新疆紅肉蘋果不同時期果皮花青苷含量變化及其合成相關基因表達
2021-09-08 01:19:14童盼盼張亞若王江波
江西農業學報 2021年8期
王 龍,王 芳,湯 蕾,童盼盼,張亞若,王江波,3*
(1.塔里木大學 植物科學學院,新疆 阿拉爾 843300;2.塔里木大學 南疆特色果樹高效優質栽培與深加工技術國家地方聯合工程實驗室,新疆 阿拉爾843300;3.新疆生產建設兵團 塔里木盆地生物資源保護利用重點實驗室,新疆 阿拉爾 843300)
紅肉蘋果(MalusniedzwetzkyanaDieck)為薔薇科蘋果屬果樹,是我國珍貴樹種,該果樹開花早,結果早,豐產,抗逆性強,其花、葉、果實等器官富含花青苷,且帶有不同程度的紅色[1]。新疆紅肉蘋果是一種果肉、果皮及果核均為紅色且擁有多種生物活性物質的獨特蘋果屬果實[2]。花青苷對果實顏色的形成有著重要的作用,花青苷是新疆紅肉蘋果中多酚的一種,屬黃酮類化合物[3]。
花青苷為蘋果體內重要的次生代謝產物,由葡萄糖代謝產生的苯丙氨酸為前體物質,一步步在各種酶的催化下合成而來[4]。花青苷的合成受到結構基因與轉錄因子的協同調控[5],其中部分對花青苷合成起調控作用的基因表達受環境因子的影響,比如溫度、光照和營養狀況[6-7]。前人對花青苷合成協同作用的結構基因的研究主要集中在與花青苷合成相關酶密切相關的結構基因ANS、UFGT、F3H、PYL、DFR、CHS和對結構基因具有調控作用的轉錄因子MYB、bHLH、TTG1等上[8-11]。
目前,有關紅肉蘋果花青苷合成所涉及到的酶和調控因子已有部分研究,但對于花青苷含量及其與相關基因表達量之間的關系尚未明確,因此,本研究以新疆紅肉蘋果克孜阿爾瑪、夏紅肉及國外引進紅肉品種紅色之愛為試材,測定了其果實在不同發育時期果皮花青苷含量及相關基因的表達量,旨在進一步了解花青苷合成機制,為培育紅肉蘋果新品種提供理論參考。
1 材料與方法
1.1 試驗材料
試驗材料選取輪臺南疆特色果樹資源圃的紅肉品種夏紅肉、克孜阿爾瑪,以及第一師阿拉爾市十二團從國外引進的紅肉品種紅色之愛,在盛花后30、60、90、120、150 d(紅色之愛在盛花后30 d未采集樣品)在樹冠外圍中部的4個不同方向隨機選取無病蟲害、大小基本相同的10個果實,各3次重復,用泡沫冰盒帶回實驗室,將果皮快速切成細小塊狀,用液氮處理后分別裝入提前準備好的實驗袋,放入-80 ℃超低溫冰箱保存備用。
1.2 試驗方法
1.2.1 花青苷提取及含量測定 花青素提取方法為pH示差法,并根據實際情況稍作調整[12]。取出部分所需樣品,快速用液氮研磨充分后準確稱取0.4 g,分別用2 mL的pH=1的緩沖液(含50 mmol/L KCl和150 mmol/L HCl)和pH=4.5的緩沖液(含400 mmol/L NaCH3CO2和240 mmol/L HCl)在低溫下抽提4 h,直至樣品基本呈白色;抽提完成后,在12000 r/min,4 ℃條件下離心15 min;取上清液測量其在510 nm的吸光度值(酶標儀)。花青素濃度計算公式為:花青素濃度(μg/g FW)=(ApH 1-ApH 4.5)×1000×484.8/24825×6。在本研究中0.4 g鮮樣近似為0.4 mL體積。
1.2.2 引物設計 在NCBI上查詢花青苷合成相關的候選基因,并參考An[13]和孫曉紅[14]等報道的基因序列,設計花青苷合成相關基因的實時熒光定量PCR(quantitative Real Time PCR, qRT-PCR)引物。引物名稱及序列見表1。

表1 qRT-PCR的引物
1.2.3 果皮RNA提取及qRT-PCR分析 按照上海生工SK8661柱式植物總RNA抽提純化試劑盒的說明書進行樣品果皮RNA的提取,實驗步驟稍作變動[19]。用NanoDrop One核酸蛋白分析儀測定OD260、OD280,計算檢測RNA的純度和濃度。同時將提取的RNA樣品分別加入溴酚藍,混勻后用含有EB的1.0%瓊脂糖凝膠電泳檢測RNA完整性,在紫外投射儀上觀察RNA分子的大小、完整性、電泳譜帶的清晰度;用反轉錄試劑盒HiScript?Ⅲ 1st Strand cDNA Synthesis Kit(+gDNA wiper)(cat.Z005163,Vazyme)合成cDNA,稀釋備用。
以Md-Actin作為qRT-PCR內參基因,對蘋果花青苷合成相關基因的相對表達量進行比較。定量PCR于美國ABI 型Q5熒光定量qPCR儀上進行,反應體系按SYBR?Green PCR Master Mix(cat. #RR820A,TaKaRa)說明書進行。PCR反應程序為:94 ℃預變性30 s;94 ℃ 5 s,60 ℃保持30 s,95 ℃延伸15 s,60 ℃ 1 min,95 ℃ 15 s,42個循環。在默認條件下讀取CT值。計算2-△△CT值,進行相對表達量分析,每個發育期樣品基因表達量為3種生物學重復的平均值。
1.2.4 數據統計分析 實驗所得數據用DPS數據處理系統和Excel進行整理、統計、分析和繪圖。
2 結果與分析
2.1 3種蘋果品種果實在不同發育時期果皮的花青苷含量變化
在果實不同發育時期果皮花青苷含量變化結果如圖1,在花后150 d時的外觀表型見圖2。紅肉蘋果夏紅肉果皮花青苷含量在盛花后30 d達到最高,其含量為135.732 mg/g,隨后開始下降,在90 d時降至最低,然后持續上升,在盛花后150 d時其花青苷含量再次有明顯升高;克孜阿爾瑪果皮花青苷含量變化趨勢與夏紅肉相同;紅色之愛果皮花青苷含量在盛花后60 d達到最高,其含量為88.840 mg/g,而后持續降低,在120 d時達到最低,在150 d時有所回升。在盛花后30 d時,夏紅肉與克孜阿爾瑪花青苷含量存在顯著差異;盛花后60 d,紅色之愛與夏紅肉、克孜阿爾瑪的花青苷含量存在顯著差異;盛花后90、120 d這3個品種之間互相存在顯著差異;盛花后150 d夏紅肉、紅色之愛與克孜阿爾瑪存在顯著性差異。
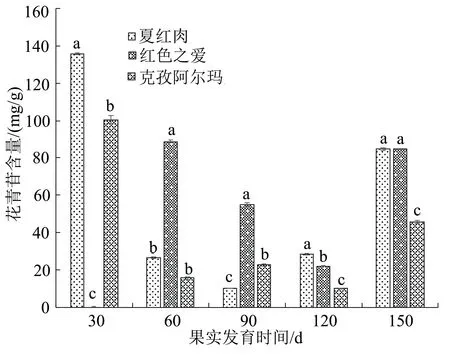
圖1 3種蘋果品種不同發育時期果皮花青苷含量

圖2 3種紅肉蘋果品種花后150 d時的外觀表型
2.2 3種蘋果品種果實不同發育時期果皮花青苷合成相關結構基因的相對表達量分析
對3種蘋果不同發育時期果皮花青苷合成結構基因進行qRT-PCR分析,結果(圖3)表明:在6個結構基因中F3H在3種蘋果盛花后90 d和120 d的表達量遠遠高于其他5個結構基因的表達量。結構基因CHI在夏紅肉果實發育的后4個時期表達量都高于其他品種,其次是紅色之愛、克孜阿爾瑪。結構基因CHS在各紅肉蘋果發育的各個時期表達量呈升高、降低再回升的趨勢,總體上,在盛花后60 d表達量達到最高,在盛花后120 d表達量最低。結構基因UFGT、DFR、F3H在紅色之愛盛花后120 d的表達量都大幅度高于其他各品種,同時這3個基因在夏紅肉盛花后90 d的表達量都高于其他時期,其中結構基因DFR在紅色之愛盛花后150 d表達量最高。結構基因F3H在夏紅肉盛花后90 d表達量最高,約為紅色之愛、克孜阿爾瑪的3~73倍。在6個結構基因中除UFGT、DFR、CHS外,其他3個基因在紅肉蘋果克孜阿爾瑪果實發育的后3個時期的表達量都低于其他兩種紅肉蘋果。
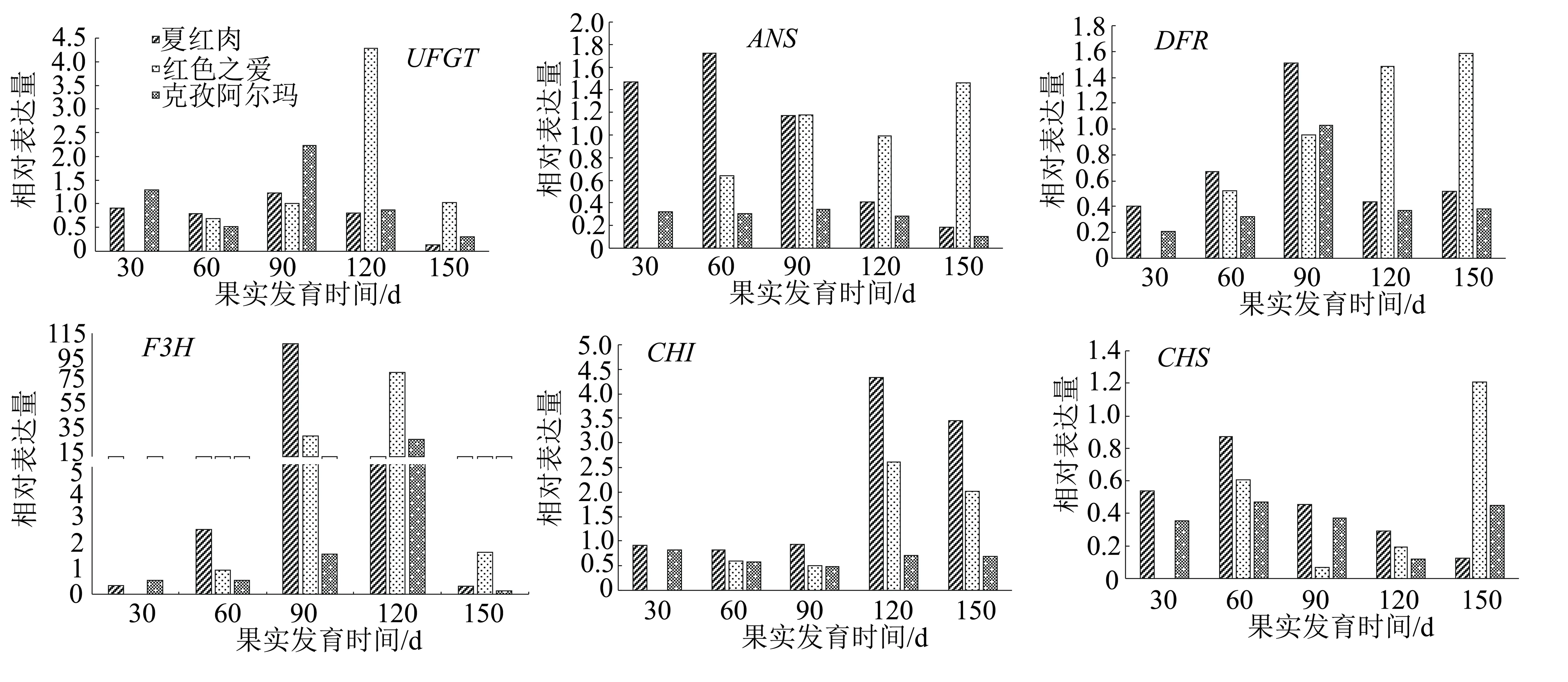
圖3 3種蘋果品種果實不同發育時期果皮花青苷合成結構基因的相對表達量
2.3 3種蘋果品種果實不同發育時期果皮花青苷生物合成轉錄因子表達量分析
對3種蘋果不同發育時期果皮花青苷合成轉錄因子進行qRT-PCR分析,結果(圖4)表明,轉錄因子MYB10的表達量遠高于其他轉錄因子。在各紅肉蘋果盛花后60 d與150 d轉錄因子bLHL3與TTG1的表達量整體低于其他3個時期,這兩種轉錄因子在紅色之愛盛花后120 d的表達量達到最高,大幅度高于其他品種和其它時期;紅色之愛盛花后90 d轉錄因子bHLH33的表達量達到最高且大幅度高于其他各品種。在克孜阿爾瑪與夏紅肉發育全部時期4種轉錄因子的變化趨勢都呈先降低再升高再降低的趨勢,在紅色之愛發育的各個時期呈先升高再降低的趨勢。
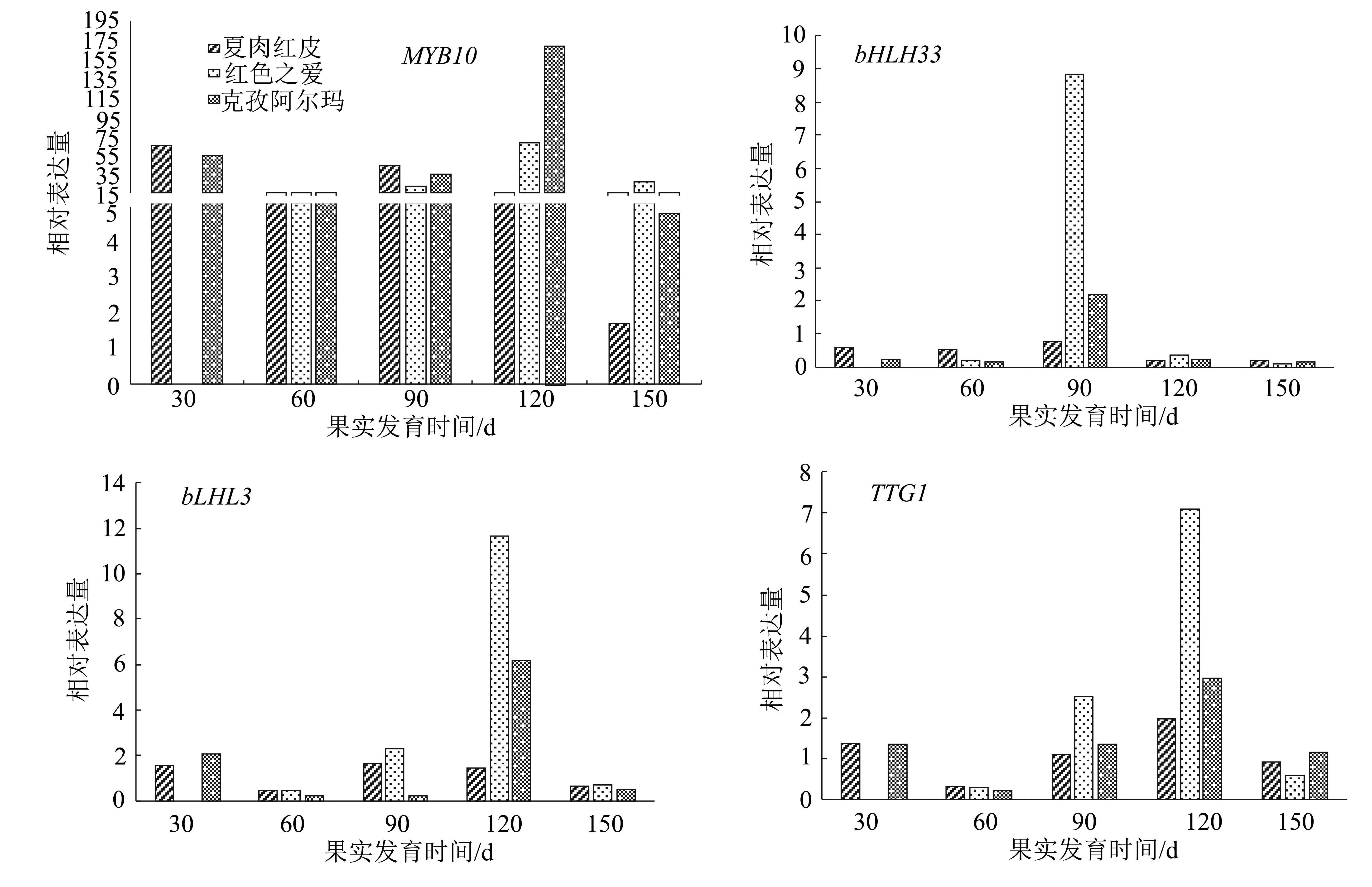
圖4 3種蘋果品種果實不同發育時期果皮花青苷合成相關轉錄因子的相對表達量
3 討論與結論
3.1 討論
花青苷作為植物重要的色素之一,對果皮色澤影響重大[15]。夏紅肉果皮和克孜阿爾瑪的果皮在盛花后30 d花青苷含量達到最高,其次是紅色之愛果皮在盛花后60 d花青苷含量達到最高;隨著果實的生長發育,各個品種的花青苷含量開始下降,下降至最低后持續上升,至150 d再次大幅度增高。孫曉紅[16]研究發現紅肉蘋果果實在盛花后30 d花青苷含量達到最高,隨著紅肉蘋果成熟度的增加果實中花青苷含量開始降低,但部分品種在120 d和150 d果實花青苷含量有所回升,這與本實驗結果基本一致。
花青苷在植物特定位置上的表達水平是由結構基因和調控基因共同控制的,經過試驗、數據等分析發現,新疆紅肉蘋果夏紅肉花青苷含量在盛花后150 d時有明顯升高,而結構基因在克孜阿爾瑪、夏紅肉盛花后120 d與150 d時表達量有明顯升高。結構基因除F3H外,其他基因UFGT、ANS、DFR、CHI、CHS的表達量在各個處理之間差異較小,結構基因F3H在夏紅肉盛花后90 d的表達量遠高于其他品種,在盛花后120 d在紅色之愛中的表達量遠高于其他品種。結構基因轉錄因子MYB10在克孜阿爾瑪和紅色之愛盛花后120 d的表達量遠高于其他品種。轉錄因子bHLH33在紅色之愛盛花后90 d的表達量遠高于其他品種;bLHL3在紅色之愛盛花后120 d的表達量達到最高。因此,紅色之愛果皮花青苷含量與結構基因F3H以及轉錄因子MYB10、bLHLH33的高水平表達有關,說明新疆紅肉蘋果花青苷的合成與其結構基因F3H、CHI關系密切,而其調控可能是由轉錄因子MYB10起主導作用,轉錄因子bHLH33和TTG1為主要協助,bHLH3轉錄因子作為次要協助。然而宋楊等[17]在研究中發現澤西越桔果皮中花青苷含量與CHS、F3H和UFGT的關系更為密切,在越桔花青苷合成過程中可能起更加重要的作用。而孫曉紅等[18]的研究結果認為導致紅勛5號和新疆1號紅肉蘋果花青苷含量高的主要原因可能是具有R6R6啟動子基因型和高水平表達的ANS、UFGT基因及MYB10、TTG1、bHLH33轉錄因子。前人對花青苷的研究結果都存在著多多少少的差異,也正是這些差異說明果皮紅色性狀的調控機制非常復雜,具體機制的明確則需要更專業更深入的研究和實踐。
3.2 結論
本試驗研究結果表明:夏紅肉果皮和克孜阿爾瑪果皮中的花青苷含量變化趨勢相同,在盛花后30 d花青苷含量均達到最高,然后持續下降至最低,在盛花后150 d其含量再次有明顯升高;紅色之愛果皮花青苷含量在盛花后60 d達到最高;結構基因F3H、CHI與轉錄因子MYB10、bLHLH33的高水平表達對花青苷合成有促進作用。
