魚藤酮處理人神經母細胞瘤細胞的氧化水平研究
2021-08-25 02:04:10張萍邵靚
中國當代醫藥 2021年20期
張 萍 邵 靚
1.江西省人民醫院神經內科,江西南昌 330000;2.江西省人民醫院心血管內科,江西南昌 330000
多巴胺能神經元損傷和凋亡導致多巴胺分泌減少,是帕金森病的直接原因[1]。帕金森病多巴胺能神經元損傷及減少的原因和保護策略是目前帕金森病研究的重點及熱點。多種因素均可引起多巴胺能神經元損傷,其中環境因素所導致的帕金森病日益增多,引起越來越多的重視[2]。如何減輕環境所致的多巴胺能神經元損傷,減少有機農藥相關的帕金森病的發生具有現實意義。本研究從細胞氧化應激與帕金森病多巴胺能神經元減少的機制入手,從細胞氧化應激、細胞內抗氧化物及氧化物的水平干預等方面探討魚藤酮與多巴胺能神經元損傷的相關關系,可能為臨床帕金森病的治療提供切實有效的依據。
1 材料與方法
1.1 一般資料
1.1.1 實驗細胞 人神經母細胞瘤細胞(human neuroblastoma cells,SH-SY5Y)細胞購于武漢大學中國典型培養物保藏中心。
1.1.2 儀器與試劑 本研究采用的主要儀器與試劑見表1。

表1 儀器與試劑
1.2 細胞培養
在37°C,5% CO2恒定條件下,用含1%抗生素(青霉素和鏈霉素)的10%胎牛血清(美國Gibco)培養SH-SY5Y 細胞2 d。
1.3 實驗分組
根據細胞不同處理方式,將細胞分為正常組和魚藤酮組:正常組中1 mL 培養基加入1 μL 二甲基亞砜(dimethyl sulfoxide,DMSO)溶液;魚藤酮組中使用DMSO 稀釋10 mmol/L 魚藤酮溶液為不同濃度(25、50、100、200、500、1000、2000 nM)(nM:nmol/L)進行干預。
1.4 MTT 檢測
3-(4,5-二甲基噻唑-2)-2,5-二苯基四氮唑溴鹽(MTT),能與活性較強的細胞線粒體中的琥珀酸脫氫酶反應,被還原為不溶于水的藍紫色結晶并沉積在細胞中。使用酶標儀比色對比,在490 nm 波長處測定吸光值,可以間接反應存活細胞數量。
1.5 光鏡觀察細胞形態變化
活性較好的細胞呈橢圓形,突起較多;活性較差或死亡的細胞,突起消失變圓。
1.6 細胞內氧化應激水平檢測
利用流式細胞術,在37°C 下保持20 min,使用2′-7′-二氯二氫熒光素二乙酸酯(DCFH-DA)染色正常組和魚藤酮組細胞。其激發波長485 nm,發射波長530 nm。硫代巴比妥酸活性物質(thiobarbituric acid reactive substances,TBARS)和超氧化物根據實驗試劑盒步驟評估超氧化物歧化酶(superoxide dismutase,SOD)水平。
1.7 統計學方法
采用SPSS 13.0 統計學軟件進行數據分析,計量資料用均數±標準差(±s)表示,兩組間比較采用t檢驗;計數資料用率表示,兩組間比較采用χ2檢驗,以P<0.05 為差異有統計學意義。
2 結果
2.1 不同濃度魚藤酮處理SH-SY5Y 細胞存活率的比較
分別使用25、50、100、200、500、1000、2000 nM 魚藤酮干預SH-SY5Y 細胞后,以正常組細胞活性為100%,其他各組細胞與正常組比較,活性分別為(98.19±3.43)%、(95.79±3.95)%、(92.45±4.76)%、(86.63±3.89)%、(78.02±4.97)%、(62.50±5.21)%、(49.37±5.69)%。當魚藤酮濃度達到200 nM 及以上時,細胞活性明顯下降(P<0.05),與正常組比較,差異均有統計學意義(P<0.05)(圖1)。
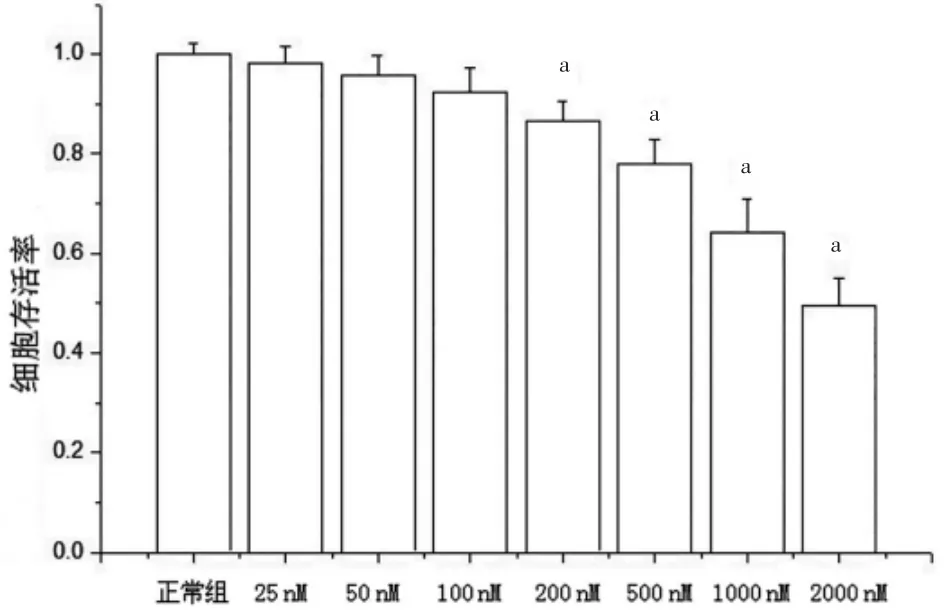
圖1 不同濃度魚藤酮處理SH-SY5Y 細胞存活率的比較
2.2 不同魚藤酮處理的SH-SY5Y 細胞光鏡結果
分別使用25、50、100、200、500、1000、2000 nM 魚藤酮干預SH-SY5Y 細胞后,光鏡下可見從200 nM開始細胞生長受限,隨著濃度的改變,細胞形態學及細胞數量減少更為明顯,2000 nM 的魚藤酮加入培養基后細胞幾乎停止生長,突起消失變圓,細胞生長與魚藤酮濃度呈濃度依賴的負相關(圖2)。
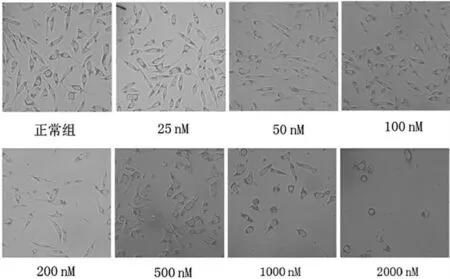
圖2 不同魚藤酮處理的SH-SY5Y 細胞光鏡結果(×400 倍)
2.3 魚藤酮干預后SH-SY5Y 細胞氧化應激水平的改變
選取500 nM 魚藤酮處理的SH-SY5Y 細胞24 h后,利用流式細胞術測定細胞內氧化簇(reactive oxygen species,ROS)水平,結果顯示,魚藤酮組細胞內ROS 水平高于正常組,差異有統計學意義(P<0.05)(圖3)。通過酶聯反應比色檢測發現,500 nM 魚藤酮處理后細胞內SOD 表達水平低于正常組,差異有統計學意義(P<0.05);細胞內脂質過氧化表達水平高于正常組,差異有統計學意義(P<0.05)(圖4)。

圖3 500 nM 魚藤酮處理細胞24 h 細胞ROS 水平
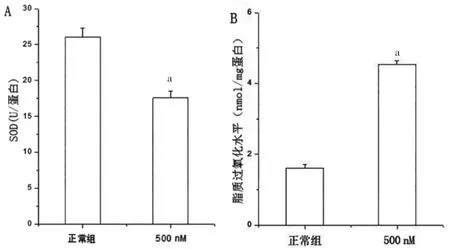
圖4 500 nM 魚藤酮處理的SH-SY5Y 細胞24 h 細胞SOD 水平及脂質過氧化水平
3 討論
帕金森病又稱震顫麻痹,是最常見的神經退行性疾病之一,主要病理表現為Lewy 體形成,中腦多巴胺能神經元變性死亡,紋狀體多巴胺含量顯著減少[3]。研究表明氧化應激是帕金森病發病機制中最常見的原因之一[4]。氧化應激通常指抗氧化物和氧化物間的失衡而導致的細胞損傷,包括抗氧化物如過氧化氫酶、超氧化物歧化酶、過氧化物還原酶家族、谷胱甘肽過氧化物酶等,以及氧化物如丙二醛、硫代巴比妥酸反應物質、丙烯醛等[5]。ROS 可以攻擊核酸導致DNA 單鏈和雙鏈斷裂,DNA 蛋白交聯、嘌呤和嘧啶堿基改變以及導致蛋白質編碼RNAs 和非編碼RNAs 的氧化修飾,可以攻擊蛋白質的骨干和側鏈,從而導致蛋白質的錯誤折疊和聚集[6]。同時,ROS 系統具有多種生物學作用,參與了激酶級聯反應的活化,鈣動員和信號傳導、微調控氧化還原敏感性基因的表達、在神經干細胞的分化以及神經發生中起作用[7-13]。在帕金森病患者的血液中還原型谷胱甘肽顯著高于正常組,而其它抗氧化因子濃度和過氧化氫酶的活性均有下降[14]。
長時間接觸農藥證實與神經系統變性疾病相關,這些農藥包括百草枯、代森錳、艾氏劑、狄氏劑、異狄氏劑、甲氰菊脂以及有機磷等。這些農藥均可誘導細胞氧化應激。已被確定與帕金森病發病相關的環境毒素1-甲基-4-苯基-1,2,3,6-四氫吡啶就可通過增加多巴胺能神經元內的氧化應激[15]。魚藤酮是一種線粒體呼吸鏈復合體Ⅰ的抑制劑,能夠通過抑制線粒體復合物Ⅰ,產生能量危機,而增加ROS 和OS 的產生,從而抑制線粒體呼吸鏈電子傳遞作用,造成跨膜電位差改變及線粒體膜通透性增強,導致細胞線粒體功能喪失,氧化應激激活,細胞凋亡壞死,最終造成在黑質致密部多巴胺能神經元的損失[16]。本課題組既往研究也顯示觸發氧化應激可以使暴露于氧化劑或淀粉樣蛋白-B 的肽(抗體)誘導的半胱氨酸殘基有利于分子間二硫鍵和隨后形成細胞質聚集體的不可逆氧化,這種不溶性蛋白質可能最終促進細胞死亡[17]。本研究通過魚藤酮處理多巴胺能神經母細胞系SH-SY5Y 細胞,證明其結果能造成氧化應激,這一定程度上可以模仿農藥等毒物造成帕金森病的機制。
許多抗氧化劑的研究也證實對帕金森病有神經保護作用。如銀杏葉提取物可對抗MPP+的神經毒性作用[18],另一項研究表明銀杏葉提取物對MPTP 引起的氧化應激有保護作用[19]。促甲狀腺激素釋放激素因其抗氧化應激、抗谷氨酸鹽的毒性作用、抗caspase誘導的細胞死亡、阻止DNA 片段化和抗炎作用而被研究于神經系統變性疾病如阿爾茨海默病與帕金森病[20]。既往本課題組研究也證實,通過敲低GAPDH 或者給予銀杏葉提取物可以抑制氧化應激水平,從而改善魚藤酮誘導的多巴胺能神經細胞死亡[21]。
綜上所述,本研究通過魚藤酮處理多巴胺能神經細胞,結果造成細胞內氧化應激,細胞內ROS 水平升高,脂質過氧化增強,SOD 水平下降,而抑制魚藤酮通過氧化應激途徑誘導的多巴胺能神經元損傷是治療帕金森病的潛在策略。本研究僅限于體外細胞研究,有待進一步進行動物研究和人群研究證實降低氧化應激水平治療帕金森病的有效性和安全性。
猜你喜歡
保健醫苑(2022年6期)2022-07-08 01:25:28
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
人大建設(2019年12期)2019-05-21 02:55:32
活力(2019年22期)2019-03-16 12:47:04
西南軍醫(2016年6期)2016-01-23 02:21:19
醫學研究雜志(2015年5期)2015-06-10 06:43:26
西南軍醫(2015年2期)2015-01-22 09:09:37
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
食品科學(2013年15期)2013-03-11 18:25:48
