蘋果葉片不定芽再生過(guò)程的差異表達(dá)基因鑒定與分析
2021-09-02 12:22:44劉鍇何閃閃張彩霞張利義卞書迅袁高鵬李武興康立群叢佩華韓曉蕾
中國(guó)農(nóng)業(yè)科學(xué) 2021年16期
劉鍇,何閃閃,張彩霞,張利義,卞書迅,袁高鵬,李武興,康立群,叢佩華,韓曉蕾
蘋果葉片不定芽再生過(guò)程的差異表達(dá)基因鑒定與分析
劉鍇,何閃閃,張彩霞,張利義,卞書迅,袁高鵬,李武興,康立群,叢佩華,韓曉蕾
中國(guó)農(nóng)業(yè)科學(xué)院果樹(shù)研究所/農(nóng)業(yè)部園藝作物種質(zhì)資源利用重點(diǎn)實(shí)驗(yàn)室/國(guó)家蘋果育種中心,遼寧興城 125100
【】篩選分析‘GL-3’蘋果葉片不定芽再生過(guò)程中的差異表達(dá)基因(differentially expressed gene,DEG),進(jìn)一步解析蘋果葉片不定芽再生的潛在分子機(jī)制,為提高蘋果的遺傳轉(zhuǎn)化效率提供理論參考。‘GL-3’蘋果繼代組培苗葉片外植體接種在再生培養(yǎng)基上,分別于0、3、7、14和21 d后取樣并提取RNA,構(gòu)建mRNA文庫(kù)后采用Illumina Nova seq平臺(tái)進(jìn)行測(cè)序。篩選出各時(shí)間點(diǎn)的DEGs,根據(jù)GO(Gene ontology)和KEGG(Kyoto encyclopedia of genes and genomes)注釋結(jié)果以及官方分類,使用R軟件中的phyper函數(shù)對(duì)篩選到的DEGs進(jìn)行GO和KEGG富集分析;利用BLAST軟件進(jìn)行基因比對(duì)注釋;重點(diǎn)分析植物再生相關(guān)的激素、酶、轉(zhuǎn)錄因子、多胺等DEGs;采用qRT-PCR對(duì)DEGs進(jìn)行定量驗(yàn)證。再生培養(yǎng)基上培養(yǎng)3、7、14和21 d的蘋果葉片外植體與對(duì)照組相比,分別篩選到5 250、4 937、6 852、6 493個(gè)DEGs,4個(gè)時(shí)間點(diǎn)共有的DEGs有3 027個(gè)。DEGs的GO功能富集顯示,4個(gè)時(shí)間點(diǎn)篩選到的共有DEGs中上調(diào)表達(dá)的DEGs主要與氧化還原過(guò)程、細(xì)胞外圍、蛋白激酶活性和有機(jī)環(huán)化合物結(jié)合等功能有關(guān);下調(diào)表達(dá)的DEGs主要與單細(xì)胞代謝過(guò)程、鈣離子結(jié)合、光合膜和類囊體部分等功能有關(guān)。DEGs的KEGG通路富集分析顯示,4個(gè)時(shí)間點(diǎn)篩選到的共有DEGs中上調(diào)表達(dá)的DEGs主要富集在磷酸戊糖途徑、植物激素信號(hào)轉(zhuǎn)導(dǎo)、植物-病原菌相互作用和內(nèi)質(zhì)網(wǎng)蛋白質(zhì)加工等途徑中;下調(diào)表達(dá)的DEGs主要富集在-亞麻酸代謝、苯丙烷生物合成、碳代謝和光合作用等途徑中。對(duì)與植物離體葉片再生相關(guān)的激素、酶、轉(zhuǎn)錄因子和多胺等相關(guān)DEGs的表達(dá)模式進(jìn)行分析發(fā)現(xiàn),這些DEGs大部分呈上調(diào)表達(dá)趨勢(shì)。經(jīng)qRT-PCR驗(yàn)證后,所檢測(cè)基因的表達(dá)趨勢(shì)與轉(zhuǎn)錄組測(cè)序結(jié)果一致。通過(guò)對(duì)蘋果葉片不定芽再生過(guò)程中不同時(shí)間點(diǎn)的基因表達(dá)譜進(jìn)行檢測(cè)和對(duì)比分析,獲得了大量與蘋果葉片不定芽再生相關(guān)的基因,研究結(jié)果為深入探討蘋果離體葉片再生機(jī)理提供了理論依據(jù)。
蘋果;不定芽再生;RNA-Seq;差異表達(dá)基因;影響因子
0 引言
【研究意義】蘋果是廣受歡迎的大眾水果,在全球溫帶地區(qū)廣泛種植,我國(guó)蘋果產(chǎn)業(yè)發(fā)展迅速,據(jù)2016年統(tǒng)計(jì)數(shù)據(jù)顯示,我國(guó)蘋果栽培面積和產(chǎn)量均居世界首位,已成為世界上最大的蘋果生產(chǎn)國(guó)[1]。蘋果分子生物學(xué)[1]在栽培蘋果()測(cè)序完成后發(fā)展迅速[2],分子輔助選擇技術(shù)也逐步應(yīng)用到蘋果育種中,提高了育種效率,縮短了育種周期[3]。然而蘋果的遺傳轉(zhuǎn)化效率低,嚴(yán)重制約著蘋果分子生物學(xué)的發(fā)展[4-5]。根癌農(nóng)桿菌介導(dǎo)的遺傳轉(zhuǎn)化體系是目前蘋果遺傳轉(zhuǎn)化的最優(yōu)體系,主要以離體葉片作為基因轉(zhuǎn)化的受體材料,離體葉片不定芽再生能力是影響蘋果遺傳轉(zhuǎn)化效率的關(guān)鍵因素之一[6],然而蘋果離體葉片不定芽再生的分子機(jī)制目前尚不明確。葉外植體的芽再生與基因型密切相關(guān)[7-8],研究發(fā)現(xiàn),‘嘎拉’‘喬納金’和‘自由’等品種的再生能力較強(qiáng),而‘富士’‘平邑甜茶’等品種的離體葉片很難再生不定芽[9]。‘GL-3’是由沈陽(yáng)農(nóng)業(yè)大學(xué)選育的‘嘎拉’實(shí)生后代,具有再生能力強(qiáng)和對(duì)農(nóng)桿菌敏感的特性[6]。近年來(lái),RNA-Seq在植物再生關(guān)鍵基因挖掘和植物再生機(jī)制研究中發(fā)揮著越來(lái)越重要的作用[10]。利用RNA-Seq鑒定和分析‘GL-3’蘋果離體葉片再生不定芽過(guò)程中的差異表達(dá)基因,進(jìn)一步解析蘋果再生調(diào)控過(guò)程的分子機(jī)制,為提高蘋果遺傳轉(zhuǎn)化效率奠定理論基礎(chǔ)。【前人研究進(jìn)展】離體葉片不定芽再生是一個(gè)多級(jí)發(fā)育過(guò)程,其中包括體細(xì)胞對(duì)植物激素信號(hào)的感知與傳遞,啟動(dòng)細(xì)胞分裂增殖,獲得具有器官再生能力的去分化以及形成器官的再分化等,這是一個(gè)復(fù)雜的基因調(diào)控過(guò)程[11]。近年來(lái),已鑒定出許多植物不定芽離體發(fā)生過(guò)程中的關(guān)鍵基因和轉(zhuǎn)錄因子。Che等[12]利用RNA-Seq分析了擬南芥不定芽離體發(fā)生過(guò)程中的基因表達(dá)譜。在愈傷組織形成過(guò)程中,IAA-氨基合成酶基因、GCN5-relatedN-乙酰基轉(zhuǎn)移酶基因等顯著上調(diào)表達(dá);在不定芽分化過(guò)程中,與細(xì)胞分裂素信號(hào)轉(zhuǎn)導(dǎo)有關(guān)的基因如等差異表達(dá)最為顯著。研究發(fā)現(xiàn),擬南芥S突變體完全喪失芽再生能力,表明是調(diào)控芽再生的關(guān)鍵基因[13]。過(guò)量表達(dá)植物年齡相關(guān)miR156亞家族基因可以顯著提高離體葉片的莖尖再生數(shù)[14]。最新研究發(fā)現(xiàn),介導(dǎo)的組蛋白H3K4me2去乙酰化是離體器官不定芽再生的重要條件,敲除突變體的芽再生能力顯著下降[15]。【本研究切入點(diǎn)】當(dāng)前對(duì)蘋果葉片不定芽再生能力的研究多集中在植株基因型和不定芽再生體系創(chuàng)建上,相關(guān)的分子機(jī)制研究較少,對(duì)蘋果再生調(diào)控過(guò)程的分子機(jī)制尚不清晰。【擬解決的關(guān)鍵問(wèn)題】本研究以遺傳轉(zhuǎn)化效率高、再生能力強(qiáng)的‘GL-3’蘋果葉片為研究試材,對(duì)葉片外植體不定芽再生過(guò)程中不同時(shí)間點(diǎn)的試材進(jìn)行轉(zhuǎn)錄組測(cè)序,通過(guò)差異基因篩選、GO富集分析、KEGG富集分析和關(guān)鍵基因表達(dá)模式分析,解析蘋果葉片不定芽再生的分子機(jī)制。
1 材料與方法
試驗(yàn)于2019年8月至2020年4月在中國(guó)農(nóng)業(yè)科學(xué)院果樹(shù)研究所/農(nóng)業(yè)部園藝作物種質(zhì)資源利用重點(diǎn)實(shí)驗(yàn)室/蘋果育種中心實(shí)驗(yàn)室進(jìn)行。
1.1 葉片接種培養(yǎng)
本試驗(yàn)所用植物材料為沈陽(yáng)農(nóng)業(yè)大學(xué)果樹(shù)分子生物學(xué)實(shí)驗(yàn)室惠贈(zèng)的蘋果‘GL-3’組培苗。外植體選擇繼代培養(yǎng)30 d左右的‘GL-3’組培苗頂部的幼嫩葉片,且葉片生理狀態(tài)基本一致。將葉片垂直主脈橫切3—5刀(不切斷葉緣)后,葉背朝下接種于再生培養(yǎng)基上培養(yǎng)(25℃、避光)。再生培養(yǎng)基配方為:MS+TDZ 2.0 mg?L-1+NAA 0.1 mg?L-1+蔗糖35 g?L-1+瓊脂6.2 g?L-1,pH 5.8。分別于0、3、7、14和21 d后取樣。0 d取樣的葉片作為對(duì)照組,3、7、14和21 d取樣的葉片作為處理組,對(duì)照組及處理組均設(shè)置3次生物學(xué)重復(fù)。
1.2 轉(zhuǎn)錄組測(cè)序
提取上述樣品的總RNA,檢測(cè)合格后委托北京貝瑞和康生物技術(shù)有限公司采用Illumina Nova seq測(cè)序平臺(tái)進(jìn)行二代雙向測(cè)序。測(cè)序得到的數(shù)據(jù)經(jīng)堿基識(shí)別分析后得到原始數(shù)據(jù)raw reads,過(guò)濾掉低質(zhì)量、含N比例大于10%、接頭污染的reads后得到適合分析的數(shù)據(jù)clean reads。使用比對(duì)軟件BWA[16]將數(shù)據(jù)比對(duì)‘金冠’蘋果基因組。
1.3 差異表達(dá)基因的篩選
使用基于泊松分布的DEGseq算法[13]進(jìn)行DEGs的檢測(cè),將差異倍數(shù)為兩倍以上(fold change,F(xiàn)C≥2)且Q-value≤0.001(adjusted P-value≤0.001)的基因定義為顯著差異表達(dá)基因,將每個(gè)時(shí)間點(diǎn)差異倍數(shù)最大且FPKM≥10的30個(gè)DEGs定義為極顯著差異表達(dá)基因。
1.4 差異表達(dá)基因的功能分析
根據(jù)GO和KEGG注釋結(jié)果以及官方分類,使用R軟件中的phyper函數(shù)對(duì)篩選到的DEGs進(jìn)行GO和KEGG富集分析。
1.5 差異基因的qRT-PCR熒光定量分析
反轉(zhuǎn)錄采用PrimeScriptTMRT reagent Kit with gDNA Eraser試劑盒(TaKaRa公司),qRT-PCR熒光定量分析采用TB Green染料。用蘋果肌動(dòng)蛋白基因?yàn)閮?nèi)標(biāo)參照[17],熒光定量引物由金唯智生物科技有限公司合成(引物序列見(jiàn)表1),樣品3次重復(fù),基因表達(dá)倍數(shù)通過(guò)2-△△Ct法計(jì)算[18]。

表1 差異表達(dá)基因的定量引物驗(yàn)證
2 結(jié)果
2.1 離體葉片不定芽再生過(guò)程的形態(tài)變化
對(duì)再生培養(yǎng)基上培養(yǎng)的離體葉片狀態(tài)進(jìn)行定期觀察,結(jié)果如圖1所示,葉片剛接種到培養(yǎng)基時(shí)葉面平整,緊貼培養(yǎng)基;3 d時(shí)葉片切口處微微隆起,葉片膨脹變大;7 d時(shí)葉片切口處隆起更加明顯,且切口處已有少許愈傷組織形成,葉片繼續(xù)膨脹變大;14 d時(shí)葉片切口處的愈傷組織變密變多,且有少量芽點(diǎn)出現(xiàn),葉片不再膨脹變大;21 d時(shí)不定芽數(shù)量明顯增多,并開(kāi)始變綠和進(jìn)行伸長(zhǎng)生長(zhǎng)。
2.2 差異表達(dá)基因篩選
對(duì)‘GL-3’蘋果葉片不定芽再生過(guò)程中的處理組與對(duì)照組的差異表達(dá)基因(差異表達(dá)倍數(shù)FC≥2及Q-value≤0.001)進(jìn)行篩選。再生培養(yǎng)基上培養(yǎng)3 d的葉片外植體與對(duì)照組相比共篩選到5 250個(gè)DEGs,其中1 681個(gè)上調(diào)表達(dá),3 569個(gè)下調(diào)表達(dá);培養(yǎng)7 d的葉片外植體與對(duì)照組相比,共篩選到4 937個(gè)DEGs,其中1 882個(gè)上調(diào)表達(dá),3 055個(gè)下調(diào)表達(dá);培養(yǎng)14 d的葉片外植體與對(duì)照組相比共篩選到6 852個(gè)DEGs,其中2 705個(gè)上調(diào)表達(dá),4 147個(gè)下調(diào)表達(dá);培養(yǎng)21 d的蘋果葉片外植體與對(duì)照組相比共篩選到6 493個(gè)DEGs,其中2 602個(gè)上調(diào)表達(dá),3 891個(gè)下調(diào)表達(dá)(圖2-A)。基因表達(dá)水平通過(guò)RPKM方法進(jìn)行計(jì)算(圖2-B)。對(duì)這些DEGs作韋恩圖進(jìn)行分析,葉片外植體接種于再生培養(yǎng)基3、7、14和21 d時(shí)共有的DEGs有3 027個(gè)(圖2-C)。這些數(shù)據(jù)表明‘GL-3’蘋果離體葉片不定芽再生過(guò)程在轉(zhuǎn)錄水平上發(fā)生明顯變化。

圖1 離體葉片不定芽再生過(guò)程的形態(tài)變化
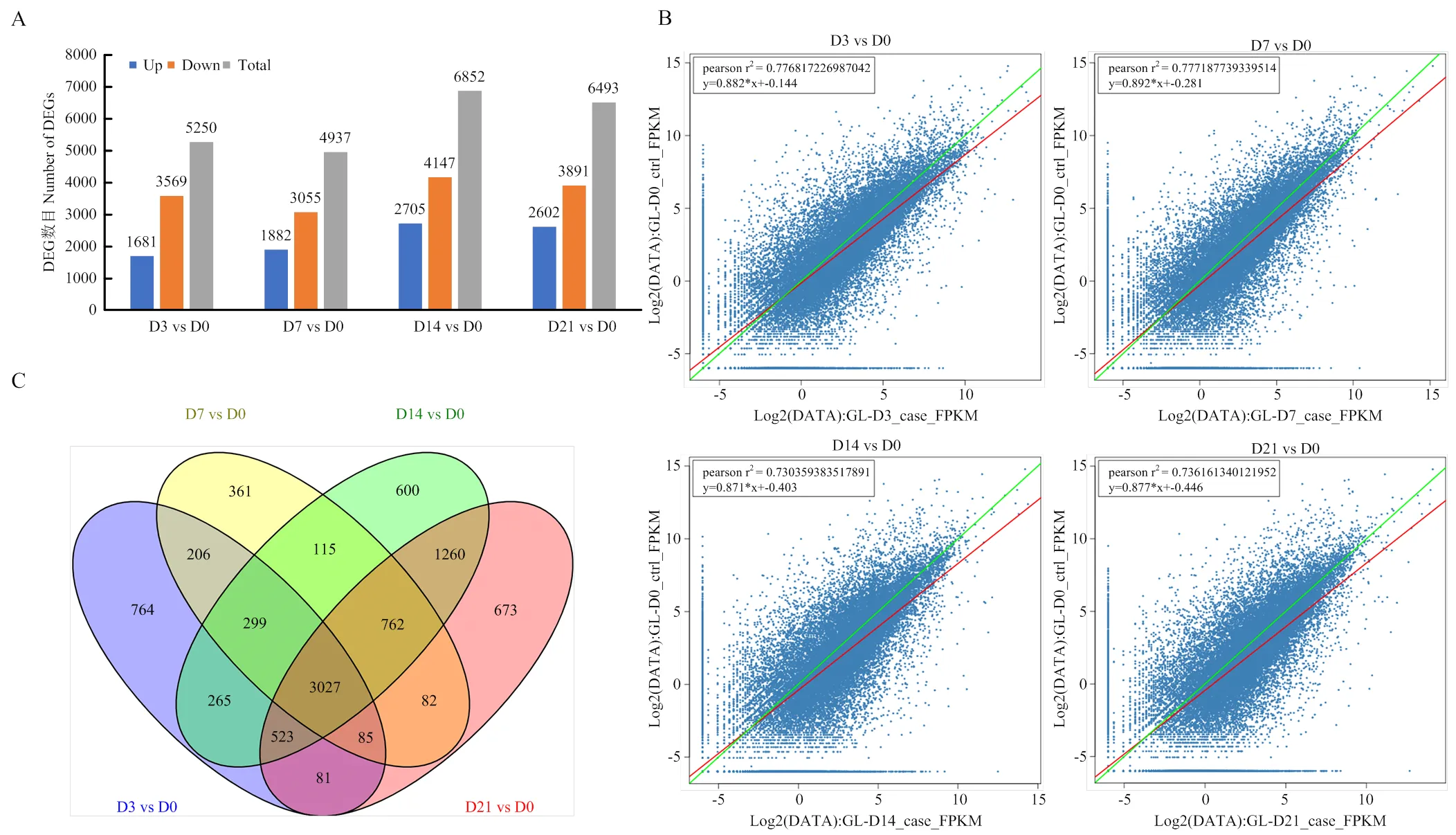
A:各時(shí)間點(diǎn)差異表達(dá)基因上下調(diào)數(shù)量統(tǒng)計(jì);B:各時(shí)間點(diǎn)差異表達(dá)的散點(diǎn)圖;C:各時(shí)間點(diǎn)差異表達(dá)基因韋恩圖
2.3 4個(gè)時(shí)間點(diǎn)共有差異表達(dá)基因的GO功能富集分析
葉片外植體接種于再生培養(yǎng)基3、7、14和21 d時(shí)共有的DEGs有3 027個(gè),其中1 046個(gè)上調(diào)表達(dá),1 981個(gè)下調(diào)表達(dá)。GO富集結(jié)果顯示,上調(diào)表達(dá)的DEGs主要與氧化還原過(guò)程(oxidation reduction process)、細(xì)胞外圍(cell periphery)、蛋白激酶活性(protein kinase activity)和有機(jī)環(huán)化合物結(jié)合(organic cyclic compound binding)等功能有關(guān)(圖3-A);下調(diào)表達(dá)的DEGs主要與單細(xì)胞代謝過(guò)程(single organism metabolic process)、鈣離子結(jié)合(calcium ion binding)、光合膜(photosynthetic membrane)和類囊體部分(thylakoid part)等功能有關(guān)(圖3-B)。

A、B分別為4個(gè)時(shí)間點(diǎn)共有的差異表達(dá)基因中上調(diào)表達(dá)和下調(diào)表達(dá)差異基因的GO富集結(jié)果
2.4 4個(gè)時(shí)間點(diǎn)共有差異表達(dá)基因的KEGG功能富集分析
對(duì)4個(gè)時(shí)間點(diǎn)共有差異表達(dá)基因進(jìn)行KEGG富集分析發(fā)現(xiàn),上調(diào)表達(dá)的DEGs主要富集在磷酸戊糖途徑(pentose phosphate pathway)、植物激素信號(hào)轉(zhuǎn)導(dǎo)(plant hormone signal transduction)、植物-病原菌相互作用(plant pathogen interaction)和內(nèi)質(zhì)網(wǎng)蛋白質(zhì)加工(protein processing in endoplasmic reticulum)等途徑中(圖4-A);下調(diào)表達(dá)的DEGs主要富集在-亞麻酸代謝(alpha linolenic acid metabolism)、苯丙烷生物合成(phenylpropanoid biosynthesis)、碳代謝(carbon metabolism)和光合作用(photosynthesis)等途徑中(圖4-B)。
2.5 極顯著差異表達(dá)基因的功能分析
4個(gè)時(shí)間點(diǎn)共有的極顯著差異表達(dá)基因主要與細(xì)胞分裂素代謝過(guò)程、氧化還原過(guò)程和磷酸化信號(hào)轉(zhuǎn)導(dǎo)系統(tǒng)等功能有關(guān),顯著富集在玉米素生物合成、植物激素信號(hào)轉(zhuǎn)導(dǎo)、生物堿的生物合成及戊糖及葡萄糖醛酸轉(zhuǎn)換等途徑中(表2)。
2.6 植物再生相關(guān)差異表達(dá)基因分析
2.6.1 植物激素相關(guān)差異表達(dá)基因分析 外源生長(zhǎng)物質(zhì)對(duì)離體器官分化的誘導(dǎo)作用依賴于對(duì)內(nèi)源激素平衡的調(diào)節(jié)。生長(zhǎng)素(IAA)和細(xì)胞分裂素(CTK)與植物不定芽再生關(guān)系最為密切,對(duì)IAA和CTK相關(guān)的DEGs的表達(dá)模式進(jìn)行分析,發(fā)現(xiàn)IAA和CTK相關(guān)的DEGs上調(diào)表達(dá)和下調(diào)表達(dá)的數(shù)目大致相等,且IAA相關(guān)的DEGs明顯多于CTK相關(guān)的DEGs(圖5)。
2.6.2 轉(zhuǎn)錄因子相關(guān)差異表達(dá)基因分析 轉(zhuǎn)錄因子在植物離體葉片不定芽再生過(guò)程中發(fā)揮至關(guān)重要的作用。在蘋果葉片外植體再生過(guò)程的各時(shí)間點(diǎn)均檢測(cè)到大量差異表達(dá)的轉(zhuǎn)錄因子,其中以AP2-EREBP、ARR、HD-ZIP、bZIP、LBD為主,并且這些轉(zhuǎn)錄因子主要呈上調(diào)表達(dá)趨勢(shì)(圖6)。

表2 極顯著差異表達(dá)基因的功能分析
2.6.3 酶相關(guān)差異表達(dá)基因分析 植物離體器官再生相關(guān)的酶類主要有超氧化物歧化酶(SOD)、過(guò)氧化物酶(POD)、過(guò)氧化氫酶(CAT)。4個(gè)時(shí)間點(diǎn)篩選到的編碼POD的DEGs最多,其次是編碼SOD的DEGs,篩選到編碼CAT的DEGs最少。對(duì)編碼這些酶的DEGs的表達(dá)模式進(jìn)行分析,編碼SOD的DEGs主要呈上調(diào)表達(dá)趨勢(shì),編碼POD、CAT的DEGs主要呈下調(diào)表達(dá)趨勢(shì)(圖7)。

圖7 編碼POD、SOD、CAT的DEGs的熱圖
2.6.4 多胺分析 植物離體器官再生相關(guān)的多胺主要有腐胺(Put)、精胺(Spm)和亞精胺(Spd)。對(duì)編碼這些多胺的DEGs表達(dá)模式進(jìn)行分析,編碼這3類多胺的DEGs主要呈上調(diào)表達(dá)趨勢(shì)且上調(diào)表達(dá)的倍數(shù)較小,21 d時(shí),一半的DEGs下調(diào)表達(dá)(圖8)。
2.7 差異表達(dá)基因的qRT-PCR檢測(cè)
選擇IAA相關(guān)基因、和,CTK相關(guān)基因,SOD相關(guān)基因,POD相關(guān)基因、,編碼植物再生相關(guān)轉(zhuǎn)錄因子的基因、、、、進(jìn)行qRT-PCR驗(yàn)證。選定的12個(gè)(8個(gè)上調(diào)表達(dá),4個(gè)下調(diào)表達(dá))DEGs都在對(duì)照組和處理組之間差異表達(dá),差異倍數(shù)較大且與分析結(jié)果密切相關(guān),qRT-PCR驗(yàn)證結(jié)果與轉(zhuǎn)錄組測(cè)序結(jié)果基本一致(圖9)。
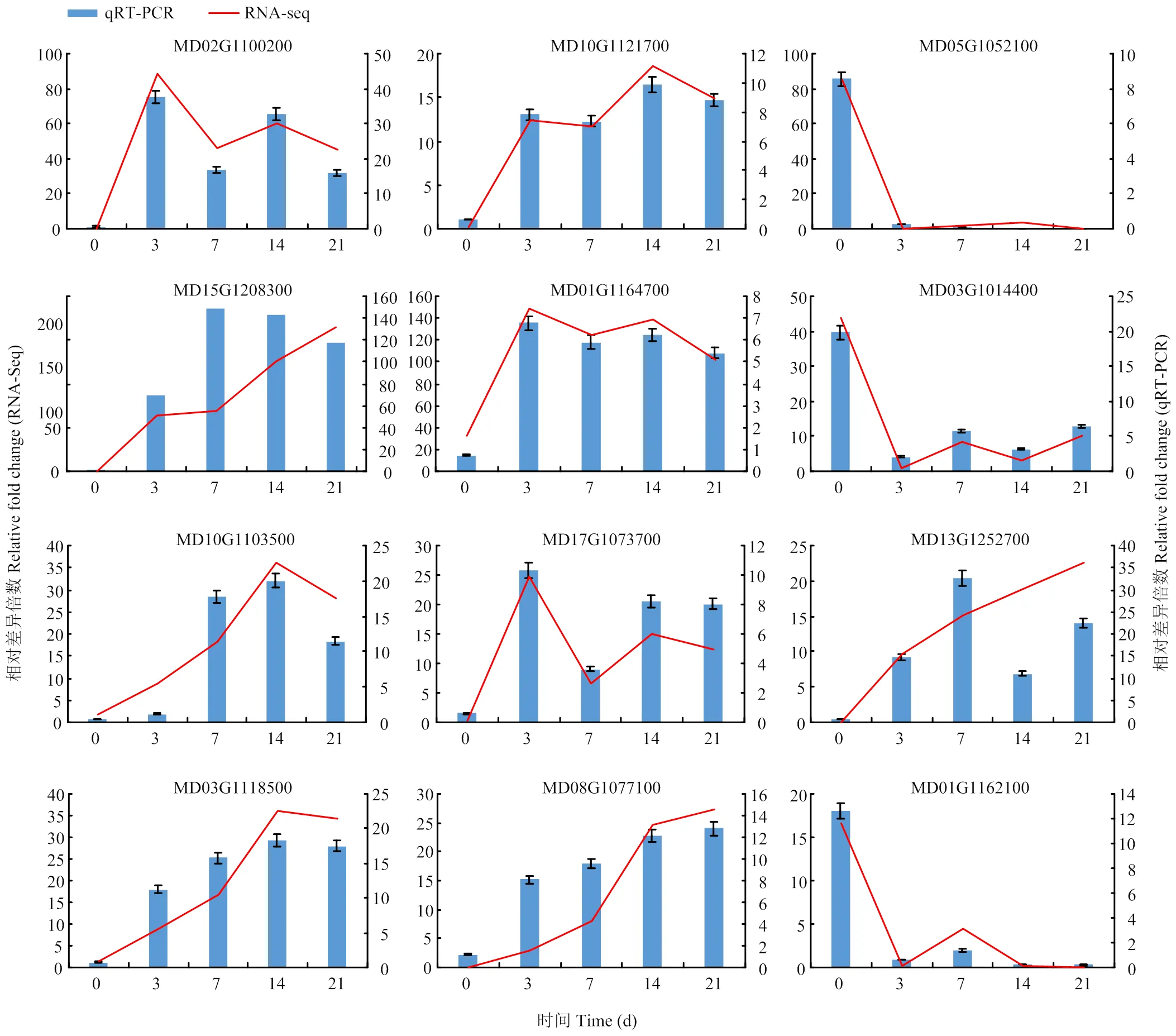
圖9 差異表達(dá)基因的qRT-PCR驗(yàn)證
3 討論
蘋果遺傳轉(zhuǎn)化效率低下,嚴(yán)重限制了蘋果功能基因組學(xué)研究的進(jìn)程,進(jìn)而阻礙了蘋果分子生物學(xué)的發(fā)展。葉片外植體再生困難是導(dǎo)致蘋果遺傳轉(zhuǎn)化效率低的主要原因之一,然而離體葉片再生不定芽的分子機(jī)制目前尚不明確。離體葉片不定芽再生是一個(gè)多級(jí)發(fā)育過(guò)程,涉及大量的轉(zhuǎn)錄和重編程[19-20]。利用RNA-Seq技術(shù)從全局角度分析蘋果葉片不定芽再生的分子機(jī)制,為建立一個(gè)高效、穩(wěn)定的離體葉片不定芽再生體系提供理論參考,對(duì)于蘋果遺傳轉(zhuǎn)化效率的提高、基因功能驗(yàn)證和性狀改良等方面具有重要意義。
植物離體葉片不定芽發(fā)生分為兩種途徑:一種是通過(guò)愈傷組織分化產(chǎn)生不定芽,一種是由葉片直接再生不定芽[21]。蘋果的遺傳轉(zhuǎn)化體系主要通過(guò)愈傷組織上再生不定芽的方式再生植株[6]。愈傷組織形成和不定芽再生是不定芽形成的2個(gè)關(guān)鍵階段,大量基因在這2個(gè)階段顯著差異表達(dá)并對(duì)細(xì)胞去分化和再分化的過(guò)程進(jìn)行調(diào)控[22],本研究對(duì)這2個(gè)階段基因的差異表達(dá)情況進(jìn)行了分析。0—14 d是愈傷組織形成階段,在3 d和7 d分別篩選到5 250和4 937個(gè)DEGs,推測(cè)這些DEGs與誘導(dǎo)愈傷組織形成相關(guān)。14—21 d愈傷組織分化產(chǎn)生不定芽,在14 d和2 1d分別篩選到6 852和6 493個(gè)DEGs,推測(cè)這些DEGs與愈傷組織分化產(chǎn)生不定芽相關(guān)。在14 d和21 d篩選到的DEGs明顯多于3 d和7 d,推測(cè)14 d時(shí)離體葉片細(xì)胞基因表達(dá)程序發(fā)生劇烈變化,這些14 d才開(kāi)始差異表達(dá)的基因在愈傷組織向不定芽分化的過(guò)程中起到重要的調(diào)控作用。
植物已分化的細(xì)胞經(jīng)過(guò)脫分化變成愈傷組織細(xì)胞,細(xì)胞內(nèi)部會(huì)發(fā)生一系列反應(yīng),造成異染色質(zhì)發(fā)生大規(guī)模解聚和重新排布,外端粒的長(zhǎng)度、端粒酶的活性也會(huì)發(fā)生改變[23]。細(xì)胞通過(guò)磷酸戊糖途徑可以產(chǎn)生大量的NADPH,為各種反應(yīng)提供還原劑[24]。推測(cè)磷酸戊糖途徑在蘋果離體葉片愈傷組織形成過(guò)程中發(fā)揮重要作用。植物細(xì)胞向愈傷組織細(xì)胞轉(zhuǎn)化過(guò)程中,細(xì)胞壁會(huì)發(fā)生大規(guī)模的解離[25]。木質(zhì)素主要是由苯丙烷類組成的復(fù)雜化合物[26],是構(gòu)成植物細(xì)胞壁的成分之一[27]。由此推測(cè)苯丙烷生物合成相關(guān)的DEGs下調(diào)表達(dá)促進(jìn)了植物細(xì)胞壁的解離過(guò)程,加速已分化細(xì)胞完成脫分化。研究發(fā)現(xiàn),脂肪酸會(huì)對(duì)中柱鞘類細(xì)胞分生能力關(guān)鍵基因的表達(dá)進(jìn)行抑制,從而對(duì)愈傷組織形成過(guò)程進(jìn)行調(diào)控[28]。推測(cè)-亞麻酸是植物體內(nèi)限制愈傷組織形成的關(guān)鍵信號(hào)物質(zhì)。由于愈傷組織再生不定芽的過(guò)程快速且不易觀察,此過(guò)程控制細(xì)胞譜系發(fā)展的機(jī)制并不十分清晰[29]。本研究對(duì)14 d和21 d篩選到的DEGs進(jìn)行功能分析發(fā)現(xiàn),DEGs主要與單細(xì)胞代謝過(guò)程、質(zhì)體、ATP結(jié)合和有機(jī)環(huán)結(jié)合等功能相關(guān)。推測(cè)這些功能在離體葉片細(xì)胞從脫分化向再分化轉(zhuǎn)變,完成不定芽的再生過(guò)程中發(fā)揮重要作用。
細(xì)胞分裂素、生長(zhǎng)素是外植體再生不定芽所必需的,細(xì)胞分裂素與生長(zhǎng)素比值高有助于外植體分化不定芽,而外源生長(zhǎng)物質(zhì)必須通過(guò)對(duì)內(nèi)源激素平衡的調(diào)節(jié)才發(fā)揮作用,從而影響器官分化[30]。本研究發(fā)現(xiàn)生長(zhǎng)素和細(xì)胞分裂素相關(guān)的DEGs上調(diào)表達(dá)和下調(diào)表達(dá)的數(shù)目大致相等,篩選到的與生長(zhǎng)素相關(guān)的DEGs數(shù)目明顯多于細(xì)胞分裂素。研究發(fā)現(xiàn),生長(zhǎng)素通過(guò)自身極性運(yùn)輸調(diào)節(jié)自身濃度梯度分布并決定干細(xì)胞的位置[31],生長(zhǎng)素主要分布于將要產(chǎn)生莖頂端分生組織的愈傷組織周圍,在離體器官再生過(guò)程中起到了決定性的作用[32]。推測(cè)植物主要通過(guò)調(diào)節(jié)內(nèi)源生長(zhǎng)素的濃度變化來(lái)促進(jìn)離體葉片不定芽的再生。篩選到的生長(zhǎng)素相關(guān)的DEGs中,MD02G1100200差異表達(dá)最為顯著,在21 d和0 d的表達(dá)量相差200多倍,該基因序列與擬南芥中的序列高度相似。是生長(zhǎng)素早期響應(yīng)基因,其蛋白產(chǎn)物能夠特異性結(jié)合生長(zhǎng)素響應(yīng)因子ARF,進(jìn)而調(diào)控生長(zhǎng)素響應(yīng)基因的表達(dá),在整個(gè)植物生長(zhǎng)素信號(hào)轉(zhuǎn)導(dǎo)過(guò)程中具有重要作用[33]。篩選到的細(xì)胞分裂素相關(guān)的DEGs中,MD15G1208300差異表達(dá)最為顯著,在21 d和0 d的表達(dá)量相差160多倍,該基因序列與擬南芥中的序列高度相似,編碼細(xì)胞分裂素氧化酶(CKX),在植物細(xì)胞內(nèi)的細(xì)胞分裂素濃度調(diào)節(jié)中發(fā)揮重要作用[34]。推測(cè)MD02G1100200和MD15G1208300分別通過(guò)對(duì)生長(zhǎng)素和細(xì)胞分裂素濃度的調(diào)節(jié)在蘋果葉片再生不定芽的過(guò)程中發(fā)揮重要作用。植物中存在多個(gè)轉(zhuǎn)錄因子家族,相關(guān)報(bào)道表明AP2、LBD、bZIP、HD-ZIP、ARR家族的轉(zhuǎn)錄因子在不定芽再生過(guò)程起主要的轉(zhuǎn)錄調(diào)控作用[35]。LBD類轉(zhuǎn)錄因子中的LBD16、LBD17、LBD18受到生長(zhǎng)素信號(hào)因子ARF7和ARF19的調(diào)控,超表達(dá)其中任何一個(gè)LBD轉(zhuǎn)錄因子都能促進(jìn)植物外植體愈傷組織的形成[36]。擬南芥LBD轉(zhuǎn)錄因子還可以與bZIP家族轉(zhuǎn)錄因子bZIP59形成轉(zhuǎn)錄復(fù)合體來(lái)調(diào)控愈傷組織形成[37]。是植物不定芽再生的關(guān)鍵基因[38],細(xì)胞分裂素信號(hào)途徑關(guān)鍵轉(zhuǎn)錄因子B型ARR可以直接結(jié)合到啟動(dòng)子區(qū)域,激活的表達(dá)[39]。此外,B型ARRs還可以與HD-ZIPIII特異性結(jié)合,形成轉(zhuǎn)錄復(fù)合體激活表達(dá)[22]。AP2家族轉(zhuǎn)錄因子有144個(gè)成員,參與植物多個(gè)生長(zhǎng)發(fā)育過(guò)程[40],多個(gè)AP2家族成員參與植物離體器官不定芽再生過(guò)程,如生長(zhǎng)素可以誘導(dǎo)、和的表達(dá),而3個(gè)轉(zhuǎn)錄因子又可激活根分生組織的特征基因和以及芽再生特征基因和的表達(dá),從而賦予愈傷組織再生芽的能力[41]。本研究篩選到了大量差異表達(dá)的AP2、LBD、bZIP、ARR家族轉(zhuǎn)錄因子,并且這些轉(zhuǎn)錄因子主要呈上調(diào)表達(dá)趨勢(shì),推測(cè)這些轉(zhuǎn)錄因子在一定程度上參與了葉片不定芽再生的調(diào)控過(guò)程。植物離體培養(yǎng)過(guò)程中,酶的活性和種類的變化會(huì)對(duì)離體器官的發(fā)生產(chǎn)生重要影響,與離體器官發(fā)生相關(guān)的酶類主要有SOD[42]、POD[43]、CAT[44]等。SOD在植物脫分化及愈傷組織、根、芽分化期間的活性一般增強(qiáng),可以將IAA氧化分解,從而影響植物體內(nèi)激素的平衡,是離體器官發(fā)生過(guò)程中重要的酶[45]。本研究篩選到的SOD相關(guān)的DEGs在不定芽再生過(guò)程中顯著上調(diào)表達(dá)。推測(cè)SOD通過(guò)對(duì)內(nèi)源生長(zhǎng)素濃度的調(diào)節(jié)影響蘋果葉片離體再生不定芽的過(guò)程。多胺是生物體內(nèi)的一類活性物質(zhì),在植物細(xì)胞中主要有腐胺、精胺和亞精胺[46]。研究發(fā)現(xiàn),內(nèi)源激素對(duì)外植體形態(tài)發(fā)生的影響可能通過(guò)多胺來(lái)實(shí)現(xiàn)或協(xié)同作用,多胺在調(diào)控形態(tài)發(fā)生時(shí)主要起第二信使的作用[47-48]。本研究篩選到的腐胺、精胺和亞精胺相關(guān)的DEGs均顯著上調(diào)表達(dá),推測(cè)在蘋果葉片離體再生過(guò)程中,植物激素變化引起細(xì)胞內(nèi)多胺變化,隨后引起生理生化變化,最終影響到形態(tài)發(fā)生。
4 結(jié)論
再生培養(yǎng)基上培養(yǎng)3、7、14和21 d的蘋果葉片外植體與對(duì)照組相比,分別篩選到5 250、4 937、6 852和6 493個(gè)DEGs,4個(gè)時(shí)間點(diǎn)共有的DEGs有3 027個(gè)。4個(gè)時(shí)間點(diǎn)共有的DEGs主要與分子功能、細(xì)胞成分和生物過(guò)程相關(guān),顯著富集在植物激素信號(hào)轉(zhuǎn)導(dǎo)、磷酸戊糖途徑、苯丙烷生物合成和-亞麻酸合成等途徑中。蘋果葉片外植體在外源激素的誘導(dǎo)下,通過(guò)對(duì)大量植物再生相關(guān)基因的轉(zhuǎn)錄激活,對(duì)植物體內(nèi)多種功能和代謝途徑進(jìn)行調(diào)節(jié),從而使離體葉片通過(guò)愈傷組織分化產(chǎn)生不定芽,內(nèi)源IAA和CTK的相互作用在這個(gè)過(guò)程中發(fā)揮至關(guān)重要的調(diào)控作用。
[1] 叢佩華, 張彩霞, 韓曉蕾, 田義, 張利義, 李武興. 我國(guó)蘋果育種研究現(xiàn)狀及展望. 中國(guó)果樹(shù), 2018(6): 1-5.
CONG P H, ZHANG C X, HAN X L, TIAN Y, ZHANG L Y, LI W X. Current research situation and prospect of apple breeding in China. China Fruits, 2018(6): 1-5. (in Chinese)
[2] VELASCO R, ZHARKIKH A, AFFOURTIT J, DHINGRA A, CESTARO A, KALYANARAMAN A, FONTANA P, BHATNAGAR S K, TROGGIO M, PRUSS D, SALVI S, PINDO M, BALDI P, CASTELLETTI S, CAVAIUOLO M, COPPOLA G, COSTA F, COVA V, Dal RI A, GOREMYKIN V,. The genome of the domesticated apple (×Borkh.). Nature Genetics, 2010, 42(10): 833-839.
[3] 常源升, 程來(lái)亮, 王海波, 何平, 李慧峰, 李林光. 蘋果分子標(biāo)記及輔助育種研究進(jìn)展. 園藝學(xué)報(bào), 2017, 44(9): 1658-1680.
CHANG Y S, CHENG L L, WANG H B, HE P, LI H F, LI L G. Review of molecular marker and marker assisted breeding of apple. Acta Horticulturae Sinica, 2017, 44(9): 1658-1680. (in Chinese)
[4] JAMES D J, PASSEY A J, BARBARA D J, BEVAN M. Genetic transformation of apple (Mill.) using a disarmed Ti-binary vector. Plant Cell Reports, 1989, 7(8): 658-661.
[5] EUN S S, KWAN J S, SUNG J, CHANG Y Y, ILL M C. Silver nitrate and aminoethoxyvinylglycine affect Agrobacterium -mediated apple transformation. Plant Growth Regulation, 2005, 45(1): 75-82.
[6] DAI H Y, LI W R, HAN G F, YANG Y, MA Y E, LI H, ZHANG Z H. Development of a seedling clone with high regeneration capacity and susceptibility toin apple. Scientia Horticulturae, 2013, 164: 202-208.
[7] VIDAL N, MALLóN R, VALLADARES S, MEIJOMíN A M, VIEITEZ A M. Regeneration of transgenic plants by- mediated transformation of somatic embryos of juvenile and mature. Plant Cell Reports, 2010, 29(12): 1411-1422.
[8] RICHARD L B, RALPH S, DELORES L. Adventitious shoot regeneration of pear (spp.) genotypes. Plant Cell, Tissue and Organ Culture (PCTOC), 2012, 108(2): 229-236.
[9] 趙政陽(yáng), 付潤(rùn)民, 稅守岐, 張秀琴, 黃英. 蘋果試管苗葉片再生植株研究. 陜西農(nóng)業(yè)科學(xué), 1992(6): 18-19.
ZHAO Z Y, FU R M, SHUI S Q, ZHANG X Q, HUANG Y. Study on the regeneration of apple plantlets from the leaves. Shaanxi Journal of Agricultural Sciences, 1992(6): 18-19. (in Chinese)
[10] IIZASA S, IIZASA E, WATANABE K, NAGANO Y. Transcriptome analysis reveals key roles of AtLBR-2 in LPS-induced defense responses in plants. BMC Genomics, 2017, 18(1): 995.
[11] 向亞男, 黃蕊蕊, 顧婷婷, 甘立軍. 基于RNA-Seq的擬南芥不定芽再生過(guò)程的基因表達(dá)譜分析. 南京農(nóng)業(yè)大學(xué)學(xué)報(bào), 2018, 41(2): 308-320.
XIANG Y N, HUANG R R, GU T T, GAN L J. Analysis of RNA- Seq-based expression profiles during adventitious shoot regeneration in. Journal of Nanjing Agricultural University, 2018, 41(2): 308-320. (in Chinese)
[12] CHE P, LALL S, NETTLETON D, HOWELL S H. Gene expression programs during shoot, root, and callus development intissue culture. Plant Physiology, 2006, 141(2): 620-637.
[13] MAYER K F X, SCHOOF H, HAECKER A, LENHARD M, JüRGENS G, LAUX T. Role of WUSCHEL in regulating stem cell fate in theshoot meristem. Cell, 1998, 95(6): 805-815.
[14] ZHANG T Q, LIAN H, TANG H B, DOLEZAL K, ZHOU C M, YU S, CHEN J H, CHEN Q, LIU H T, LJUNG K, WANG J W. An intrinsic MicroRNA timer regulates progressive decline in shoot regenerative capacity in plants. The Plant Cell, 2015, 27(2): 349-360.
[15] HIROYA I, KAORU S, PAUL T T, HARUKA T, SATOSHI K, YAYOI I, TAKUYA S, TAKU S, MITSUHIRO A, TAKAMASA S, SOICHI I, KENGO M, MOTOAKI S, TETSUJI K, ELLIOT M M, SACHIHIRO M. Primed histone demethylation regulates shoot regenerative competency. Nature Communications, 2019, 10(1): 1786.
[16] LI H, DURBIN R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics, 2009, 25(14): 1754-1760.
[17] OH Y, DONOFRIO N, PAN H Q, COUGHLAN S, BROWN D E, MENG S W, MITCHELL T, DEAN R A. Transcriptome analysis reveals new insight into appressorium formation and function in the rice blast fungus. Genome Biology, 2008, 9(5): R85.
[18] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods (San Diego, Calif.), 2001, 25(4): 402-408.
[19] ATTA R, LAURENS L, BOUCHERON-DUBUISSON E, GUIVARC'H A, CARNERO E, GIRAUDAT-PAUTOT V, RECH P, CHRIQUI D. Pluripotency ofxylem pericycle underlies shoot regeneration from root and hypocotyl explants grown. The Plant Journal, 2009, 57(4): 626-644.
[20] SUGIMOTO K, JIAO Y L, MEYEROWITZ E M.regeneration from multiple tissues occurs via a root development pathway. Developmental Cell, 2010, 18(3): 463-471.
[21] DUCLERCQ J, SANGWAN-NORREEL B, CATTEROU M, SANGWAN R S. De novo shoot organogenesis: from art to science. Trends in Plant Science, 2011, 16(11): 597-606.
[22] ZHANG T Q, LIAN H, ZHOU C M, XU L, JIAO Y L, WANG J W. A two-step model for de novo activation of WUSCHEL during plant shoot regeneration. The Plant Cell, 2017, 29(5): 1073-1087.
[23] GRAFI G, BEN-MEIR H, AVIVI Y, MOSHE M Y, DAHAN Y, ZEMACH A. Histone methylation controls telomerase-independent telomere lengthening in cells undergoing dedifferentiation. Developmental Biology, 2007, 306(2): 838-846.
[24] WILLIAMS A C, FORD W C L. Functional significance of the pentose phosphate pathway and glutathione reductase in the antioxidant defenses of human sperm. Biology of Reproduction, 2004, 71(4): 1309-1316.
[25] FAJKUS J, FULNE?KOVá J, HULáNOVá M, BERKOVá K, ?íHA K, MATYá?EK R. Plant cells express telomerase activity upon transfer to callus culture, without extensively changing telomere lengths. Molecular and General Genetics MGG, 1998, 260(5): 470-474.
[26] BHATIA R, DALTON S, ROBERTS L A, MORON-GARCIA O M, IACONO R, KOSIK O, GALLAGHER J A, BOSCH M. Modified expression of ZmMYB167 inandleads to increased cell wall lignin and phenolic content. Scientific Reports, 2019, 9(1): 8800.
[27] TAKEDA Y, KOSHIBA T, TOBIMATSU Y, SUZUKI S, MURAKAMI S, YAMAMURA M, RAHMAN M M, TAKANO T, HATTORI T, SAKAMOTO M, UMEZAWA T. Regulation of CONIFERALDEHYDE 5-HYDROXYLASE expression to modulate cell wall lignin structure in rice. Planta, 2017, 246(2): 337-349.
[28] SHANG B, XU C, ZHANG X, CAO H, XIN W, HU Y. Very-long- chain fatty acids restrict regeneration capacity by confining pericycle competence for callus formation in. Proceedings of the National Academy of Sciences of the United States of America, 2016, 113(18): 5101-5106.
[29] 孫貝貝, 劉杰, 葛亞超, 盛李宏, 陳呂琴, 胡小梅, 楊仲南, 黃海, 徐麟. 植物再生的研究進(jìn)展. 科學(xué)通報(bào), 2016, 61(36): 3887-3902.
SUN B B, LIU J, GE Y C, SHENG L H, CHEN L Q, HU X M, YANG Z N, HUANG H, XU L. Recent progress on plant regeneration. Chinese Science Bulletin, 2016, 61(36): 3887-3902. (in Chinese)
[30] GAJ M D. Factors influencing somatic embryogenesis induction and plant regeneration with particular reference to(L.) heynh. Plant Growth Regulation, 2004, 43(1): 27-47.
[31] GRIENEISEN V A, XU J, MARéE A F M, HOGEWEG P, SCHERES B. Auxin transport is sufficient to generate a maximum and gradient guiding root growth. Nature, 2007, 449(7165): 1008-1013.
[32] GORDON S P, HEISLER M G, REDDY G V, OHNO C, DAS P, MEYEROWITZ E M. Pattern formation during de novo assembly of theshoot meristem. Development (Cambridge, England), 2007, 134(19): 3539-3548.
[33] ANGELA K S, SANG H L, JONATHAN P W, NATHALIE G, HIRONORI I, DIRK I, WENDY A P, ANGUS S M, PAUL J O, WILLIAM M G. The SAUR19 subfamily of SMALL AUXIN UP RNA genes promote cell expansion. The Plant Journal, 2012, 70(6): 978-990.
[34] BARTRINA I, OTTO E, STRNAD M, WERNER T, SCHMüLLING T. Cytokinin regulates the activity of reproductive meristems, flower organ size, ovule formation, and thus seed yield in. The Plant Cell, 2011, 23(1): 69-80.
[35] 許智宏, 張憲省, 蘇英華, 胡玉欣, 徐麟, 王佳偉. 植物細(xì)胞全能性和再生. 中國(guó)科學(xué)(生命科學(xué)), 2019, 49(10): 1282-1300.
XU Z H, ZHANG X S, SU Y H, HU Y X, XU L, WANG J W. Plant cell totipotency and regeneration. Science in China (Series C), 2019, 49(10): 1282-1300. (in Chinese)
[36] FAN M Z, XU C Y, XU K, HU Y X. LATERAL ORGAN BOUNDARIES DOMAIN transcription factors direct callus formation inregeneration. Cell Research, 2012, 22(7): 1169-1180.
[37] XU C Y, CAO H F, ZHANG Q Q, WANG H Z, XIN W, XU E J, ZHANG S Q, YU R X, YU D X, HU Y X. Control of auxin-induced callus formation by bZIP59–LBD complex in Arabidopsis regeneration. Nature Plants, 2018, 4(2): 108-115.
[38] LAUX T, MAYER K F, BERGER J, JüRGENS G. The WUSCHEL gene is required for shoot and floral meristem integrity in. Development (Cambridge, England), 1996, 122(1): 87-96.
[39] MENG W J, CHENG Z J, SANG Y L, ZHANG M M, RONG X F, WANG Z W, TANG Y Y, ZHANG X S. Type-BRESPONSE REGULATORs specify the shoot stem cell niche by dual regulation of WUSCHEL. The Plant Cell, 2017, 29(6): 1357-1372.
[40] HORSTMAN A, WILLEMSEN V, BOUTILIER K, HEIDSTRA R. AINTEGUMENTA-LIKE proteins: hubs in a plethora of networks. Trends in Plant Science, 2014, 19(3): 146-157.
[41] KAREEM A, DURGAPRASAD K, SUGIMOTO K, DU Y J, PULIANMACKAL A J, TRIVEDI Z B, ABHAYADEV P V, PINON V, MEYEROWITZ E M, SCHERES B, PRASAD K. PLETHORA genes control regeneration by a two-step mechanism. Current Biology, 2015, 25(8): 1017-1030.
[42] SHAFI A, GILL T, SREENIVASULU Y, KUMAR S, AHUJA P S, SINGH A K. Improved callus induction, shoot regeneration, and salt stress tolerance inoverexpressing superoxide dismutase from. Protoplasma, 2015, 252(1): 41-51.
[43] TANG W, HARRIS L C, OUTHAVONG V, NEWTON R J. Antioxidants enhanceplant regeneration by inhibiting the accumulation of peroxidase inpine (Mill.). Plant Cell Reports, 2004, 22(12): 871-877.
[44] SRIVASTAVA S, DWIVEDI U N. Plant regeneration from callus of Cuscuta reflexa-an angiospermic parasite- and modulation of catalase and peroxidase activity by salicylic acid and naphthalene acetic acid. Plant Physiology & Biochemistry, 2001, 39(6): 529-538.
[45] CHAI M L, JIA Y F, CHEN S, GAO Z S, WANG H F, LIU L L, WANG P J, HOU D Q. Callus induction, plant regeneration, and long-term maintenance of embryogenic cultures in[L.] Merr. Plant Cell, Tissue and Organ Culture (PCTOC), 2011, 104(2): 187-192.
[46] FLORES H E, GALSTON A W. Osmotic stress-induced polyamine accumulation in cereal leaves I. physiological parameters of the response. Plant Physiology, 1984, 75(1): 102-109.
[47] SHOEB F, YADAV J S, BAJAJ S, RAJAM M V. Polyamines as biomarkers for plant regeneration capacity: Improvement of regeneration by modulation of polyamine metabolism in different genotypes of indica rice. Plant Science, 2001, 160(6): 1229-1235.
[48] MUKHOPADHYAY A, CHOUDHURI M M, SEN K, GHOSH B. Changes in polyamines and related enzymes with loss of viability in rice seeds. Phytochemistry, 1983, 22(7): 1547-1551.
Identification and Analysis of Differentially Expressed Genes in Adventitious Shoot Regeneration in Leaves of Apple
LIU Kai, HE ShanShan, ZHANG CaiXia, Zhang LiYi, BIAN ShuXun, YUAN GaoPeng, LI WuXing, KANG LiQun, CONG PeiHua, HAN XiaoLei
Research Institute of Pomology, Chinese Academy of Agricultural Sciences/Key Laboratory of Horticultural Crop Germplasm Resources Utilization, Ministry of Agriculture and Rural Areas/National Apple Breeding Center, Xingcheng 125100, Liaoning
【】In this study, the differentially expressed genes (DEGs) in adventitious shoot regeneration of ‘GL-3’ apple leaves were screened. The potential mechanism of adventitious shoot regeneration of apple leaves was analyzed, which will contribute to develop an efficient genetic transformation system for apple. 【】The explants of ‘Gl-3’ apple were cultured on regeneration medium. Samples were taken for RNA extraction and construction of mRNA library at 3, 7, 14 and 21 d post culture, respectively, further sequenced on the Illumina Nova seq platform. On the basis of the Kyoto Encyclopedia of Gene and Genome (KEGG) and Gene ontology (GO), the terms and pathway enrichment were then analyzed using the Phyper function with R software. Gene annotation was performed by using BLAST software. The DEGs related to plant regeneration, such as hormones, enzymes, transcription factors (TFs) and polyamines were analyzed, the expression levels of DEGs were verified by qRT-PCR. 【】Compared with the control group, 5 250, 4 937, 6 852 and 6 493 DEGs were identified at 3, 7, 14 and 21 d post culture, respectively, and 3 027 DEGs were shared in all four points. GO functional enrichment analysis showed that the up-regulated DEGs shared in all four points were mainly related to oxidation reduction process, cell periphery, protein kinase activity and organic cyclic compound binding, while the down-regulated DEGs were mainly related to single organism metabolic process, calcium ion binding, photosynthetic membrane and thylakoid part. KEGG pathway enrichment analysis indicated that the up-regulated DEGs shared in all four points were significantly enriched in pentose phosphate pathway, plant hormone signal transduction, plant pathogen interaction and protein processing in endoplasmic reticulum, while the down-regulated DEGs were significantly enriched in alpha linolenic acid metabolism, phenylpropanoid biosynthesis, carbon metabolism and photosynthesis. In addition, the DEGs encoding transcription factors, enzymes, and components of hormone biosynthesis and signaling pathways were analyzed. The results of qRT-PCR showed that most of these DEGs were up-regulated, which was consistent with data of RNA-Seq. 【】Through the detection and comparative analysis of large-scale gene expression profiles in adventitious shoot of ‘GL-3’ apple leaves at different time points, a number of genes related to adventitious shoot regeneration of apple leaves were obtained, which could provide a basis for further study on the mechanism of apple leaves in vitro regeneration.
apple; leaves regeneration; RNA-Seq; differentially expressed genes; impact factors

10.3864/j.issn.0578-1752.2021.16.011
2020-09-19;
2021-01-07
中央級(jí)公益性科研院所基本科研業(yè)務(wù)費(fèi)專項(xiàng)(Y2019XK09)、國(guó)家現(xiàn)代農(nóng)業(yè)產(chǎn)業(yè)技術(shù)體系建設(shè)專項(xiàng)(CARS-27)、中國(guó)農(nóng)業(yè)科學(xué)院科技創(chuàng)新工程(CAAS-ASTIP-2016-RIP-02)
劉鍇,E-mail:liukai2429@163.com。通信作者韓曉蕾,E-mail:hanxiaolei@caas.cn
(責(zé)任編輯 趙伶俐)
猜你喜歡
音樂(lè)探索(2022年2期)2022-05-30 21:01:37
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國(guó)特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52
