褐藻寡糖對鰱魚魚糜穩定性、分子間作用力及肌原纖維蛋白結構的影響
2021-09-05 02:38:42楊天耿文豪鄭志紅汪秋寬陳勝軍劉麗叢海花
肉類研究 2021年7期
楊天 耿文豪 鄭志紅 汪秋寬 陳勝軍 劉麗 叢海花


摘 要:為區別不同質量濃度(0.5、1.0 g/100 mL)褐藻寡糖,包括酸解甘露糖醛酸寡糖(acidolyzed mannose oligosaccharide,AMO)、酸解古羅糖醛酸寡糖(acidolyzed guluronic oligosaccharide,AGO)和酶解褐藻酸鈉寡糖(enzymolysis alginate oligosaccharide,EAO)對鰱魚魚糜及肌原纖維蛋白結構的影響,對鰱魚魚糜熱穩定性、分子間作用力以及肌原纖維蛋白傅里葉變換紅外光譜、內源熒光光譜及紫外吸收光譜進行分析。結果表明:AMO質量濃度對鰱魚魚糜熱穩定性作用無規律,但在總體上,EAO提高了魚糜的熱穩定性,且與EAO質量濃度呈正相關,低質量濃度AMO與AGO更容易降低魚糜的冰點和可凍結水含量,在提高魚糜的抗凍性方面性能更優越;1.0 g/100 mL AGO保護離子鍵效果最好,EAO保護了肌原纖維蛋白的離子鍵、降低了氫鍵含量;褐藻寡糖修飾后,所有魚糜-褐藻寡糖復合物的β-轉角、α-螺旋含量減少,β-折疊、無規則卷曲含量上升;3 種褐藻寡糖對肌原纖維蛋白三級結構的展開有不同影響。
關鍵詞:鰱魚魚糜;肌原纖維蛋白;褐藻寡糖;穩定性;分子間作用力;蛋白質結構
Abstract: This study compared the effects of different concentrations (0.5 and 1.0 g/100 mL) of acidolyzed mannose oligosaccharide (AMO), acidolyzed guluronic oligosaccharide (AGO) and enzymolysis alginate oligosaccharide (EAO) on the thermal stability, intermolecular forces, Fourier transform spectra, endogenous fluorescence spectra and UV absorption spectra of myofibrillar proteins in silver carp surimi. The results showed that AMO affected the thermal stability of silver carp surimi irrespective of its concentration. However, on the whole, EAO increased the thermal stability of surimi, and this effect was positively correlated with EAO concentration. Low concentrations of AMO and AGO could more readily reduce the freezing point and freezable water content of surimi, and have stronger ability to increase the freezing resistance. AGO at 1.0 g/100 mL had the best protective effect on ionic bonds, and EAO was able to protect ionic bonds of myofibrillar proteins and reduce the hydrogen bond content. After oligosaccharide modification, the contents of β-turn and α-helix of myofibrillar proteins decreased, while the contents of β-sheet and random coil increased. The three oligosaccharides had different effects on tertiary structure unfolding of myofibrillar proteins in silver carp surimi.
Keywords: silver carp surimi; myofibrillar protein; alginate oligosaccharides; stability; intermolecular force; protein structure
DOI:10.7506/rlyj1001-8123-20210428-116
中圖分類號:TS254.4 ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? 文獻標志碼:A 文章編號:1001-8123(2021)07-0001-08
引文格式:
楊天, 耿文豪, 鄭志紅, 等. 褐藻寡糖對鰱魚魚糜穩定性、分子間作用力及肌原纖維蛋白結構的影響[J]. 肉類研究, 2021, 35(7): 1-8. DOI:10.7506/rlyj1001-8123-20210428-116. ? ?http://www.rlyj.net.cn
YANG Tian, GENG Wenhao, ZHENG Zhihong, et al. Effects of alginate oligosaccharides on the stability, intermolecular forces and myofibrillar protein structures of silver carp surimi[J]. Meat Research, 2021, 35(7): 1-8. DOI:10.7506/rlyj1001-8123-20210428-116. ? ?http://www.rlyj.net.cn
我國是淡水魚養殖大國,鰱魚(Hypophthalmichthys molitrix)是四大淡水魚之一,2019年鰱魚年產量已達到381 萬t[1],在水產品加工領域占有重要地位,鰱魚以原料價格低和產量大的優勢近年來受到廣泛關注,已成為魚糜制品的潛在原料[2]。2019年我國魚糜制品產量已達到139.4 萬t[1]。鰱魚肌原纖維蛋白是魚肉的重要組成部分,也是魚肉中最主要的蛋白,在肉類的加工貯存中起到非常重要的作用[3]。抑制或減少魚糜肌原纖維蛋白在貯藏和加工過程中的變性可以改善魚糜制品的功能特性,生產高品質產品。
常用物理、化學、酶和基因工程等方法修飾蛋白質。化學法主要包括酰化、磷酸化、脫氨基、糖基化改性等,其中糖基化改性以操作條件溫和、加工安全性高及賦予蛋白質新功能等優勢受到重視[4]。研究發現,酶解褐藻酸鈉寡糖(enzymolysis alginate oligosaccharide,EAO)可提高肌原纖維蛋白在低NaCl濃度溶液中的溶解度[5]和保水性能[6];山梨糖醇、山梨糖醇與海藻糖/蔗糖復合,可抑制肌原纖維蛋白的聚集[7];經葡萄糖糖基化改性后具有高溶解度的肌原纖維蛋白熱穩定性顯著提高[8];右旋糖酐[9]和葡萄糖[10]可提高肌原纖維蛋白的乳化性。
褐藻酸鈉的分子式為(C6H7O6Na)n,相對分子質量為32 000~200 000,其結構單元相對分子質量理論值為198.11[11-13],在pH>12時呈膠體狀態,在pH<3時形成不溶性凝膠,由β-D-甘露糖醛酸(M)和α-L-古羅糖醛酸(G)通過1→4糖苷鍵以3 種方式(MM、GG和GM)連接成直鏈線性高聚合物[14]。褐藻寡糖具有良好的溶解度、生物利用度和多重生物活性[15],如促進植物生長[16]、保護神經元[17]、抗氧化[18]、免疫調節[19]、抗炎、抗腫瘤[20-21]等。褐藻寡糖是低分子低聚物,通常用酸水解、氧化和生物酶水解的方法降解褐藻酸鈉獲得。酸水解法可分別使用草酸、鹽酸、硫酸或甲酸等降解獲得聚甘露糖醛酸(MM)和聚古羅糖醛酸(GG)[22],進一步降解獲得酸解甘露糖醛酸寡糖(acidolyzed mannose oligosaccharide,AMO)、酸解古羅糖醛酸寡糖(acidolyzed guluronic oligosaccharide,AGO),但酸法降解的反應耗時長,一般需4~12 h,反應條件不易控制,能耗高,產物回收率低,且“三廢”污染嚴重,產品外觀色澤差[22]。酶解法條件溫和、可控性強,生產的EAO特異性高[23]。
冷凍魚糜是魚糜制品加工行業主要的流通料[24],淡水魚糜在冷凍貯藏和加工過程中需要更好地保存品質,褐藻寡糖可作為冷凍海產品原料的抗凍劑。本研究系統考察不同來源褐藻寡糖(酸解法分別獲得AMO和AGO,酶解法獲得EAO)對冷凍鰱魚魚糜的穩定作用及對肌原纖維蛋白分子結構的影響,為進一步研究冷凍、解凍等加工過程和消化過程中蛋白質變化提供參考。
1 材料與方法
1.1 材料與試劑
冷凍鰱魚糜,2019年6月12日購于洪湖市井力水產食品股份有限公司。AMO、AGO ? 實驗室自制;EAO 中科榮信生物科技有限公司;溴化鉀(光譜純) 天津市大茂化學試劑廠;氯化鈉(分析純) 遼寧泉瑞試劑有限公司;氫氧化鈉(分析純) 天津市東麗區天大化學試劑廠;乙醇(分析純) 天津市北聯精細化學品開發有限公司;尿素(分析純) 國藥集團化學試劑有限公司;β-巰基乙醇(分析純) 上海雅吉生物科技有限公司;三羥甲基氨基甲烷(Tris)(分析純) 青島捷世康生物科技有限公司;食鹽 濰坊千源鹽化有限公司。
1.2 儀器與設備
DS-1高速組織搗碎機 上海標本模型廠;HR/T20MM立式高速冷凍離心機 湖南赫西儀器裝備有限公司;SynergyH1/H1M酶標儀 美國伯騰儀器有限公司;Q20差示掃描量熱(differential scanning calorimetry,DSC)儀 美國TA儀器公司;F-2700熒光分光光度計日本Hitachi公司;NEXUS670傅里葉變換紅外光譜(Fourier-transform infrared spectroscopy,FTIR)儀美國PerkinElmer公司;UV-9000雙光束紫外-可見分光光度計 上海元析儀器有限公司。
1.3 方法
1.3.1 酸解法制備AMO、AGO
參考趙丹等[25]的方法略作修改,采用酸水解法制備AMO、AGO。取2 g褐藻酸鈉,加入100 mL蒸餾水浸泡24 h,加入80 mL 0.3 mol/L鹽酸,沸水回流水解10 h,冷卻后5 000×g離心15 min,取沉淀溶于60 mL 0.8 g/100 mL Na2CO3溶液,用0.5 mol/L鹽酸調節pH值至2.85,5 000×g離心15 min得上清液與沉淀2,在上清液中加入2 倍體積乙醇(體積分數95%),5 000×g離心15 min后得沉淀1,沉淀1和沉淀2分別用體積分數75%乙醇和體積分數95%乙醇反復洗滌,再將2 種沉淀冷凍干燥,分別得到AMO和AGO。
1.3.2 魚糜-褐藻寡糖復合物的制備
參考楊姣等[26]的方法略作修改。將冷凍鰱魚魚糜于4 ℃層析柜中解凍過夜,取20 g魚糜,加入1.5%食鹽,擂潰5 min,添加10 mL 0.5 g/100 mL或1.0 g/100 mL的AMO、AGO、EAO溶液,將魚糜水分含量調整為76%,以添加10 mL蒸餾水的樣品為空白,繼續擂潰2 min使水分與魚糜充分混勻,得到添加不同質量濃度3 種褐藻寡糖的魚糜-褐藻寡糖復合物。
1.3.3 DSC法測定復合物穩定性
參考黃海[27]的方法。稱取13~15 mg魚糜-褐藻寡糖復合物,放入氧化鋁坩堝底部密封,使用標準品對DSC儀進行校準后進行樣品測定,以密封的空坩堝為對照。實驗采用的測定程序為:初始溫度與結束溫度設定為15 ℃,先以10 ℃/min降溫到-50 ℃并保持10 min,再以5 ℃/min升溫到15 ℃保持1 min,以5 ℃/min升溫到100 ℃后以50 ℃/min降溫到15 ℃。
在升溫曲線上,冰點理論上為結晶剛剛熔解完成的溫度,但由于試樣熔化完后,基線受升溫速率和樣品量的影響,出現不同程度的滯后現象,因此把相變峰的峰值溫度確定為魚肉的冰點[28]。
水分形態分析[29]:假設魚糜樣品中水分的總焓變與純水相同,可凍結水(包括自由水和不易流動水)含量可通過樣品在0 ℃附近的焓變計算得到,按式(1)計算。
在40~80 ℃分析復合物的熱穩定性[30]。使用TA Universal Analysis軟件進行數據記錄并分析各處理組樣品DSC圖譜各峰的峰值溫度(Tp)和總變性焓(ΔH3)。
1.3.4 分子間作用力測定
根據Li Fengtang等[31]方法稍作修改。取2 g魚糜-褐藻寡糖復合物,分別與10 mL 0.05 mol/L?NaCl(SA)、0.6 mol/L NaCl(SB)、0.6 mol/L?NaCl+1.5 mol/L尿素(SC)、0.6 mol/L?NaCl+8 mol/L尿素(SD)、0.6 mol/L?NaCl+8 mol/L尿素+1.5 mol/L β-巰基乙醇(SE)混合,4 000 r/min均質3~5 min后,于4 ℃靜置1 h,8 000 r/min離心10 min,使用考馬斯亮藍法測定上清液中蛋白質含量。離子鍵、氫鍵、疏水相互作用力、二硫鍵含量分別按式(3)~(6)計算,每個樣品重復測定3 次。
離子鍵含量/(mg/mL)=ρSB-ρSA (3)
氫鍵含量/(mg/mL)=ρSC-ρSB (4)
疏水相互作用力/(mg/mL)=ρSD-ρSC (5)
二硫鍵含量/(mg/mL)=ρSE-ρSD (6)
式中:ρSA為溶解于SA溶液中的蛋白質質量濃度/(mg/mL);ρSB為溶解于SB溶液中的蛋白質質量濃度/(mg/mL);ρSC為溶解于SC溶液中的蛋白質質量濃度/(mg/mL);ρSD為溶解于SD溶液中的蛋白質質量濃度/(mg/mL);ρSE為溶解于SE溶液中的蛋白質質量濃度/(mg/mL)。
1.3.5 肌原纖維蛋白的提取
參考儀淑敏等[32]的方法。分別取上述魚糜-褐藻寡糖復合物5 g,加入25 mL 10 mmol/L、pH 7.2 Tris-HCl緩沖液,4 000 r/min均質2 min后,于4 ℃、5 000 r/min離心15 min,取沉淀加入20 mL 10 mmol/L Tris-HCl緩沖液(含0.6 mol/L NaCl,pH 7.2),于4 000 r/min均質30 s,4 ℃、4 500 r/min離心20 min后取上清,上清液即為肌原纖維蛋白溶液,用雙縮脲試劑測定上清液中肌原纖維蛋白含量。用10 mmol/L Tris-HCl緩沖液(含0.6 mol/L?NaCl,pH 7.2)將肌原纖維蛋白配制成質量濃度
1.0 mg/mL的溶液。
1.3.6 FTIR測定肌原纖維蛋白中蛋白質二級結構含量
根據Xu Yujuan等[33]方法稍作修改。將提取的各肌原纖維蛋白溶液進行冷凍干燥,取2 mg冷凍干燥后的肌原纖維蛋白粉末,與100 mg干燥的無水溴化鉀充分混合,在約18 N的壓力下加壓2 min,制成溴化鉀壓片。用FTIR儀對其進行測定和分析,分辨率4 cm-1,光譜掃描范圍400~4 000 cm-1。Peak Fit軟件用于酰胺Ⅰ譜帶的原始光譜曲線擬合。1 600~1 639 cm-1被認為是β-折疊結構波段,1 640~1 650 cm-1被認為是無規則卷曲結構波段,1 651~1 660 cm-1被認為是α-螺旋結構波段,1 661~1 700 cm-1被認為是β-轉角結構波段[34]。利用Peak Fit軟件,根據積分面積占比計算各二級結構的相對含量。
1.3.7 肌原纖維蛋白內源熒光光譜測定
取各組肌原纖維蛋白溶液,采用熒光分光光度計進行熒光光譜掃描,激發波長為280 nm,掃描范圍300~450 nm,測定熒光強度[35]。猝滅常數(Ksv)用Stern-Volmer方程計算[36],如式(7)所示。
式中:F0和F分別為空白組與添加褐藻寡糖組的熒光強度最大值;[Q]為褐藻寡糖質量濃度/(mg/mL)。
1.3.8 肌原纖維蛋白紫外吸收光譜測定
取各組肌原纖維蛋白溶液,以10 mmol/L Tris-HCl緩沖液(含0.6 mol/L NaCl,pH 7.2)作為對照。將樣品置于石英比色皿中,用紫外-可見分光光度計在波長230~350 nm獲得紫外吸收光譜。使用OriginPro 2017軟件從紫外吸收光譜獲得二階導數光譜[37]。
1.4 數據處理
采用SPSS軟件對實驗中所得數據進行顯著性分析(P<0.05),使用OriginPro 2017軟件制表、繪圖。
2 結果與分析
2.1 復合物DSC測定結果及分析
DSC技術可以定量測定樣品的物理轉變及化學反應,水分狀態的改變對食品的貯藏性、加工性和商品的利用價值都有極大影響。DSC技術可用來測定蛋白的結晶點、冰點等[38]。
由圖1可知,DSC曲線顯示了明顯的熔融溫度(Tp1),其中放熱峰為水分結晶峰。解凍鰱魚魚糜經褐藻寡糖修飾后其熔融溫度改變,表明加熱過程中晶態結構被破壞[39]。
由表1可知:除了1.0 g/100 mL AGO組,褐藻寡糖導致解凍鰱魚魚糜的水分結晶峰Tp1升高,且低質量濃度組更高;AMO組、EAO組水分結晶峰ΔH1升高,且高質量濃度組升高幅度較大,而AGO組ΔH1降低,且AGO高質量濃度組降低幅度較大,水分結晶峰的出現可能是由于本研究設置的程序中水分結晶峰值在-50~15 ℃升溫速率較大。
寡糖修飾后加快水分的相變,導致魚糜水分分布的改變,由于寡糖的親水性使復合物的親水性增強,從而提高了樣品的可凍結水含量[40]。高質量濃度褐藻寡糖對魚糜水分分布(可凍結水含量和非凍結水含量比例)的影響較大。1.0 g/100 mL AMO、1.0 g/100 mL AGO提高了冰點峰Tp2,而0.5 g/100 mL AMO、AGO、EAO則降低了冰點峰Tp2。1.0 g/100 mL AMO、AGO、EAO組ΔH2升高,可凍結水含量升高,而0.5 g/100 mL AMO、AGO組ΔH2降低,可凍結水含量降低,這是因為在空間允許的情況下,水分子有形成更多氫鍵的趨勢,以形成類似“籠形”的聚集體結構,水分子與寡糖之間也會形成部分氫鍵,以利于上述結構的形成,使更多水分子禁錮在寡糖-蛋白復合網絡結構中,可凍結水的流動性減弱,導致冰點降低,吸熱峰的峰值向低溫區移動[41]。高質量濃度褐藻寡糖組冰點峰值溫度升高,可能與EAO由M、G 2 種單體聚合而成有關,其較MM、GG聚合體較復雜。總的來說,AMO與AGO在抗凍性方面性能更優越,且低質量濃度(0.5 g/100 mL)褐藻寡糖更容易降低冰點、可凍結水含量更低,能更好地抵抗低溫,便于貯藏[42]。
DSC可以表征肌原纖維蛋白的結構變化,進而推斷熱穩定性[43]。圖1中40~80 ℃有3 個峰,其中峰Ⅰ代表肌球蛋白頭部引起的熱流變化,峰Ⅱ代表肌球蛋白尾部和肌漿蛋白引起的熱流變化,峰Ⅲ代表肌動蛋白引起的熱流變化,各峰的峰值溫度(表1中TpⅠ、TpⅡ、TpⅢ)為蛋白變性溫度,該溫度越高,蛋白質穩定性越高,峰Ⅰ、峰Ⅱ、峰Ⅲ峰面積的積分和為總變性焓(ΔH3)[44]。由表1可知,褐藻寡糖修飾后TpⅠ均升高,其中1.0 g/100 mL EAO、0.5 g/100 mL AMO、0.5 g/100 mL AGO組升高程度較大,表明褐藻寡糖修飾后,魚糜肌原纖維蛋白的熱穩定性增強[45]。寡糖能夠通過非共價相互作用和物理纏結改變魚糜肌原纖維蛋白的構象,使分子結構中的羥基與肌原纖維蛋白分子中的一些基團發生反應,同時也能增強肌原纖維蛋白的疏水相互作用[46],影響多肽鏈空間結構的重排[47],從而避免蛋白質變性。除1.0 g/100 mL AMO與0.5 g/100 mL EAO組Tp Ⅱ升高,其他組Tp Ⅱ均降低;除0.5 g/100 mL AMO組Tp Ⅲ降低,其他組Tp Ⅲ均升高,且經高質量濃度褐藻寡糖修飾后更高。AMO、AGO組ΔH3均升高,且高質量濃度AMO組升高程度更大。總的來說,AMO、AGO組熱穩定性都有所改善[47],且經高質量濃度褐藻寡糖修飾后樣品的熱穩定性較強。
2.2 復合物分子間作用力測定結果及分析
魚糜凝膠系統中的網絡結構主要是通過蛋白質分子間與分子內的相互作用來維持,如離子鍵、氫鍵、疏水相互作用和二硫鍵[48]。離子鍵和氫鍵是相對于疏水相互作用和二硫鍵較弱的鍵合力[49]。
由表2可知,鰱魚魚糜中存在大量離子鍵和氫鍵,與空白組相比較,1 g/100 mL AMO和1 g/100 mL EAO組離子鍵含量顯著降低(P<0.05),而1 g/100 mL AGO組離子鍵含量顯著升高(P<0.05),經低質量濃度組褐藻寡糖修飾后,離子鍵含量均升高,離子鍵可以維持肌原纖維蛋白三級結構,離子鍵被破壞意味著蛋白發生了聚集。與空白組相比較,除1 g/100 mL EAO組外,其他組氫鍵含量均降低,這種現象可能是由于寡糖在蛋白展開時與蛋白質競爭與水結合。EAO組疏水相互作用力下降,而0.5 g/100 mL AMO組疏水相互作用力顯著增加(P<0.05),且AGO組疏水相互作用力均上升,表明褐藻寡糖修飾對維持肌原纖維蛋白構象的穩定起著重要作用[50]。褐藻寡糖修飾后,高質量濃度褐藻寡糖組魚糜二硫鍵含量均上升,其中1 g/100 mL AMO組二硫鍵含量較高,巰基氧化,形成二硫鍵,導致蛋白質交聯增加,從而改善魚糜凝膠的性質[24]。
2.3 魚糜肌原纖維蛋白FTIR測定結果及分析
由圖2可知,魚糜肌原纖維蛋白在FTIR圖譜上有多個吸收波段,其中1 629 cm-1處的特征吸收峰為酰胺Ⅰ帶C=O的伸縮振動;1 555 cm-1處的特征吸收峰為酰胺Ⅱ帶C-N的伸縮與N-H的彎曲振動[51];1 180~953 cm-1有一系列重疊峰,為C-O和C-C伸縮振動和C-H的彎曲模式。褐藻寡糖修飾后,肌原纖維蛋白酰胺Ⅱ帶的特征吸收峰波數略有降低,可能是褐藻寡糖與蛋白發生靜電相互作用的原因[34];1 180~953 cm-1區域的透光率變大,可能是肌原纖維蛋白與寡糖之間形成了共價鍵,導致體系中羥基和碳氧鍵增多,使得該波段吸收增強[10]。加入褐藻寡糖后,3 192 cm-1處的吸收增強,這種變化可能是由N-H的減少引起的,這也證明了褐藻寡糖與肌原纖維蛋白之間可能通過共價鍵結合[34]。
酰胺Ⅰ帶是被公認的二級結構特征吸收峰,蛋白質的二級結構主要通過非共價作用力維持,當環境發生變化時,蛋白質分子將重新排列,以達到最低能量,維持相對穩定的狀態[47]。
由表3可知,魚糜肌原纖維蛋白二級結構中相對含量較高的主要是β-折疊,其次是無規則卷曲、β-轉角及α-螺旋。褐藻寡糖修飾后,α-螺旋相對含量均降低,高質量濃度褐藻寡糖修飾后α-螺旋相對含量低于低質量濃度組,α-螺旋相對含量降低表明褐藻寡糖通過與蛋白質分子相互作用抑制水溶性蛋白質和疏水殘留物的暴露[52]。褐藻寡糖修飾后,β-轉角相對含量均降低,其中高、低質量濃度EAO組β-轉角相對含量降低差距不大。除AGO組外,其他組β-折疊相對含量都有所升高,高、低質量濃度EAO組β-折疊相對含量增加較均衡。無規則卷曲結構相對含量均升高,尤其是AGO組,且AGO組與EAO低質量濃度組升高幅度較大,無規則卷曲相對含量的升高表明褐藻寡糖修飾使蛋白結構更加無序。可能是褐藻寡糖的修飾使β-轉角、α-螺旋向無規則卷曲轉變,肌原纖維蛋白具有更隨機的結構[53]。
2.4 魚糜肌原纖維蛋白內源熒光光譜測定結果及分析
蛋白質內源性色氨酸(Trp)熒光對于其周邊微環境的極性非常敏感,因此常用于監測蛋白質三級結構的變化[54]。由圖3可知,空白組熒光最大發射波長為331 nm,褐藻寡糖修飾后,魚糜肌原纖維蛋白的最大發射波長并沒有變化,但是熒光強度均有所增強,酸解樣品高質量濃度條件下熒光強度較高,酶解樣品低質量濃度條件下熒光強度較高。可能是由于褐藻寡糖修飾后,與蛋白之間發生聚合反應,蛋白反應性熒光強度升高[53],酸解AMO與AGO可能更容易與蛋白反應,且高質量濃度褐藻寡糖為蛋白提供了更多的結合位點[53]。一般來說,生色團與猝滅劑相互作用會降低熒光量子產率。本研究結果與其相悖,可能歸因于褐藻寡糖與肌原纖維蛋白發生靜電相互作用,增加了疏水性Trp發色團微環境的極性,使Trp殘基更多地埋藏在肌原纖維蛋白分子中,從而導致與猝滅劑的相互作用程度較低[55]。
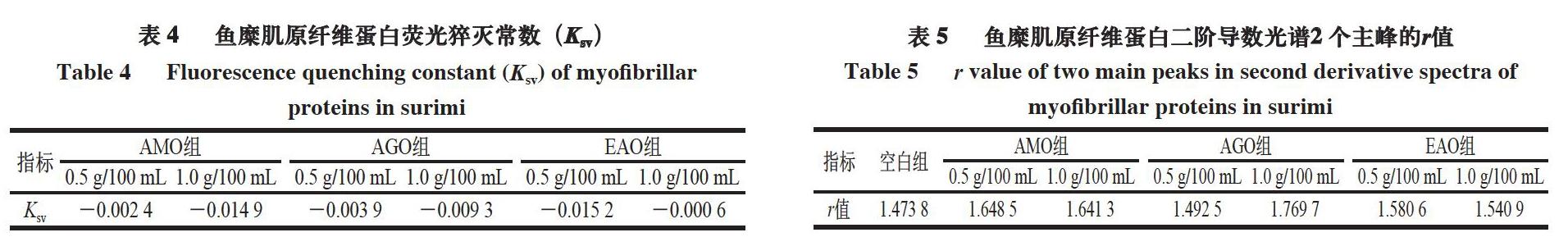
由表4可知,所有褐藻寡糖修飾后樣品Ksv皆為負值,也說明褐藻寡糖修飾后使肌原纖維蛋白溶液的極性改變。
2.5 魚糜肌原纖維蛋白紫外吸收光譜測定結果及分析
色氨酸(Trp)、酪氨酸(Tyr)和苯丙氨酸(Phe)的R基團含有苯環共軛雙鍵,分別在280、275、257 nm波長處有吸收峰。肌原纖維蛋白的最大吸收波長在275 nm附近,可能是由于Tyr未包埋在蛋白質內部。
由圖4可知,在275、280 nm波長處,各樣品的最大吸收峰并未發生移動,表明肌原纖維蛋白中Tyr微環境變化不明顯。高質量濃度褐藻寡糖組肌原纖維蛋白紫外吸收峰強度增強,且3 種褐藻寡糖中AMO組增加幅度較大,說明褐藻寡糖與肌原纖維蛋白之間發生分子內或分子間的相互作用[56]。
紫外二階光譜可以進一步研究肌原纖維蛋白構象的細微差異,在288 nm(Trp、Tyr共同作用)和296 nm(Trp作用)波長處有2 個正吸收峰,在284、291 nm波長處有2 個負吸收峰[57],因此需采用紫外二階導數生成r值(r=a/b,a、b分別為第1次和第2次正吸收峰與負吸收峰的差值),來分析Tyr殘基微環境。且褐藻寡糖修飾后,二階導數光譜峰的位置幾乎未發生變化。
Tyr在極性溶劑中的暴露增加會導致r值增加。由表5可知,與空白組相比,褐藻寡糖修飾后r值增大,1.0 g/100 mL AGO組增加幅度較大,3 種褐藻寡糖修飾后r值均升高,可能是由于褐藻寡糖通過大量羥基與Tyr形成氫鍵,從而使Tyr暴露于分子表面[57],也表明1.0 g/100 mL AGO修飾后的蛋白質聚集程度較高[58]。
3 結 論
褐藻寡糖(0.5、1.0 g/100 mL)的修飾對鰱魚魚糜的穩定性有一定的改善,1 g/100 mL組的熱穩定性更強,0.5 g/100 mL組魚糜冷凍穩定性顯示出較好的優勢。褐藻寡糖的修飾對蛋白質離子鍵具有保護作用,且1 g/100 mL AGO組最優;褐藻寡糖通過靜電相互作用與蛋白結合,且改變了肌原纖維蛋白的二級結構,使β-轉角、α-螺旋向無規則卷曲轉變,蛋白結構更加無序;褐藻寡糖修飾后,改變了蛋白周圍的極性環境,色氨酸被埋藏于分子內,而酪氨酸更容易暴露,使蛋白結構更加聚集。
本研究討論了不同來源的褐藻寡糖作為抗凍劑或熱穩定劑在冷凍魚糜行業的應用,研究結果有宏觀尺度的對鰱魚魚糜品質的影響,也有分子尺度的對肌原纖維蛋白結構的修飾,仍需對微觀結構進行表征,以進一步明確褐藻寡糖對蛋白質的修飾機制。本研究為系統理解復雜食品組分結構變化提供了依據,為開發新產品和應用提供了參考。
參考文獻:
[1] 農業農村部漁業漁政管理局. 2020中國漁業統計年鑒[EB/OL]. (2020-06) [2021-06-15]. https://data.cnki.net/trade/Yearbook/Single/N2021020168?z=Z009.
[2] Abdollahi M, Rezaei M, Jafarpour A, et al. Sequential extraction of gel-forming proteins, collagen and collagen hydrolysate from gutted silver carp (Hypophthalmichthys molitrix), a biorefinery approach[J]. Food Chemistry, 2018, 242: 568-578. DOI:10.1016/j.foodchem.2017.09.045.
[3] 于傳, 王嵬, 喬翠平, 等. 金線魚-白鰱魚混合魚糜凝膠蛋白質構象和作用力的研究[J]. 食品安全質量檢測學報, 2020, 11(22): 60-69.
[4] 榮婧, 仇超穎, 胡曉, 等. 鳶烏賊肌原纖維蛋白糖基化產物功能特性研究[J]. 南方水產科學, 2018, 14(1): 70-78. DOI:10.3969/j.issn.2095-0780.2018.01.009.
[5] Sato R, Katayama S, Sawabe T, et al. Stability and emulsion-forming ability of water-soluble fish myofibrillar protein prepared by conjugation with alginate oligosaccharide[J]. Journal of Agricultural and Food Chemistry, 2003, 51(15): 4376-4381. DOI:10.1021/jf021023v.
[6] 黃菊, 丁晨, 謝超, 等. 海藻膠低聚寡糖對秘魯魷魚(Peru squid)魚糜品質特性的影響研究[J]. 海洋與湖沼, 2015, 46(3): 659-664. DOI:10.11693/hyhz20150300073.
[7] Uresti R M, Velazquez G, Vazquez M, et al. Effect of sugars and polyols on the functional and mechanical properties of pressure-treated arrowtooth flounder (Atheresthes stomias) proteins[J]. Food Hydrocolloids, 2005, 19(6): 964-973. DOI:10.1016/j.foodhyd.2004.12.006.
[8] LI Shibo, HE Zhifei, LI Mingqi, et al. A study of the physicochemical properties of rabbit glycated myofibrillary protein with high solubility in low ionic strength medium[J]. International Journal of Biological Macromolecules, 2020, 147: 241-249. DOI:10.1016/j.ijbiomac.2020.01.069.
[9] Fujiwara K, Oosawa T, Saeki H. Improved thermal stability and emulsifying properties of carp myofibrillar proteins by conjugation with dextran[J]. Journal of Agricultural and Food Chemistry, 1998, 46(4): 1257-1261. DOI:10.1021/jf9708148.
[10] 楊宇鴻, 董士遠, 靳衛亞, 等. 草魚肌原纖維蛋白-葡萄糖糖基化產物的理化特性及乳化特性研究[J]. 食品工業科技, 2018, 39(18): 77-82; 142. DOI:10.13386/j.issn1002-0306.2018.18.015.
[11] Mahmood S J, Siddique A. Ionic studies of sodium alginate isolated from Sargassum terrarium (brown algea) karachi coast with 2,1-electrolyte[J]. Journal of Saudi Chemical Society, 2010, 14(1): 117-123. DOI:10.1016/j.jscs.2009.12.018.
[12] Bemiller J N. Carbohydrate chemistry for food scientists[M]. 3th. Woodhead Publishing and AACC International Press, 2019: 293-301. DOI:10.1016/B978-0-12-812069-9.00014-5.
[13] Osojnik G, Natasha P U. Biopolymer nanostructures for food encapsulation purposes[M]. Salt Lake City: Academic Press, 2019: 1; 335-338. DOI:10.1016/B978-0-12-815663-6.00013-6.
[14] 周緒霞, 徐鋆, 丁玉庭. 酶解制備褐藻膠寡糖及其產物的抗氧化活性分析[J]. 食品與發酵工業, 2014, 8(2): 116-120. DOI:10.13995/j.cnki.11-1802/ts.2014.02.008.
[15] 王雪妹, 王晶, 張全斌. 褐藻多糖硫酸酯對脂多糖誘導大鼠腎小球系膜細胞NO產生量的影響[J]. 海洋科學, 2014, 38(10): 1-5. DOI:10.11759/hykx20131218002.
[16] Luan L, Nagasawa N, Ha V, et al. Enhancement of plant growth stimulation activity of irradiated alginate by fractionation[J]. Radiation Physics and Chemistry, 2009, 78(9): 796-799. DOI:10.1016/j.radphyschem.2009.05.001.
[17] 董曉莉, 耿美玉, 管華詩, 等. 褐藻酸性寡糖對帕金森病大鼠紋狀體、杏仁核多巴胺釋放的影響[J]. 中國海洋藥物, 2003, 22(5): 9-12.
[18] 吳燕燕, 張婉, 李來好, 等. 海藻中抗氧化、保濕功能活性物質的研究進展[J]. 海洋科學, 2015, 39(9): 138-142. DOI:10.11759hykx20140729002.
[19] Iwamoto M, Kurachi M, Nakashima T, et al. Structure-activity relationship of alginate oligosaccharides in the induction of cytokine production from RAW264.7 cells[J]. FEBS Letters, 2005, 579(20): 4423-4429. DOI:10.1016/j.febslet.2005.07.007.
[20] Iwamoto Y, Xu X, Tamura T, et al. Enzymatically depolymerized alginate oligomers that cause cytotoxic cytokine production in human mononuclear cells[J]. Bioscience, Biotechnology, and Biochemistry, 2003, 67(2): 258-263. DOI:10.1271/bbb.67.258.
[21] YANG J H, BANG M A, JANG C H, et al. Alginate oligosaccharide enhances LDL uptake via regulation of LDLR and PCSK 9 expression[J]. Journal of Nutritional Biochemistry, 2015, 26(11): 1393-1400. DOI:10.1016/j.jnutbio.2015.07.009.
[22] 胡博旸, 聶瑩, 孫路, 等. 酸降解法制備褐藻寡糖抗氧化性的研究[J]. 食品工業科技, 2016, 37(10): 136-140. DOI:10.13386/j.issn1002-0306.2016.10.018.
[23] LI Shangyong, WANG Zhipeng, WANG Linna, et al. Combined enzymatic hydrolysis and selective fermentation for green production of alginate oligosaccharides from Laminaria japonica[J]. Bioresource Technology, 2019, 281: 84-89. DOI:10.1016/j.biortech.2019.02.056.
[24] MI Hongbo, LI Yi, WANG Cong, et al. The interaction of starch-gums and their effect on gel properties and protein conformation of silver carp surimi[J]. Food Hydrocolloids, 2020, 112: 106290. DOI:10.1016/j.foodhyd.2020.106290.
[25] 趙丹, 汪秋寬, 張慧怡, 等. 海帶粗提物及褐藻酸鈉寡糖的保濕與抗氧化作用研究[J]. 水產科學, 2012, 31(6): 358-362. DOI:10.3969/j.issn.1003-1111.2012.06.010.
[26] 楊姣, 安玥琦, 陳雨欣, 等. 魚糜制品加熱過程中過熟味的特征風味成分解析[J]. 現代食品科技, 2020, 36(8): 265-280. DOI:10.13982/j.mfst.1673-9078.2020.8.0169.
[27] 黃海. DSC在食品中的運用[J]. 食品與機械, 2002(2): 6-9. DOI:10.3969/j.issn.1003-5788.2002.02.002.
[28] 楊協力, 朱文學, 劉麗莉. DSC在食品加工與貯藏中的應用及發展趨勢[J]. 保鮮與加工, 2008, 8(4): 1-4. DOI:10.3969/j.issn.1009-6221.2008.04.002.
[29] 胡佳慧, 熊光權, 喬宇, 等. 微凍貯藏過程中能量代謝酶活性和蛋白質氧化降解對鱸魚質構特性的影響[J]. 食品科學, 2018, 39(23): 236-243. DOI:10.7506/spkx1002-6630-201823034.
[30] CHENG Jingrong, ZHU Mingjun, LIU Xueming. Insight into the conformational and functional properties of myofibrillar protein modified by mulberry polyphenols[J]. Food Chemistry, 2020, 308: 125592. DOI:10.1016/j.foodchem.2019.125592.
[31] LI Fengtang, ZHAO Jianxin, HUANG Jie, et al. Improvement of the quality of surimi products with over-drying potato starches[J]. Journal of Food Quality, 2017, 2017: 1-5. DOI:10.1155/2017/1417856.
[32] 儀淑敏, 李睿智, 陳楊, 等. 白鰱魚肌原纖維蛋白雙向電泳分析體系的建立[J]. 食品科學, 2017, 38(1): 41-46. DOI:10.7506/spkx1002-6630-201701007.
[33] XU Yujuan, ZHAO Xue, BIAN Guangliang, et al. Structural and solubility properties of pale, soft and exudative (PSE)-like chicken breast myofibrillar protein: effect of glycosylation[J]. Food Science and Technology, 2018, 95: 209-215. DOI:10.1016/j.lwt.2018.04.051.
[34] Xu Yujuan, Dong Ming, Tang Changbo, et al. Glycation-induced structural modification of myofibrillar protein and its relation to emulsifying properties[J]. Food Science and Technology, 2020, 117: 108664. DOI:10.1016/j.lwt.2019.108664.
[35] Estevez M, Kylli P, Puolanne E, et al. Fluorescence spectroscopy as a novel approach for the assessment of myofibrillar protein oxidation in oil-in-water emulsions[J]. Meat Science, 2008, 80(4): 1290-1296. DOI:10.1016/j.meatsci.2008.06.004.
[36] JIANG Jiang, ZHANG Zhipeng, ZHAO Jing, et al. The effect of non-covalent interaction of chlorogenic acid with whey protein and casein on physicochemical and radical-scavenging activity of in vitro protein digests[J]. Food Chemistry, 2018, 268: 334-341. DOI:10.1016/j.foodchem.2018.06.015.
[37] WANG Guan, LIU Manman, CAO Liwei, et al. Effects of different NaCl concentrations on self-assembly of silver carp myosin[J]. Food Bioscience, 2018, 24: 1-8. DOI:10.1016/j.fbio.2018.05.002.
[38] 艾文婷, 張敏, 黃汝國, 等. 熱分析技術在食品熱物性研究中的應用[J]. 食品工業科技, 2016, 37(19): 377-380. DOI:10.13386/j.issn1002-0306.2016.19.065.
[39] 于源, 張敏, 邵弘. 差示掃描量熱法在大豆蛋白產品品質檢測中的應用[J]. 大豆科技, 2008(1): 26-28; 36. DOI:10.3969/j.issn.1674-3547.2008.01.013.
[40] 邵穎, 王小紅, 吳文錦, 等. 食鹽添加量對預制鱸魚冷藏保鮮及熱加工特性的影響[J]. 農業工程學報, 2016, 32(12): 280-286. DOI:10.11975/j.issn.1002-6819.2016.12.040.
[41] 周緒霞, 姜珊, 顧賽麒, 等. 油茶籽油對魚糜凝膠特性及凝膠結構的影響[J]. 食品科學, 2017, 38(9): 27-33. DOI:10.7506/spkx1002-6630-201709005.
[42] 楊嘉琪, 宋春麗. 糖基化及酶解對大豆蛋白功能性質的影響[J]. 食品與發酵工業, 2020, 46(1): 125-129. DOI:10.13995/j.cnki.11-1802/ts.022122.
[43] LIANG Feng, LIN Lin, HE Tianhai, et al. Effect of transglutaminase on gel properties of surimi and precocious Chinese mitten crab (Eriocheir sinensis) meat[J]. Food Hydrocolloids, 2020, 98: 105261. DOI:10.1016/j.foodhyd.2019.105261.
[44] 李俠, 孫圳, 楊方威, 等. 適宜凍結溫度保持牛肉蛋白穩定性抑制水分態變[J]. 農業工程學報, 2015, 31(23): 238-245. DOI:10.11975/j.issn.1002-6819.2015.23.032.
[45] 陳雨桐, 梁宏閃, 李斌. 單寧酸對小麥醇溶蛋白結構及功能性質的影響[J]. 食品工業科技, 2020, 41(6): 39-46; 52. DOI:10.13386/j. issn1002-0306.2020.06.007.
[46] 劉宏影, 馬瑩瑩, 李秀霞, 等. 解凍方式對金線魚肌原纖維蛋白熱穩定性和組織結構的影響[J]. 華中農業大學學報, 2019, 38(6): 25-32. DOI:10.13300/j.cnki.hnlkxb.2019.06.004.
[47] 郭金英, 賀億杰, 韓四海, 等. 魔芋葡甘聚糖對冷凍小麥面團面筋蛋白結構和功能特性的影響[J]. 食品科學, 2019, 40(24): 33-39. DOI:10.7506/spkx1002-6630-20181225-294.
[48] NUNEZ-Flores R, Cando D, Borderías A J, et al. Importance of salt and temperature in myosin polymerization during surimi gelation[J]. Food Chemistry, 2018, 239(15): 1226-1234. DOI:10.1016/j.foodchem.2017.07.028.
[49] ZHANG Jinchuang, LIU Li, JIANG Yuanyong. High-moisture extrusion of peanut protein-/carrageenan/sodium alginate/ wheat starch mixtures: Effect of different exogenous polysaccharides on the process forming a fibrous structure[J]. Food Hydrocolloids, 2020, 99: 105311. DOI:10.1016/j.foodhyd.2019.105311.
[50] 汪之和, 朱孔輝, 施文正, 等. 鳊魚魚糜蛋白質結構與凝膠特性的關系[J]. 水產學報, 2010, 34(5): 814-819. DOI:10.3724/SP.J.1231.2010.06715.
[51] Muyonga J H, Cole G B, Duodu K G. Fourier transform infrared (FTIR) spectroscopic study of acid soluble collagen and gelatin from skins and bones of young and adult Nile perch (Lates niloticus)[J]. Food Chemistry, 2004, 86(3): 325-332. DOI:10.1016/j.foodchem.2003.09.038.
[52] Walayat N, XIONG Zhouyi, XIONG Hanguo, et al. The effectiveness of egg white protein and β-cyclodextrin during frozen storage: functional, rheological and structural changes in the myofibrillar proteins of Culter alburnus[J]. Food Hydrocolloids, 2020, 105: 105842. DOI:10.1016/j.foodhyd.2020.105842.
[53] XU Zhenzhen, HUANG Guoqing, XU Tongcheng, et al. Comparative study on the Maillard reaction of chitosan oligosaccharide and glucose with soybean protein isolate[J]. International Journal of Biological Macromolecules, 2019, 131: 601-607. DOI:10.1016/j.ijbiomac.2019.03.101.
[54] 汪蘭, 吳文錦, 喬宇, 等. 凍藏條件下魔芋葡甘聚糖降解產物對肌原纖維蛋白結構的影響[J]. 食品科學, 2015, 36(22): 244-249. DOI:10.7506/spkx1002-6630-201522046.
[55] 周向軍, 楊雪純, 劉海玉, 等. 木糖醇和甘露醇對花生蛋白結構和功能特性的影響[J]. 食品與機械, 2018, 34(7): 17-22. DOI:10.13652/j.issn.1003-5788.2018.07.004.
[56] XU Yujuan, ZHAO Yuqi, WEI Zhixin, et al. Modification of myofibrillar protein via glycation: physicochemical characterization, rheological behavior and solubility property[J]. Food Hydrocolloids, 2020, 105: 105852. DOI:10.1016/j.foodhyd.2020.105852.
[57] 孫亞天, 程志先, 李果, 等. 大豆分離蛋白與殼聚糖相互作用的光譜法研究[J]. 核農學報, 2018, 32(4): 723-731. DOI:10.11869/j.issn.100-8551.2018.04.0723.
[58] PENG Zeyu, ZHU Mingming, ZHANG Juan, et al. Physicochemical and structural changes in myofibrillar proteins from porcine Longissimus dorsi subjected to microwave combined with air convection thawing treatment[J]. Food Chemistry, 2021, 343: 128412. DOI:10.1016/j.foodchem.2020.128412.
