褪黑素對脂多糖誘導的奶牛乳腺上皮細胞炎癥反應的緩解作用
2021-09-06 12:34:38李洪洋南雪梅
動物營養學報 2021年8期
李洪洋 南雪梅 孫 鵬
(中國農業科學院北京畜牧獸醫研究所,動物營養學國家重點實驗室,北京 100193)
奶牛乳房炎是一種高發性多因素疾病,生理狀態、營養水平、環境條件、管理制度等方面的差異均有可能引起奶牛乳房炎的發生,導致產奶量和乳品質受到影響,致使牧場經濟效益大幅降低[1]。病源微生物是引起奶牛患乳房炎的主要原因之一,常見的致病菌包括金黃色葡萄球菌、大腸桿菌和無乳鏈球菌,涵蓋革蘭氏陽性菌與革蘭氏陰性菌兩大類[2-3],它們分布于空氣、水體以及奶牛可接觸物體表面,極易導致奶牛罹患乳房炎。致病菌由乳頭導管入侵并進入乳腺組織,隨后引發感染和炎癥反應[4]。由于前期治療乳房炎的有效藥物較少,使得抗生素憑借顯著的抗菌效果受到廣泛應用。但隨著抗生素的大量使用,由其導致了耐藥性、藥物殘留以及由此引發的食品安全等問題,因此,亟需尋求更為安全有效且成本低廉的天然活性物質以替代抗生素。
褪黑素(melatonin,MT)是一種主要由松果體合成分泌的吲哚類激素[5],其在皮膚[6]、骨髓[7]、視網膜[8]、線粒體[9]亦可少量合成,在體內分布廣泛,成為重要的“同步信號”,可有效調控機體的生物節律[10]。隨著相關研究的深入,發現MT可發揮抗氧化[11]和免疫[12]等生物學效應。研究證實,MT及其多級代謝產物具有強大的抗氧化功能,是一種有效的自由基捕獲劑[13]。除自身強有力的抵抗自由基的活性外,MT還能夠通過促進抗氧化物酶的生成和增效抗氧化物酶的活性,從而發揮抗氧化效應[14]。研究表明,MT對免疫細胞具有增殖及增效作用,MT在谷胱甘肽和谷胱甘肽還原酶缺乏的情況下,可參與維持中性粒細胞的活性和功能[15]。MT與T細胞的關系也十分密切,現已證實T細胞內含有合成MT所需的4種酶類,并可在特定情況下高水平分泌。同時,MT對T細胞的激活和分化也存在調控功能,尤其是輔助性T細胞17(Th17)、調節性T細胞(Treg)和記憶T細胞[16]。近年來,除MT清除體內自由基、促進免疫應答的功能外,其抗炎效應亦受到廣泛關注[17]。因此,本試驗旨在通過脂多糖(lipopolysaccharide,LPS)誘導奶牛乳腺上皮細胞(bovine mammary epithelial cells,BMECs)炎癥反應模型,探究MT對LPS引起的BMECs炎癥反應的緩解作用及潛在機制,為應用MT防治奶牛乳房炎提供科學依據。
1 材料與方法
1.1 試驗材料
BMECs獲贈于北京畜牧獸醫研究所智慧畜牧業創新團隊,液氮保存;LPS[大腸桿菌(E.coli)O55∶B5]購自美國Sigma公司;MT購自上海易恩化學技術有限公司;胎牛血清(fetal bovine serum,FBS)、DMEM+GlutaMAX培養基、F-12培養液、D-Hank’s平衡鹽溶液(Hank’s balanced salt solution,HBSS,不含鈣、鎂)、0.05%胰蛋白酶(trypsin)-乙二胺四乙酸(EDTA)消化液和Pen Strep雙抗混合液購自美國Thermo Fisher公司;腫瘤壞死因子-α(tumor necrosis factor,TNF-α)、白細胞介素-6(interleukin 6,IL-6)和白細胞介素-1β(interleukin 1β,IL-1β)的酶聯免疫吸附測定(ELISA)試劑盒購自上海瑞番生物科技有限公司;四甲基偶氮唑藍[3-(4,5)-dimethylthiahiazo(-z-y1)-3,5-di-phenytetrazoliumromide,MTT]細胞增殖及細胞毒性檢測試劑盒購自北京酷來博科技有限公司,Western blot相關試驗材料購自上海碧云天公司;一抗β-肌動蛋白(β-actin)和核因子-κB同源蛋白(P65)購自美國Immuno Way公司,Toll樣受體4(TLR4)購自美國Santa Cruz公司,核因子-κB抑制蛋白(IκB-α)購自美國CST公司;二抗購自美國Jackson ImmunoResearch公司。
1.2 試驗方法
1.2.1 BMECs的培養
BMECs復蘇后,使用完全培養基(DMEM/F12,含10% FBS和1%雙抗)于37 ℃、5%二氧化碳(CO2)培養,傳3~8代后用于后續試驗[18]。
1.2.2 LPS炎性模型構建
1.2.2.1 LPS處理液的配制
將10 mg LPS溶解于10 mL HBSS(含1%雙抗)配制1 mg/mL的LPS儲存液,分裝后-20 ℃儲存。使用時取1 mL儲存液溶于9 mL的無血清DMEM/F12培養基(含1%雙抗)配制100 μg/mL處理母液,再通過稀釋添加不同體積無血清DMEM/F12培養基(含1%雙抗)進一步配制濃度分別為0.1、0.5、1.0、5.0和10.0 μg/mL的LPS處理液,現用現配[10]。
1.2.2.2 LPS誘導BMECs炎癥反應模型
將BMECs按1×104個/孔接種于96孔板,每孔加入基礎培養基200 μL,邊緣孔各加入200 μL HBSS,培養24 h后,更換無血清DMEM/F12培養基(含1%雙抗)培養12 h。隨后每列分別更換為不同濃度(0、0.1、0.5、1.0、5.0和10.0 μg/mL)的LPS處理液100 μL,繼續分別培養6和12 h,每個濃度設6個重復。預試驗中,與0 μg/mL組相比,細胞活性和炎性因子水平出現顯著差異視為炎癥誘導成立[19]。
1.2.2.3 MTT檢測法
按照MTT細胞增殖及細胞毒性檢測試劑盒說明書,避光條件下每孔加入20 μL MTT溶液,培養處理4 h;取出后每孔加入100 μL二甲基亞砜(DMSO)終止反應,搖床10 min后,用酶標儀(SpectraMax M5,美國Molecular Devices公司)在570 nm處測定吸光度。
1.2.2.4 炎性細胞因子含量檢測
按ELISA測定試劑盒說明書進行TNF-α、IL-6、IL-1β含量檢測,用酶標儀在450 nm處測定吸光度。
1.2.3 MT對BMECs活性的影響
1.2.3.1 MT處理液的配制
稱取0.023 2 g MT溶解于1 mL乙醇中[10],再將溶解于乙醇的MT樣品溶解于99 mL無血清DMEM/F12培養基(含1%雙抗),配制濃度為1 mmol/L的MT儲存液,以10 mL為單位分裝后-20 ℃儲存。使用時10 mL儲存液1∶9稀釋于無血清DMEM/F12培養基(含1%雙抗)配制處理母液,再用無血清DMEM/F12培養基(含1%雙抗)進一步配制濃度分別為1、5、10、50和100 μmol/L的MT處理液,現用現配。
1.2.3.2 MT對BMECs活性的影響
將BMECs按1×104個/孔接種于96孔板,每孔加入基礎培養基200 μL,邊緣孔各加入200 μL HBSS,培養24 h后,更換DMEM/F12培養基(不含FBS,含1%雙抗)培養12 h。隨后每列分別更換為不同濃度(0、1、5、10、50和100 μmol/L)的MT處理液100 μL,繼續分別培養48 h。每個濃度設6個重復,利用MTT法測定細胞活性。
1.2.4 MT對LPS誘導的BMECs炎癥反應的影響
試驗共分為7組,對照組(C組)BMECs不進行LPS誘導和MT處理,LPS組BMECs進行LPS誘導12 h,LPS+MT組BMECs進行LPS誘導12 h后再經不同濃度(1、5、10、50和100 μmol/L)MT處理48 h。每組設4個重復。
1.2.5 炎性細胞因子含量檢測
按ELISA測定試劑盒說明書對各組的細胞培養基進行炎性細胞因子(TNF-α、IL-6、IL-1β)含量檢測,使用酶標儀于450 nm處測定其吸光度值。
1.2.6 Western blot檢測炎癥反應相關蛋白的表達
使用HBSS清洗各組BMECs樣本。在細胞培養皿中添加細胞裂解液,用細胞刮刀刮取細胞后,將懸液移至新的做好標記的1.5 mL離心管中放在冰上進行充分裂解。4 ℃、12 000 r/min離心10 min后,將樣品的上清液移入離心管中。測定蛋白濃度,并調整蛋白濃度一致為4.0 μg/μL。隨后制備十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳凝膠,待膠體凝后將膠板組裝于電泳槽中,倒滿電泳緩沖液依次加樣20 μg/孔。隨后120 V、15 min和200 V、45 min進行電泳。電泳結束后采用三明治濕轉法,用提前配制好的轉膜液進行轉膜。隨后,對膜進行封閉和一抗、二抗孵育,置于全自動化學發光儀(Tanon,5200)中顯色后,使用Image J分析軟件V1.8.0分析條帶的灰度值。
1.3 數據處理與分析
試驗數據采用Excel 365進行整理,應用SPSS 19.0軟件進行統計,MTT法和Western blot檢測的試驗結果進行獨立t檢驗,ELISA法的試驗結果進行單因素方差分析(one-way ANOVA),并采用Duncan氏法進行多重比較。P<0.05代表差異顯著,P<0.01代表差異極顯著。試驗結果用平均值±標準差(mean±SD)表示。
2 結果與分析
2.1 LPS誘導BMECs炎癥細胞模型
預試驗中,利用MTT法測定細胞活性和ELISA法測定TNF-α、IL-6和IL-1β含量,確定LPS誘導細胞炎癥模型的處理濃度為10 μg/mL,處理時間為12 h。
2.2 MT對BMECs活性的影響
不同濃度(1、5、10、50和100 μmol/L)的MT作用于BMECs 48 h后,采用MTT法測定細胞活性,結果如圖1所示,不同濃度的MT處理對BMECs活性無顯著影響(P>0.05),其中1和5 μmol/L MT組BMECs活性較高。

數據柱標注不同小寫字母表示差異顯著(P<0.05),相同或無字母表示差異不顯著(P>0.05)。圖2同。
2.3 MT對LPS誘導的BMECs炎性因子含量的影響
如圖2所示,LPS誘導的BMECs中TNF-α、IL-6和IL-1β含量升高,其中,LPS組BMECs中IL-6和IL-1β含量顯著高于對照組(P<0.05),LPS組BMECs中TNF-α含量與對照組差異不顯著(P>0.05)。BMECs經MT處理48 h后,與LPS組相比,10和50 μmol/L MT組BMECs中TNF-α含量顯著降低(P<0.05),1、5、10和100 μmol/L MT組BMECs中IL-6含量顯著降低(P<0.05),10 μmol/L MT組BMECs中IL-1β含量顯著降低(P<0.05)。綜上所述,10 μLPS組的MT對LPS誘導的BMECs炎性因子含量升高的抑制效果最顯著。
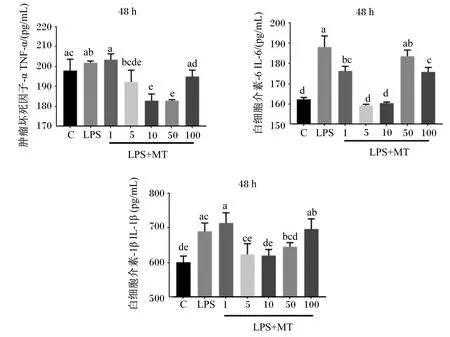
C代表對照組;LPS代表LPS組;LPS+MT代表LPS+MT組;1、5、10、50、100分別代表1、5、10、50、100 μmol/L的MT。下圖同。
2.4 MT對LPS誘導的BMECs中TLR4、P65和IκB-α蛋白表達量的影響
如圖3所示,與對照組相比,LPS組BMECs中TLR4蛋白表達量有所升高(P>0.05);與LPS組相比,不同劑量MT處理的BMECs中TLR4蛋白表達量均有所下降,其中50和100 μmol/L MT組差異顯著(P<0.05)。與對照組相比,LPS組BMECs中P65蛋白表達量顯著降低(P<0.05);與LPS組相比,不同劑量MT處理的BMECs中P65蛋白表達量均有所升高,其中1和50 μmol/L MT組差異顯著(P<0.05),10 μmol/L MT組差異極顯著(P<0.01)。與對照組相比,LPS組BMECs中P65蛋白表達量顯著降低(P<0.05);與LPS組相比,不同劑量MT處理的BMECs中IκB-α蛋白表達量均有所升高,其中1、10和50 μmol/L MT組差異顯著(P<0.05),5 μmol/L MT組差異極顯著(P<0.01)。

數據柱標注*表示與LPS組相比差異顯著(P<0.05),標注**表示與LPS組相比差異極顯著(P<0.01)。
3 討 論
3.1 MT對LPS誘導的BMECs炎癥反應的影響
近年來,相繼發現MT的抗氧化、免疫和抗炎等生物學效應[11-12,17],利用MT作為抗炎藥物緩解奶牛乳房炎不失為一種可嘗試的新思路。LPS作為大腸桿菌外膜的組成成分,利用其誘導BMECs建立體外炎癥模型在奶牛乳房炎的防治研究中被廣泛應用[18]。因此,本試驗旨在探究MT對LPS誘導的BMECs炎癥反應的影響。本試驗結果顯示,MT能夠降低LPS誘導的BMECs中炎性細胞因子含量,抑制TLR4蛋白表達量,抑制P65蛋白表達量的降低,升高IκB-α蛋白表達量,表明本試驗中MT對LPS誘導的BMECs的炎癥反應具有緩解作用。
3.2 MT對LPS誘導的BMECs中炎性細胞因子含量的影響
研究表明,體內細胞因子含量的變化是誘發機體炎癥反應的主要原因[20]。TNF-α、IL-1β和IL-6作為炎性細胞因子,在體內具有重要的信息傳遞作用[21]。它們與細胞中含有的特異性受體結合后,通過不同的信號傳導通路激發機體的免疫應答反應。現有研究發現,MT進行預處理的Caco-2細胞再經IL-1β炎癥誘導時,其炎性因子IL-6含量與對照組相比有所降低,表明MT對炎癥反應具有預防作用[22]。由于乳房炎屬于一種防治兼顧的炎癥疾病,因此本試驗重點研究其對BMECs炎癥反應的治療功效。本試驗中測定了MT對LPS誘導12 h后BMECs培養上清液中TNF-α、IL-1β和IL-6含量的影響,發現經LPS誘導12 h后BMECs培養上清液中TNF-α、IL-1β和IL-6含量顯著升高,加入MT后顯著抑制了以上炎性細胞因子含量的升高,且基于這3種炎性細胞因子的綜合抑制效果發現,10 μmol/L MT的作用效果最佳,表明MT對LPS誘導的BMECs炎癥反應具有緩解作用。與本試驗結果相似,Mishra等[23]利用高濃度乙醇誘導小鼠肝臟組織損傷后,發現肝細胞中TNF-α、IL-1β和IL-6含量顯著升高,添加MT后得到有效抑制。Naveen等[24]研究發現,支氣管炎、鼻炎、肺纖維化及哮喘等疾病發生時,炎性細胞因子含量大幅升高,MT通過細胞外信號調節激酶1/2(ERK1/2)和磷脂酰肌醇3-激酶(PI3K)/蛋白激酶B(Akt)信號通路抑制了肺部成纖維細胞中炎性細胞因子的過度表達,因此MT對BMECs中炎性細胞因子的抑制亦與信號通路密切相關。
3.3 MT對LPS誘導的BMECs中TLR4、P65和IκB-α蛋白表達的影響
研究發現,TLR4可識別LPS并通過信號傳導激活核因子-κB(NF-κB)[25]。NF-κB是一種轉錄因子,廣泛參與炎癥反應和先天性免疫應答等細胞活動。正常生理狀態下,NF-κB與NF-κB抑制蛋白(IκB)結合,以無活性的NF-κB/IκB復合物形式存在于細胞質中。LPS作為一種外源性激活劑,能夠刺激IκB激酶活化,導致IκB發生磷酸化并被降解,引發NF-κB/IκB復合物解聚,從而使NF-κB活化并被釋放進入細胞核內,與特定基因的啟動位點結合并調控相關基因的轉錄和翻譯。研究表明,MT可通過抗氧化級聯反應清除過量的自由基,抑制細胞因子,從而調控NF-κB介導的炎癥反應[26],MT也能夠直接調控NF-κB激發的炎癥信號通路[27]。因此,MT對于BMECs炎癥反應的緩解作用與NF-κB信號通路密切相關。本試驗中,MT能夠影響LPS誘導的BMECs中TLR4、P65和IκB-α蛋白表達量。MT處理BMECs 48 h后,不同劑量MT均能夠抑制TLR4蛋白表達量,且50和100 μmol/L MT組差異顯著,表明MT可能通過TLR4調控NF-κB介導的炎癥反應。不同劑量MT均抑制了LPS引起的P65蛋白表達量的降低,使其趨于對照組的正常水平,其中10 μmol/L MT組效果最顯著。與LPS組相比,不同劑量MT對IκB-α蛋白表達量均呈現促進效果,其中5 μmol/L MT組差異極顯著,表明MT通過促進IκB-α蛋白的表達,使NF-κB以無活性的復合物形式存在,從而抑制NF-κB引起的炎癥反應。Shao等[28]研究發現,MT有效抑制LPS誘導的小鼠乳腺上皮細胞中TLR4蛋白表達量,P65和IκB蛋白的磷酸化水平被顯著抑制,P65和IκB蛋白表達量有所升高,與本試驗結果一致。綜上所述,MT通過調控TLR4、P65和IκB-α的蛋白表達參與NF-κB炎癥通路,緩解LPS誘導的BMECs炎癥反應。
4 結 論
① MT能夠降低LPS誘導的BMECs中TNF-α、IL-1β和IL-6含量,且10 μmol/L MT組作用效果最佳。
② MT能夠抑制LPS誘導的BMECs中TLR4蛋白表達量的升高,并對P65和IκB-α蛋白表達量具有調控作用,MT可能通過參與NF-κB炎癥通路緩解LPS誘導的BMECs炎癥反應。
