為其生存環境開放,個體間信息交換不受水域限制。同時,AMOVA分析結果顯示,長江刀鱭群體間遺傳分化系數較小,群體內遺傳變異較高(99.68%),而群體間遺傳變異較低(0.32%)。該研究結果表明長江刀鱭群體間的多態性較低,群體間的遺傳分化較小,遺傳變異主要來自群體內。推測開放的水域特征與棲息環境有助于長江刀鱭群體間遺傳信息的交流交換,是其遺傳分化較低的重要原因之一。
3.3 刀鱭群體系統進化樹及中性檢驗分析
在單倍型NJ系統進化樹中,31種單倍型聚為2支,對應單倍型顯示為Hap1~Hap13、Hap15~Hap25、Hap27~Hap31 (n=158)聚為一支,Hap14和Hap26聚為另一支,說明長江刀鱭群體中極少數個體可能發生了基因突變,也可能混雜著其他生態類型,但由于類似樣本量有限,因此,Hap14和Hap26對應單倍型還有待于進一步研究。楊金權等[29]曾通過中性檢驗和網絡親緣關系分析得出長江及其南部臨近水域的刀鱭群體約在更新世末期的0.17至0.13百萬年前有過種群的擴張歷史,并分析其可能是受到更新世末期海平面升降的影響。本研究中的中性檢驗結果均為負值且統計檢驗均達顯著水平,這說明長江刀鱭群體積累了較多的低頻突變,DNA進化偏離了中性選擇[30],指示長江刀鱭群體歷史上可能發生過種群擴張。本研究結果可進一步佐證楊金權等[29]的觀點,同時也與本文中單倍型多樣性和核苷酸多樣性分析后得出的結論一致。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
