基于16S rDNA擴增子測序分析靈芝連作覆土細菌群落的變化
2021-09-05 03:35:08黃海辰劉國輝謝福泉傅俊生吳小平
中國農學通報 2021年24期
袁 源,李 琳,黃海辰,劉國輝,謝福泉,傅俊生,吳小平
(1福建農林大學生命科學學院,福州350002;2福建農林大學菌物研究中心,福州350002;3仙芝科技(福建)股份有限公司,福建南平353400;4福建省食用菌技術推廣總站,福州350003)
0 引言
靈芝是一種天然的藥用真菌,目前已報道400多個分類單元的靈芝科,國內普遍栽培的靈芝通常是指‘赤芝’,崔寶凱等[1]認為‘赤芝’的拉丁學名為Ganoderma lingzhi Sheng H.Wu,Y.Cao&Y.C.Dai,而國內外誤用Ganoderma lucidum(Curtis)P.Karst.作為‘赤芝’的拉丁學名已經100余年。其富含多糖,三萜和靈芝酸等活性成分,具有顯著的抗腫瘤、抗氧化和降血脂等活性,具有良好的藥用價值和保健功能,深受眾多消費者的喜愛[2]。為滿足消費者對靈芝日益增長的市場需求,人工栽培目前已成為靈芝供應的主要來源。到目前為止,中國人工栽培靈芝的歷史已超過了60年,靈芝段木覆土栽培是一種重要的靈芝的栽培模式,段木為靈芝的生長提供豐富的營養成分,覆土能穩定料中的濕度,維持良好的通氣性,該模式生產上具有顯著的低污染,易控制,產量高,并且子實體營養豐富和藥效成分全面等特點,但連作2~3年后靈芝覆土依然存在常見的連作障礙問題,往往容易引發靈芝子實體畸形,病害嚴重以及產量急劇下降等現象,將會導致大面積的靈芝基地不得不停用,制約了靈芝產業的良性發展,也不利于土地資源的循環利用[3-5]。
研究發現,連作障礙的主要原因包括微生物群落的改變、化感作用和自毒作用等[6-7]。同時,多項報道顯示靈芝連作障礙與覆土中微生物群落的變化具有較強的相關性,研究微生物群落變化有助于闡明靈芝連作障礙的機制[4-5]。微生物是土壤生態系統中關鍵的構成要素,也是一種重要的土壤質量評價指標[5]。一方面,在靈芝段木覆土栽培過程中,段木可以更新,大棚內的溫度和濕度可進行人工調節,但是靈芝的自毒作用、化感作用以及土壤理化性質的改變等均會導致覆土內微生物群落發生變化,這種菌群的改變反映了土壤質量的變化;另一方面,當土壤的質量發生變化,靈芝覆土中微生物的群落的豐度和數量又會相應的改變,從而對覆土中微生物群落進行研究就顯得尤為重要[8]。當前對靈芝連作障礙的相關研究報道主要集中在覆土中的真菌群落,如Kang等[9]基于形態學和系統發育樹分析,發現木霉屬與靈芝連作過程中產生的黃腐病有關;馬紅梅等[10]通過平板對峙培養法,發現子囊菌門的木霉屬、青霉屬和鏈孢霉屬等真菌是抑制靈芝生長主要的病原菌,而對靈芝連作覆土中細菌群落系統變化的研究卻鮮有報道。因此,文章對靈芝覆土中的細菌群落進行深入的探究。
近年來,基因測序在有關土壤微生物方面的研究中發揮著十分重要的作用,能對土壤中不同類群的微生物進行全面系統的分析[11]。16S rDNA擴增子測序技術是研究土壤微生物群落多樣性變化的基本手段之一,常用于反映樣本中細菌類群的種間差異[12]。該研究基于16S rDNA擴增子技術,對不同栽培年限靈芝覆土中細菌群落進行系統的分析,以期為靈芝連作障礙機制的研究奠定基礎。
1 材料和方法
1.1 材料與試劑
試驗菌株為‘赤芝119’(‘滬農靈芝1號’),由上海市農業科學院食用菌研究所進行新品種認定,審定編號為滬農品認食用菌(2009)第003號。土壤DNA提取試劑盒(OMEGA公司)。
1.2 儀器與設備
22R冷凍離心機(Microfuge);DK-8D型三溫三控恒溫水浴鍋(上海博迅實業有限公司);WH-861型旋渦震蕩儀(華利達公司);PCR儀(杭州朗基科學儀器有限公司);EPS300型電泳儀(上海天能科技有限公司)等。
1.3 試驗基地概況
試驗地位于仙芝科技(福建)股份有限公司有機靈芝博覽園(福建省南平市浦城縣,28°01’N,118°32’E),地處福建省最北端,屬于亞熱帶季風氣候,四季分明,全年平均氣溫17.4℃[13]。基地一共設有41個標準大棚,試驗土壤均源自基地同一區域內品質優良的沙壤土,pH呈弱酸型(5~6),適于靈芝生長出菇,現已成功培植赤芝等14個優良品種。
1.4 試驗方法
采用靈芝段木覆土栽培模式對‘赤芝119’進行栽培,將同一批生長狀態一致的‘赤芝119’脫袋后,2018年3月中旬待其菌絲長滿段木,及時移入大棚開始栽培,分別埋入通氣較好的栽培1年(30號大棚)、連作1年(38號大棚)、2年(6號大棚)和3年(17號大棚)的覆土中,使覆土能覆蓋住段木頂部2~3 cm,進一步進行出芝管理。在此過程中,4個大棚內的光照、溫度、濕度、通風以及操作過程等均控制一致,并且每年采集一次靈芝孢子粉和采收一次靈芝。
根據浦城縣有機靈芝博覽園的靈芝生產安排,8月至9月為靈芝采收階段,9月底靈芝采收結束。為了更好的反映靈芝連作覆土中細菌群落的變化,靈芝栽培1年、連作1年、2年和3年的覆土以及基地內未栽培過靈芝的鄰近沙壤土,于2018年9月25日靈芝采收期間取自有機靈芝博覽園。試驗設計如下:在4個實驗組中,每個分組面積為150 m2的大棚分別均勻栽培100棒靈芝。以鄰近沙壤土為對照組,將栽培1年、連作1年、2年、3年的靈芝覆土采用S形取樣法,按照半徑20~25 cm、深度0~5 cm的范圍使用鐵鏟進行取樣,每個分組取5個點的混合土樣,分別記為GL0、GL1、GL2、GL3和GL4組。每個分組混合土樣大約200 g,采集后分別裝入自封袋并且編號,置于冰盒內帶回實驗室,于-20℃冰箱保存。
1.5 土壤總DNA的提取
準確稱量0.2 g土壤樣品,加入含500 mg玻璃珠的15 mL離心管中,參照OMEGA土壤DNA提取試劑盒(D5625-02)說明書,對靈芝土樣的DNA進行提取,通過1%瓊脂糖凝膠電泳和核酸微量測定儀對所獲得的DNA進行質檢和定量,每個分組設定3個生物學重復,一共15個土壤樣本(FS_1、FS_2、FS_3、ONE_S_1、ONE_S_2、ONE_S_3、TWO_S_1、TWO_S_2、TWO_S_3、THREE_S_1、THREE_S_2、THREE_S_3、FOUR_S_1、FOUR_S_2和FOUR_S_3)。
1.6 PCR擴增及測序
參考趙力的方法,采用特異性引物338F(5'-ACTCCTACGGGAGGCAGCAG-3')和 806R(5'-GGACTACHVGGGTWTCTAAT-3')對16S rDNA可變保守區(V3+V4)進行PCR擴增[14],PCR反應體系和反應條件如實驗室已有報道所示[15],對PCR擴增產物進行檢測和回收。在Qbit熒光定量系統上采用PicoGreen雙鏈DNA熒光定量測定試劑盒對純化后的PCR產物文庫進行定量。將合格的文庫(濃度>2 nmol/L)梯度稀釋后,按一定要求進行比例混合和變性處理,變性為單鏈后可進行上機測序;于MiSeq測序儀上使用MiSeq Reagent Kit試劑盒進行2×300 bp的雙端測序。
1.7 生物信息學分析流程
將靈芝樣品送至杭州聯川生物技術股份有限公司(LC-Bio)在Illumina MiSeq平臺上進行測序。依據雙端序列之間的重疊關系,采用FLASH(v1.2.8)軟件將序列拼接成長的tag,去除建庫引入的barcode和引物序列,采用Vsearch(v2.3.4)過濾嵌合體。預處理之后的clean data使用Vsearch將序列相似度大于97%的有效序列定為一個OUT(operational taxonomic unit),選出位于幾何中心的代表序列。多樣性通過QIIME(v1.8.0)分析。通過blast進行序列比對,將OTU代表序列與RDP(ribosomal database project)以及UNITE數據庫對每個代表性序列進行物種注釋。其余圖片均是通用R包(V3.2.5)實現。并將序列數據和基因注釋信息上傳到NCBI網站數據庫。
1.8 數據統計學分析
采用SPSS17.0統計軟件對測序數據進行分析處理。組間比較采用t檢驗,以P<0.05表示差異顯著,P<0.01表示差異極顯著。
2 結果與分析
2.1 OTU聚類分析
對原始數據進行質控、雙端拼接、去除低質量以及嵌合體過濾后,對高質量數據進行統計,一共得到有效序列477556條。其中,400~500 bp序列約占總序列的97.05%,小于200 bp,200~300 bp和300~40 bp的序列分別占2.20%,0.64%和0.11%。將上述15個土壤樣本的477556條有效序列按照堿基相似度大于97%聚類成為不同的OTUs,得到每個cluster的代表序列則為一個OTU[16]。如圖1所示,Venn圖可直觀表示5個分組之間共有和特有的OTU序列和豐度,不同顏色的橢圓表示不同的分組[15]。5個分組經聚類后一共獲得9129個OTU,其中,GL0、GL1、GL2、GL3和GL4組OTU的數目分別為4689、5129、6506、5186和4281個。連作1年的覆土中OTU數目達到最大值,即6506個;連作3年的靈芝覆土中OTU數目最少,為4281個。表明隨著靈芝連作年限的增加,其覆土中的細菌類群數量呈先增后減的趨勢。同時,5個分組共有的OTU數目為851個,特有的OTU數目從GL0至GL4組依次為214、101、121、510和311個,靈芝連作2年和3年覆土細菌中特有的OTU數目大幅增加,表明特有細菌類群增加。
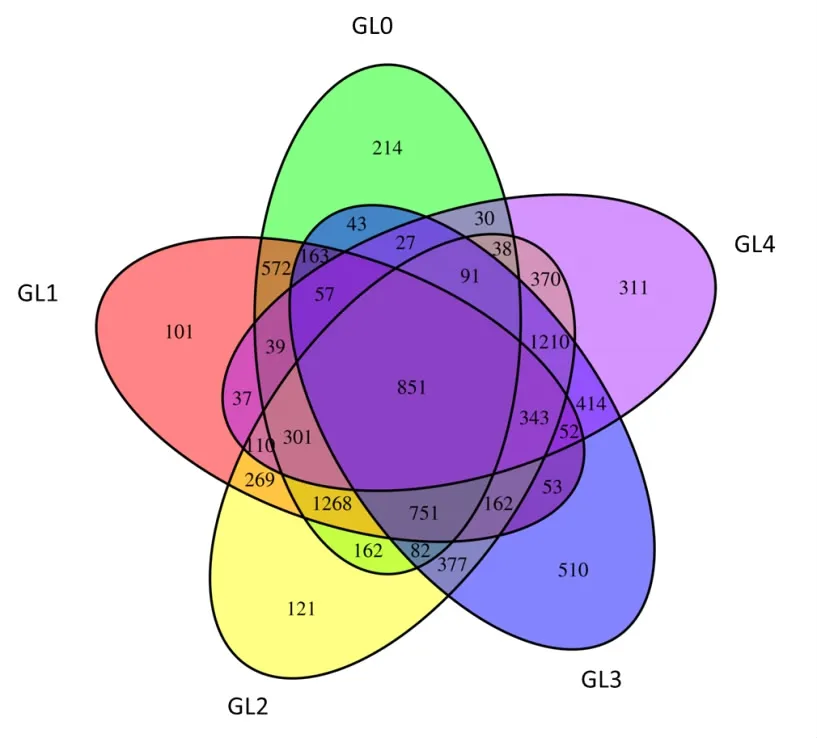
圖1 覆土細菌群落OTU聚類維恩圖
2.2 覆土細菌物種的Alpha多樣性
Alpha多樣性分析常用于衡量群落中物種豐富度、多樣性以及測序深度,通常包括chao1、observed species、Shannon、Simpson和 goods_coverage5個指數。chao1指數和observed species指數反映樣品中物種的豐富度;Shannon和Simpson指數分別反映樣品中物種的多樣性以及不同種類的物種豐度的均勻性;goods_coverage指數是指微生物覆蓋率,用于反映測序的深度,其數值越接近1,則樣本中沒有被測出的新物種概率越低。如圖2A和2B所示,通過對5個分組的chao1指數和observed species指數進行分析,發現靈芝連作1年覆土中細菌物種數目極顯著增加,隨后開始下降。如圖2C和2D的Shannon和Simpson指數所示,靈芝連作前2年覆土中細菌群落多樣性水平極顯著增加,隨后開始減少,且物種豐度的均勻性較好。如圖2E和2F所示,5個分組中goods_coverage指數均接近1,說明測序深度已足夠,基本覆蓋土壤樣品中的所有物種。由此表明,靈芝覆土中細菌的豐富度和多樣性隨著連作年限的推移均呈先增后減的趨勢,并且測序深度足夠。
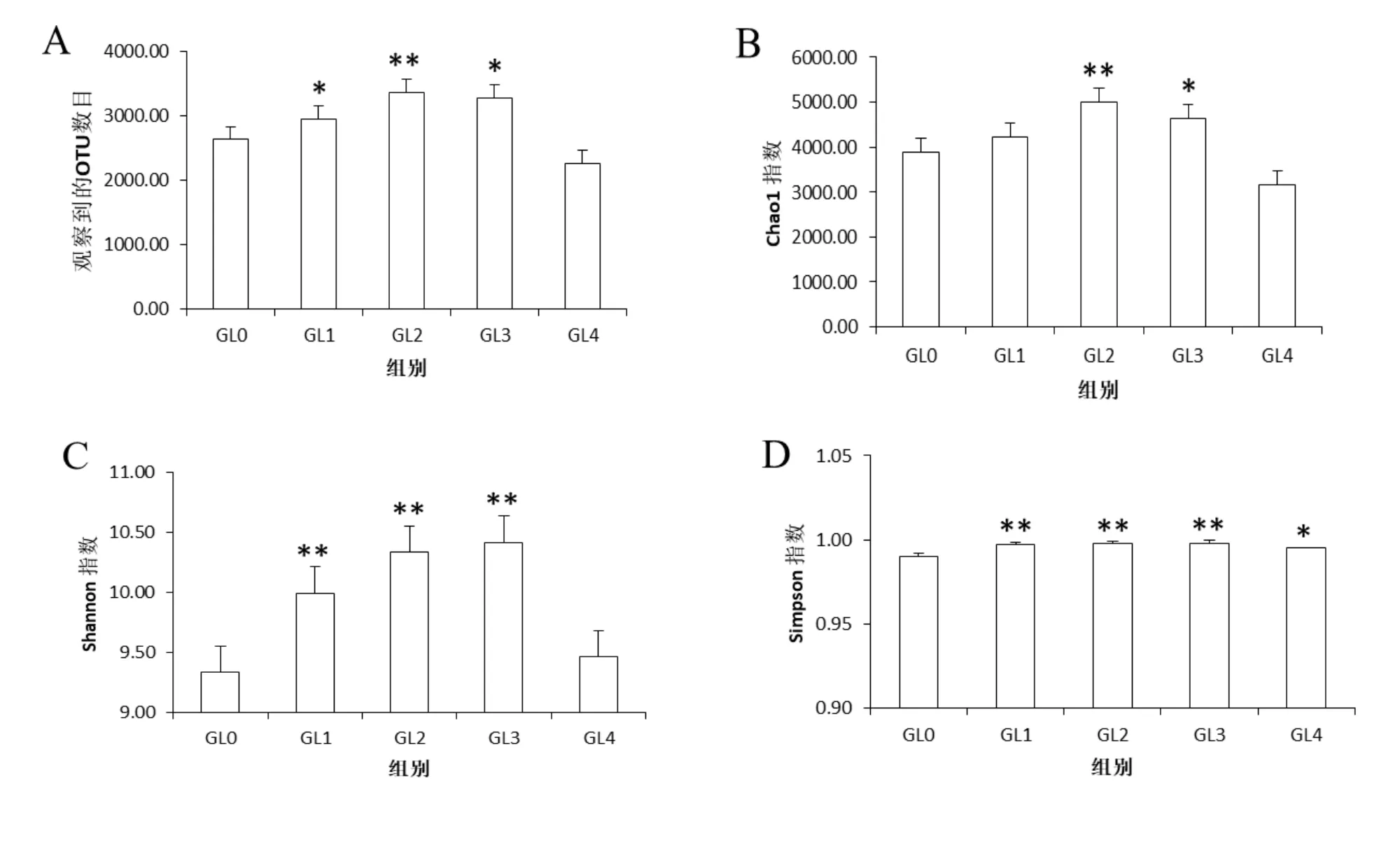
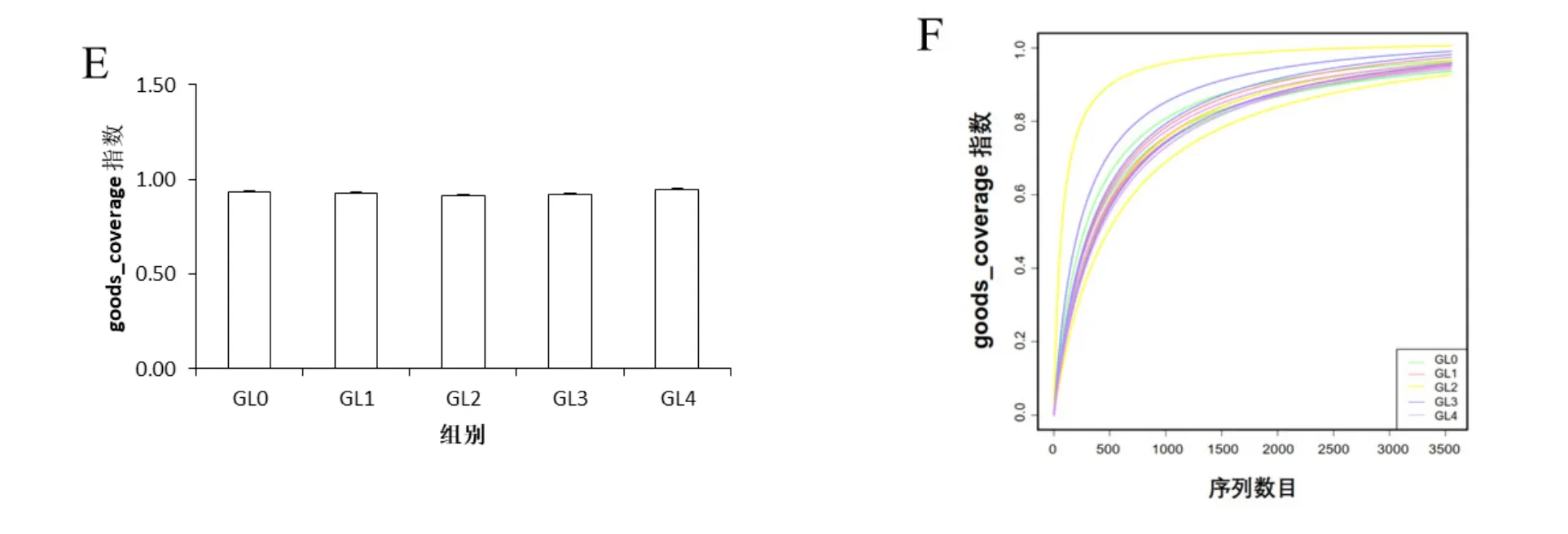
圖2 覆土細菌群落的Alpha多樣性指數的分析
2.3 覆土細菌物種的Beta多樣性
Beta多樣性在時空尺度上衡量物種組成的變化,以反映不同環境中群落間的物種組成差異[17-18]。如圖3A所示,采用非加權平均距離法(UPGMA)對15個樣本進行聚類,聚類樹展現了樣品間的相似度。其中,5個分組一共聚為兩個大分枝,GL0和GL1組靈芝覆土距離最近,表明鄰近沙壤土與栽培1年靈芝覆土之間細菌群落組成結構相似度最高,而GL2,GL3和GL4組聚為一個分枝并與GL0組的距離逐漸加大,表明連作逐漸降低了靈芝覆土中細菌物種組成的相似度。主坐標分析(PCoA)是一種常用的數據降維方法,可反映各個樣本群落組成的相似性或差異性[19]。樣本間距離越小,細菌群落組成的相似度越高;反之,細菌群落組成的差異性越大。如圖3B所示,與UPGMA聚類分析結果類似,隨著靈芝連作年限的增加,覆土中細菌組成的差異逐漸加大。此外,采用箱線圖對組內和組間Unifrac距離的差異進行比較如圖4A和4B所示,組間差異距離大于組內差異距離,表明5個分組多組比較的組間具有更顯著的差異。GL0_GL4組、GL1_GL4組和GL0_GL3組組間差異較大;GL0_GL1組組間差異較小,且GL2_GL4組存在異常值,其可能是因為兩組不同樣本之間比較的次數較多,若有個別數據與平均值差別較大則容易出現異常值,同樣表明連作逐步加大了鄰近沙壤土與連作覆土間的細菌組成差異。
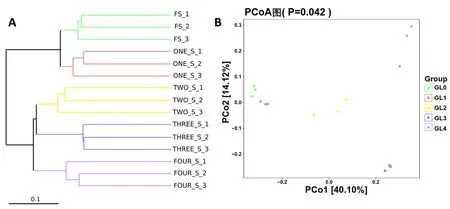
圖3 覆土細菌群落的UPGMA聚類和PCoA圖
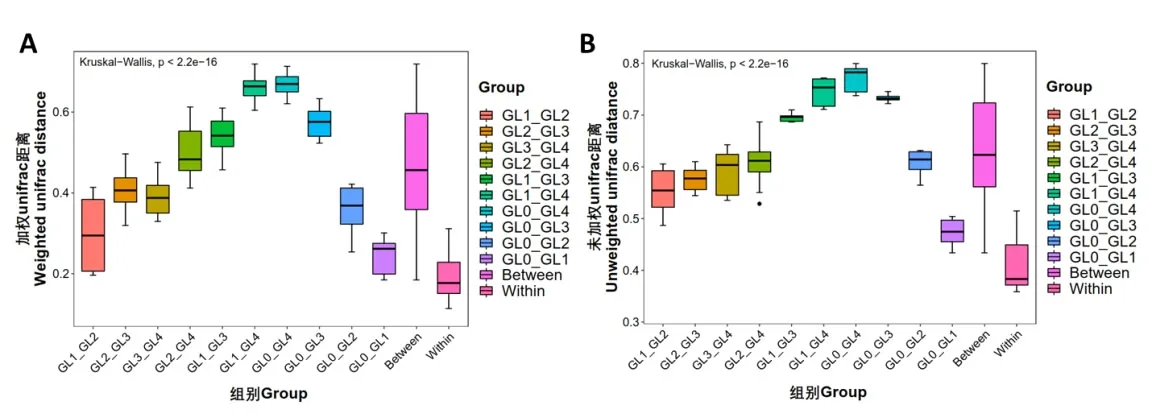
圖4 覆土細菌群落多組比較箱線圖
2.4 覆土細菌物種組成分析
2.4.1 門水平組成的分析 從15個靈芝土樣中一共獲得1050種細菌,分別隸屬于28門、89綱、169目、287科、624屬。如圖5A所示,變形菌門(Proteobacteria)、酸桿菌門(Acidobacteria)、放線菌門(Actinobacteria)和綠彎菌門(Chloroflex)的相對豐度約占細菌總序列的80%,表明5個分組中主要的優勢菌門為變形菌門、酸桿菌門、放線菌門和綠彎菌門。如圖5B的熱圖所示,隨著連作年限的增加,變形菌門、放線菌門和擬桿菌門(Bacteroidetes)的相對豐度顯著增 加 ;綠 彎 菌 門 (Chloroflex)、Bacteria_unclassified、candidate_division_WPS-2、糖化念珠菌(Candidatus_Saccharibacteria)和螺旋體門(Spirochaetes)的相對豐度顯著減少;而芽單胞菌門(Gemmatimonadetes)、厚壁菌門(Firmicutes)、硝化螺旋菌門(Nitrospirae)和疣微菌門(Verrucomicrobia)的相對豐度呈現先增加后減少的趨勢。其中,厚壁菌門和硝化螺旋菌門的相對豐度在GL2組開始下降,表明連作會影響靈芝覆土中優勢菌群的分布情況。
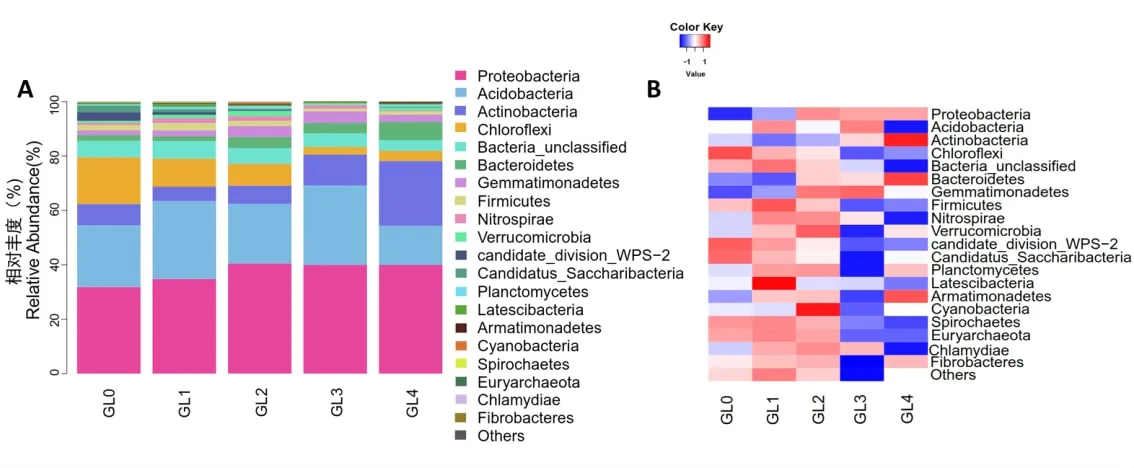
圖5 細菌群落門水平的相對豐富度
2.4.2 屬水平組成的分析 在屬水平上一共獲得624個屬,其相對豐度排名前20的優勢菌屬如圖6A所示。從圖中可以看出,在屬水平上檢出優勢細菌的相對豐度不斷減少,其他細菌相對豐度逐年增加。如圖6B所示,在靈芝栽培0、1、2、3和4年覆土中,變形菌門的溶桿菌屬(Lysobacter0.01%,0.11%,1.05%,1.14%,3.58%)與酸桿菌門的Gp6和Gp16的相對豐度隨連作年限的增加而不斷上升;變形菌門鞘氨醇單胞菌屬(Sphingomonas 5.33%,1.12%,3.09%,1.33%,2.71%)、Anaeromyxobacter(2.23% ,2.36% ,1.33% ,0.71% ,0.15%)、慢生根瘤菌屬(Bradyrhizobium 1.63%,1.54%,0.88%,0.75%,0.53%)和綠彎菌門脫氯菌屬(Dehalococcoides 8.89%,3.70%,2.02%,0.55%,0.27%)、Ktedonobacter(5.48%,3.66%,2.95%,0.09%,0.14%)的相對豐度不斷減少;變形菌門地桿菌屬(Geobacter)、假單胞菌屬(Pseudomonas)、紅游動菌屬(Rhodoplanes),酸 桿 菌 門 Gp1、Gp7、Edaphobacter、Gp3,芽單胞菌門芽單胞菌屬(Gemmatimonas)以及硝化螺旋菌門硝化螺旋菌屬(Nitrospira)相對豐度隨靈芝連作年限的增加呈現先增加后減少的趨勢。其中,地桿菌屬(Geobacter)、假單胞菌屬和Gp3的相對豐度在GL2組開始下降,表明長期連作會引起靈芝覆土中大量優勢細菌群落相對豐度的減少。
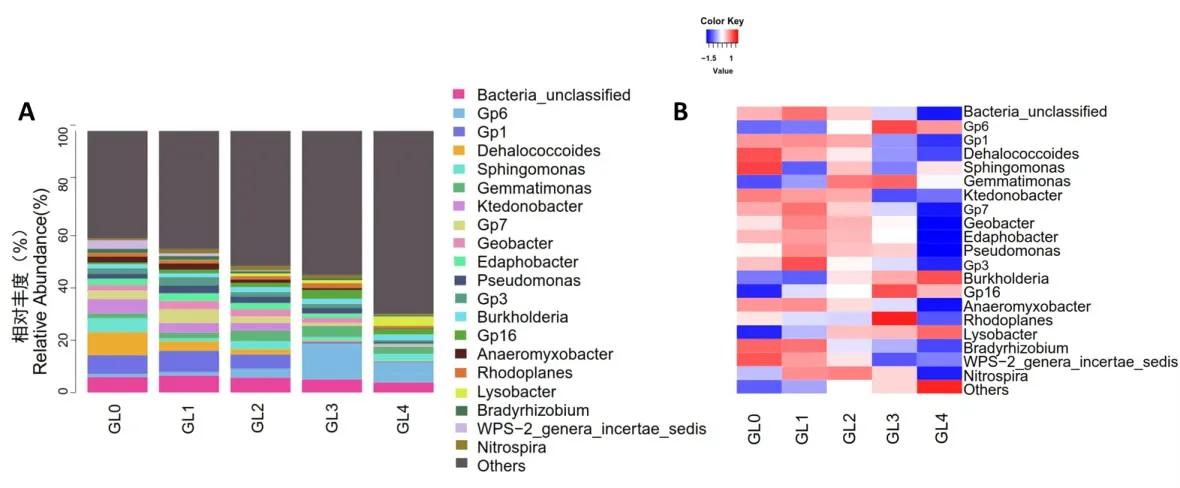
圖6 細菌群落屬水平的相對豐富度
3 討論
隨著國內健康產業的快速發展,靈芝產品因其較高的營養保健價值備受消費者喜愛,但人工栽培靈芝深受連作障礙現象的制約。以往研究發現,土壤微生物群落與連作障礙關系密切,常常表現為由“細菌型”轉向“真菌型”[8]。為了探索靈芝連作障礙的形成機制和防治措施,有人發現子囊菌門的木霉屬和青霉屬等真菌是靈芝主要的病原菌,并分析靈芝覆土微生態系統,提出對靈芝進行輪作、仿野生栽培、有益菌富集以及有機質使用的栽培方式[8,10]。吳曉明等[4]發現液氨熏蒸能有效降低靈芝覆土中的綠色木霉菌,不僅顯著提高了靈芝的生長速度,而且產量大約增長20.95%。在前期研究中,實驗室通過對靈芝連作覆土中的真菌群落多樣性進行分析,發現真菌群落的相對豐度逐年增加,并且在GL4組中青霉屬的相對豐度相對于GL0組約增加了57.92%[15]。李艷春等[20]發現在離茶樹大約25~30 cm的范圍內間作靈芝,能有效提高土壤中鞘氨醇單胞菌屬和伯克氏菌屬等有益細菌群落的相對豐度,顯著改善土壤的理化性質。然而,本研究發現靈芝連作覆土中鞘氨醇單胞菌屬等細菌群落相對豐度隨著連作年限的增加而遞減。
細菌能促進土地資源有效的循環利用,而連作障礙形成了特殊的土壤環境,會使細菌群落發生重要變化,因此解析靈芝連作覆土中細菌群落的變化情況對防治靈芝連作障礙具有重要指導意義[21]。在靈芝覆土中,變形菌門是5個分組中相對豐度最高的一類優勢菌群,約占細菌總量的34.77%~40.45%,并且其相對豐度隨著連作年限的增加而顯著增加。類似的,陳政在生姜連作土壤中也發現變形菌門的相對豐度位居第一,約占細菌類群總量的49.46%[22]。以往研究表明,變形菌門鞘氨醇單胞菌屬能促進芳香族化合物的降解,對于保護環境具有潛在的應用價值[23]。Hwang等[24]通過分析變形菌門Anaeromyxobacter Fw109-5的基因組序列,發現該菌株對地下污染物具有原位生物修復的潛力。而變形菌門慢生根瘤菌屬能較好的與降香黃檀等植物共生,具有高效的固氮能力[25]。研究結果同樣發現,變形菌門的鞘氨醇單胞菌屬、Anaeromyxobacter和慢生根瘤菌屬等作為靈芝生長過程中的優勢菌屬,隨連作的年限增加,其相對豐度均呈下降的趨勢。然而,變形菌門的溶桿菌屬是一種新型的生物防治藥物,在植物中具有防治病害和促進生長的功能[26]。其相對豐度在靈芝覆土中卻隨連作年限的增加而急劇增加,其在GL4組中大約提高了452倍,表明連作可能引發靈芝覆土中多數變形菌門優勢細菌屬的相對豐度下降。同時,在以往研究中,馬紅梅等[5]通過平板分離法發現放線菌相對豐度隨著連作年限的增加而呈現先增后減的趨勢,本研究發現放線菌相對豐度在栽培0、1、2、3和4年靈芝覆土中(7.86%,5.29%,6.72%,11.34%,23.90%)先減后增,其可能原因有待進一步探討。
酸桿菌門是靈芝覆土中僅次于變形菌門的第二優勢菌門,在土壤的循環利用和生態組建方面發揮著十分重要的作用[27]。結果發現酸桿菌門的Gp1、Gp7、Gp3和Edaphobacter的相對豐度呈現出先增后減的現象,表明長期連作會顯著降低靈芝覆土中酸桿菌門優勢菌屬的數量,不利于土壤資源的循環利用。在厭氧條件下,綠彎菌門的脫氯菌屬可對高氯代有機污染物進行脫氯并降解成為低氯代的產物,能有效的對污染的土壤進行修復和治理[28]。研究結果發現,靈芝覆土中脫氯菌屬的相對豐度逐年遞減,在GL4組中大約減少了96.96%。此外,芽單胞菌屬和硝化螺旋屬也是凈化環境常見的有益菌,田美等通過高通量測序技術發現芽單胞菌屬在污水凈化系統中發揮生物除磷作用[29]。Daims等[30]研究表明硝化螺旋屬可將亞硝酸鹽氧化成硝酸鹽,廣泛運用于水體凈化以及生態修復領域。然而,本研究中發現,覆土中芽單胞菌屬和硝化螺旋屬的細菌類群數量由于長期連作后也呈現大幅降低的趨勢。
此外,馬紅梅等[10]基于對峙平板培養法發現靈芝覆土中分離的分支桿菌屬、梭桿菌屬和產堿桿菌屬等細菌對靈芝菌絲體具有較強的化感作用,能顯著抑制靈芝菌絲體的生長。本研究進一步基于擴增子測序技術對靈芝覆土中的細菌群落的多樣性和結構組成進行系統的探究,其相比對傳統的平板分離法具有覆蓋面廣、通量高、準確和靈敏等特點,能全面系統反映覆土中細菌群落的結構[12]。實驗結果發現連作會加大靈芝覆土與鄰近沙壤土中細菌群落的物種組成差異,并且細菌群落的豐富度和多樣性呈現先增加后減少的趨勢。在屬水平上,靈芝覆土中的細菌總的相對豐度隨著連作年限的增加逐漸減少。其中,鞘氨醇單胞菌屬、Anaeromyxobacter、慢生根瘤菌屬和脫氯菌屬等土壤有益細菌的相對豐度逐年遞減,表明靈芝連作障礙與土壤中細菌群落的減少存在明顯的關聯性。本研究目前尚未對靈芝連作覆土中有益細菌群落的作用機制進行探討,有待進一步對其進行分離、回接和驗證。
4 結論
靈芝連作會逐年加大覆土中細菌群落的物種組成差異,其可能與靈芝栽培0、1、2、3和4年覆土中鞘氨醇單胞菌屬(Sphingomonas 5.33%,1.12%,3.09%,1.33%,2.71%)、Anaeromyxobacter(2.23% ,2.36% ,1.33% ,0.71%,0.15%)、慢生根瘤菌屬(Bradyrhizobium 1.63%,1.54%,0.88%,0.75%,0.53%)和脫氯菌屬(Dehalococcoides 8.89%,3.70%,2.02%,0.55%,0.27%)、Ktedonobacter(5.48%,3.66%,2.95%,0.09%,0.14%)等多種有益細菌相對豐度的減少有關,以期為靈芝連作障礙生態防控奠定理論基礎。
