芽孢桿菌有氧發酵豆粕替代魚粉對大口黑鱸幼魚生長性能、血清生化指標及腸組織結構的影響*
2021-08-25 07:04:14鐘國防田鑫鑫謝春元黃旭雄陳乃松朱站英
漁業科學進展 2021年5期
鐘國防 田鑫鑫 謝春元 黃旭雄,2,3 陳乃松,2,3 黃 衛 朱站英
(1. 上海海洋大學水產科學國家級實驗教學示范中心 上海 201306;2. 上海海洋大學農業農村部魚類營養與環境生態研究中心 上海 201306;3. 上海海洋大學農業農村部淡水水產種質資源重點實驗室 上海 201306;4. 上海希杰商貿有限公司 上海 201228;5. 湖州海皇生物科技有限公司 浙江 湖州 313105)
魚粉由于蛋白含量高、適口性好、氨基酸平衡,鈣、磷、碘、硒等含量高以及維生素種類較多等特點,是水產飼料中不可缺少的優質蛋白源(姚俊鵬等,2015),但隨著世界漁業養殖規模的迅速發展以及漁獲量的減少,導致魚粉的價格持續走高,養殖成本顯著提高,因此,魚粉替代蛋白源的研究一直為飼料研究的熱點(饒遠等, 2019; 胡鵬莉等, 2019; Zhonget al,2011)。豆粕的蛋白質含量高,氨基酸豐富,且有異黃酮等活性物質(Kishidaet al, 2000),是較為重要的魚粉蛋白替代原料。但普通豆粕有多種抗營養因子,過量使用不僅會降低水產動物對營養物質的吸收和利用,而且會對水產動物的消化系統和免疫系統造成損傷(Dunsfordet al, 1989),導致其在水產飼料中的應用比例受到限制。研究表明,豆粕經過微生物發酵得到的發酵豆粕(fermented soybean meal, FSBM)可有效降解其抗營養因子,并將大分子的大豆蛋白降解為小分子的蛋白、多肽及小肽等,且發酵過程中產生的益生菌、寡糖、乳酸、微生物和未知生長因子等活性物質能有效提高營養物質的吸收率(Honget al, 2004;李寶圣等, 2016; 張成等, 2018)。發酵豆粕產品開發,挖掘了豆粕蛋白在水產飼料中的應用潛力。劉興旺等(2010)發現,卵形鯧鲹(Trachinotus ovatus)飼料中發酵豆粕替代魚粉的水平(60.8%)顯著高于普通豆粕(45.1%)。黃雄斌等(2010)發現,對方正鯽(Carassius auratus gibelio),發酵豆粕能替代50%魚粉,而普通豆粕只能替代25%的魚粉。適量的發酵豆粕替代魚粉對魚蝦的生長無不良影響(呂云云等, 2016; 劉韜等,2018; 楊景豐等, 2018),但這些研究所用的發酵豆粕主要是以乳酸菌(Lactobacillus)為主的厭氧發酵產品。
發酵工藝對發酵豆粕產品的品質有重要影響(吳明海等, 2018)。芽孢桿菌(Bacillus)有氧發酵工藝由于發酵條件相對苛刻,特別是發酵過程中引發的產物黏稠,很難持續營造有氧環境,能全程有氧發酵的生產廠家很少。傳統的以厭氧乳酸菌為主的發酵工藝對豆粕抗營養因子特別是大分子蛋白(主要是抗原蛋白)的去除能力有限,僅能去掉14%~17.88%的大分子蛋白(表1),而本研究所用的以芽孢桿菌有氧發酵后的產品,分子質量為10~30 kDa 的小分子蛋白含量達26.38%,而對大分子蛋白的去除率達54.15%,能有效提高大分子蛋白的降解能力(表1),且發酵產品富含芽孢桿菌有益菌,產品質量優良。目前,在水產上的應用比較有限, 僅見虹鱒(Oncorhynchus mykiss)(Moniruzzamanet al, 2018)、凡納濱對蝦(Litopenaeus vannamei) (van Nguyenet al, 2018)和巖魚(Sebastes schlegeli) (Leeet al, 2016)等幾個品種,芽孢桿菌有氧發酵豆粕在大口黑鱸(Micropterus salmoides)上的應用研究還未見報道。
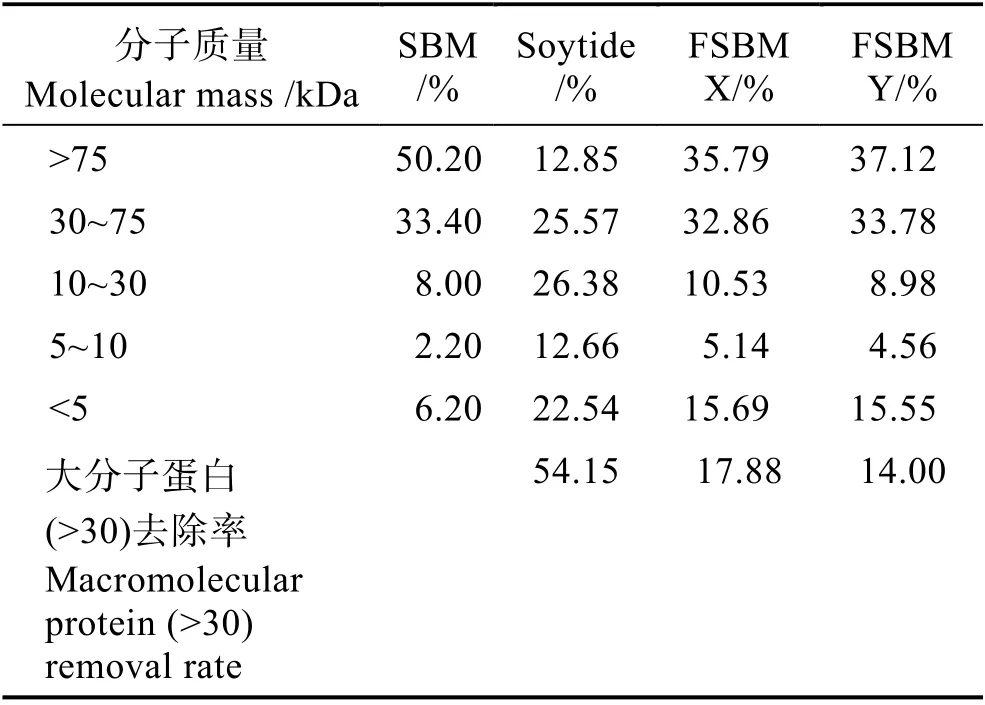
表1 乳酸菌厭氧發酵與芽孢桿菌有氧發酵對豆粕蛋白質的降解差異Tab.1 The difference of protein degradation on lactic acid bacteria fermented soybean meal and Bacillus aerobic fermentation of soybean meal
大口黑鱸又名加州鱸魚,是一種典型的肉食性魚類,其對飼料中蛋白質的營養需求較高(大于37%)(李二超等, 2011)。2018 年,我國大口黑鱸的產量達43 萬t,其中,全程投喂配合飼料養殖的比例超過50%,且大口黑鱸配合飼料中魚粉含量較高,一般為50%~55%,少數產品甚至達到60%。因此,研究大口黑鱸飼料中魚粉的替代具有極其重要的意義。本實驗研究芽孢桿菌有氧發酵豆粕替代魚粉對大口黑鱸生長性能、血清生化指標及腸道組織結構的影響,旨在為開發大口黑鱸高效、綠色環保、功能性飼料提供理論支持。
1 材料與方法
1.1 實驗設計及實驗飼料
根據大口黑鱸的營養需求,配制魚粉含量為50%的基礎飼料(FM),用芽孢桿菌有氧發酵豆粕分別替代FM 組10%(FSBM10)、20%(FSBM20)、30%(FSBM30)和40%(FSBM40)的魚粉,配制5 種等氮、等能的實驗飼料(表2)。芽孢桿菌有氧發酵豆粕(粗蛋白56.9%、粗脂肪2%)由上海希杰商貿有限公司提供,其他原料由湖州海皇生物科技有限公司提供。原料粉碎后,過60 目篩,按飼料配方稱重,逐級混勻,加入適量的水,混勻后過10 目篩,用制粒機制粒,烘箱熟化后于60℃烘干18 h,包裝后置于-20℃冰箱保存備用。
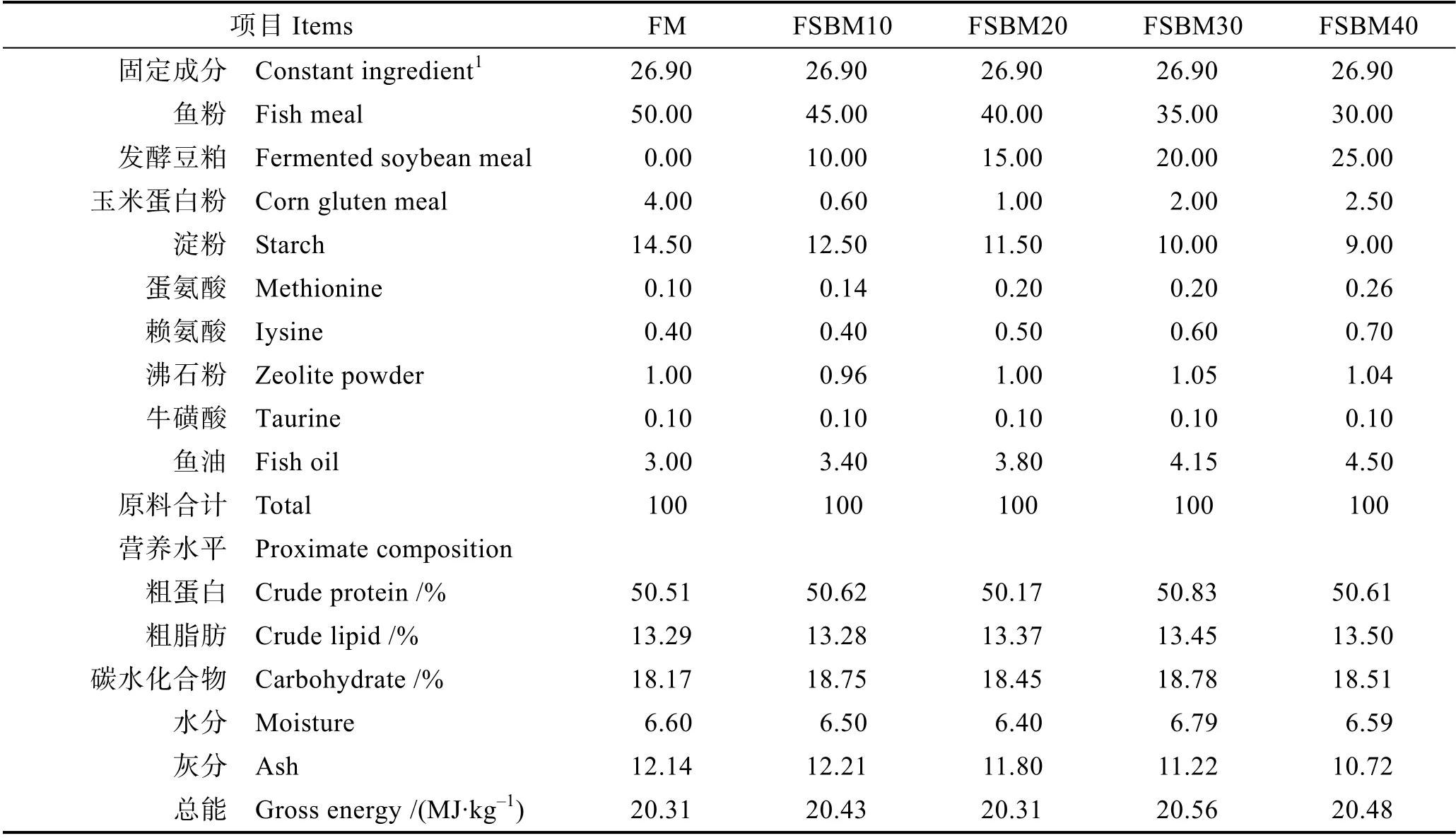
表2 飼料組成和營養水平(%風干樣品)Tab.2 Composition and nutrient levels of diets (% air dry basis)
1.2 養殖管理
實驗在上海海洋大學濱海基地室內養殖池網箱(1.0 m×1.0 m×1.2 m)中進行。大口黑鱸幼魚苗購自上海秦皇山漁業有限公司,幼魚運到基地后先暫養14 d 并用基礎飼料進行馴化。暫養結束挑選形態正常、體質健康、規格均勻的450 尾魚隨機分到15 個網箱中。實驗分5 組,每組3 個重復,每個重復30 尾魚。每天飽食投喂2 次(07:00 和18:00),投飼后1 h 收集殘餌。每周測定水質指標,養殖期間水溫為(29.0±3.5)℃,pH 值為8.5±0.5,溶解氧(DO)為4.0~5.0 mg/L,氨氮為0~0.03 mg/L。養殖實驗持續75 d。
1.3 樣品采集
養殖實驗結束后,停食24 h,每平行隨機取6 尾魚,分別測量魚體質量與體長,冰上分離內臟與肝臟并稱重記錄,用于計算體質指標。尾部抽血,放入3 個2 mL 離心管中,離心(10 min, 8000 r/min, 4℃)分離血清備用。
1.4 測定指標和方法
實驗結束后撈出網箱中魚,計數稱重,并計算成活率、增重率和飼料系數。計算方法如下:
存活率(survival rate, SR, %)=100×實驗結束時尾數/實驗開始時尾數;
增重率(weight gain rate, WGR, g)=[(末體重-初始體重)/初始體重]×100%;
飼料效率(feed efficiency, FE)=(末體重-初始體重+死亡體重)/(投料量-殘餌量);
飼料營養成分測定:水分測定采用105℃常壓干燥法,粗灰分測定采用550℃灼燒法,粗蛋白質測定采用杜馬斯燃燒法(Etheridgeet al, 1998),粗脂肪測定采用氯仿-甲醇法(黃凱等, 2011)。
血清中堿性磷酸酶(ALP)、白蛋白、總蛋白、球蛋白、谷丙轉氨酶(ALT)和谷草轉氨酶(AST)采用江蘇南京建成生物工程研究所購試劑盒測定。
1.5 腸組織切片制作及觀察
養殖實驗結束后,饑餓24 h,每個重復隨機取3 尾魚,冰上分離并取前腸、中腸和后腸組織各2~3 cm,生理鹽水沖洗后,用濾紙擦干表面水分后放入Bouin 氏液中,24 h 內換用70%的酒精保存。制作切片時,組織樣本乙醇梯度脫水,二甲苯透明處理,石蠟包埋,切片(5 μm),隨后進行展片、烤片、H&E 染色和封片等系列工作(夏青等, 2015; 陳曉瑛等, 2018)。
1.6 數據處理
實驗數據采用SPSS 17.0 軟件進行單因子方差分析(one-way ANOVA, LSD),若存在顯著差異,則進行Duncan 多重比較。所有統計值均以平均值±標準差(Mean±SD)表示,顯著水平定為P<0.05。
2 結果
2.1 芽孢桿菌有氧發酵豆粕替代魚粉對大口黑鱸幼魚生長性能的影響
由表3 可知,適量的芽孢桿菌有氧發酵豆粕能顯著提高大口黑鱸幼魚的增重率(P<0.05),且隨著發酵豆粕替代魚粉用量的增加呈先上升后下降的趨勢,10%組與對照組無顯著差異,替代量20%組的生長性能最好并顯著優于其他各實驗組(P<0.05),但當替代量達到40%時,增重率與對照組無顯著差異(P>0.05);各實驗組的飼料效率并無顯著差異(P>0.05)。各實驗組間存活率也無顯著差異(P>0.05)。
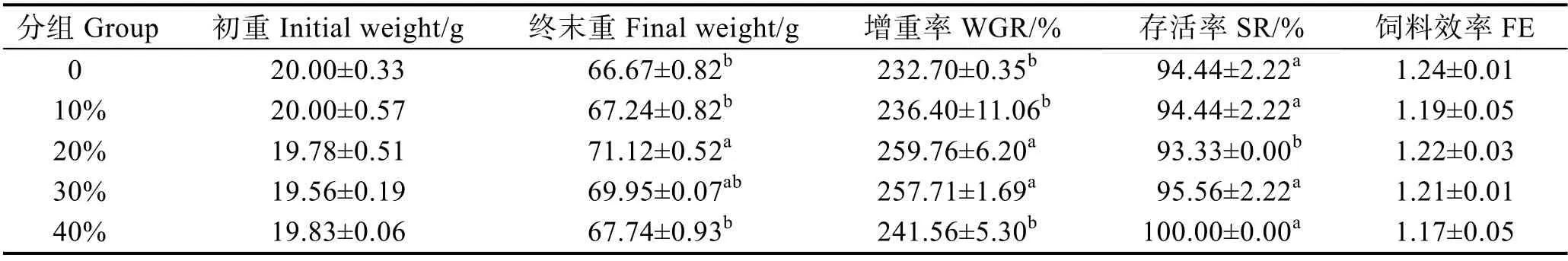
表3 芽孢桿菌有氧發酵豆粕替代魚粉對大口黑鱸生長性能的影響Tab.3 Effects of replacement of FM by FSBM on growth indices of M. salmoides
2.2 芽孢桿菌有氧發酵豆粕替代魚粉對大口黑鱸血清生化指標的影響
由表4 可知,大口黑鱸血液中的白蛋白含量隨替代量的增加先上升后下降,在20%組出現最大值,但各組間無顯著差異(P>0.05);實驗組總蛋白和球蛋白的含量顯著高于對照組(P<0.05);谷丙轉氨酶活力隨魚粉替代量的增加而顯著升高,10%組與對照組無顯著差異(P>0.05),其他各替代組顯著高于對照組(P<0.05)。10%組的谷草轉氨酶活力與對照組無顯著差異,但其他各替代組顯著高于對照組(P<0.05)。
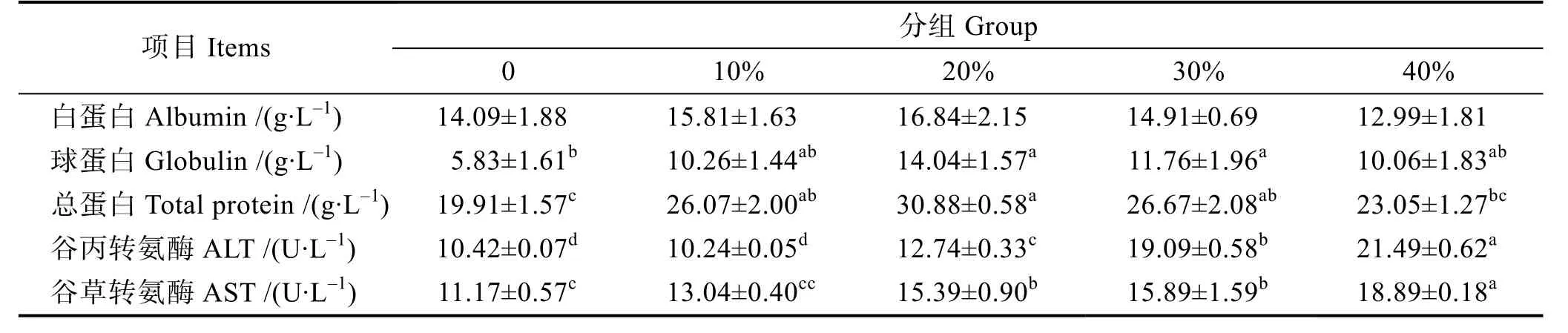
表4 芽孢桿菌有氧發酵豆粕對大口黑鱸血清生化指標的影響Tab.4 Effects of replacement of FM by FSBM on biochemical indices in serum of M. salmoides
2.3 發酵豆粕替代魚粉對大口黑鱸形體指標的影響
由表5 可知,肥滿度隨著魚粉替代量的增加呈先上升后下降的趨勢,在20%替代量時出現最大值,但無顯著差異(P>0.05);肝體比隨著替代量的增加呈下降的趨勢,但各組之間無顯著差異(P>0.05);隨著替代量的增加,替代組臟體比出現下降趨勢,且當替代量超過30%時,顯著低于對照組(P<0.05)。
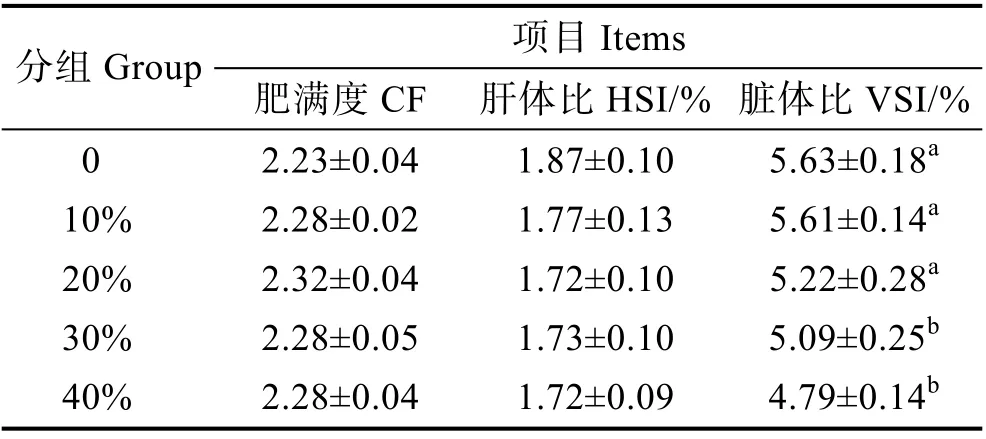
表5 芽孢桿菌有氧發酵豆粕替代魚粉對大口黑鱸形體指標的影響Tab.5 Effects of replacement of FM by FSBM on morphometry of M. salmoides
2.4 芽孢桿菌有氧發酵豆粕替代魚粉對大口黑鱸腸道組織結構的影響
腸道切片形態結構顯微觀察發現,前腸對照組大口鱸的腸絨毛結構完整,發育良好,杯狀細胞較多且排列密集有序,但40%組腸絨毛密度稍有降低,有絨毛脫落;后腸組織中觀察到上皮細胞和肌肉層的分離現象,40%組腸絨毛的完整性降低,腸絨毛減少,杯狀細胞數量減少,并且腸黏膜層與腸肌層之間的連接明顯變得疏松(圖1)。

圖1 芽孢桿菌有氧發酵豆粕替代魚粉對大口黑鱸腸道組織結構的影響Fig.1 Effects of replacement of FM by FSBM on intestinal tissue structure of M. salmoides
由表6 可知,隨著發酵豆粕替代的增加,前腸的肌層厚度呈顯著下降的趨勢(P<0.05),各實驗組前腸的絨毛長度和絨毛寬度均顯著大于對照組(P<0.05)。

表6 芽孢桿菌有氧發酵豆粕替代魚粉對大口黑鱸腸道組織結構的影響Tab.6 Effects of replacement of FM by FSBM on intestinal tissue structure of M. salmoides
中腸肌層厚度各實驗組顯著低于對照組(P<0.05),絨毛長度除10%組顯著大于對照組外,其他各實驗組顯著小于對照組(P<0.05),而絨毛寬度各組間無顯著差異(P>0.05)。
后腸肌層厚度20%和30%組顯著大于對照組,而其他實驗組顯著小于對照組(P<0.05),30%組絨毛長度顯著大于其他各組,20%和40%組絨毛長度顯著小于對照組(P<0.05),絨毛寬度20%和30%組顯著大于對照組(P<0.05)。
3 討論
3.1 芽孢桿菌有氧發酵豆粕對大口黑鱸幼魚生長性能的影響
豆粕替代魚粉受到限制主要是因為抗營養因子對魚生長的消極影響,而對豆粕抗營養因子的研究推動了發酵豆粕產品的快速發展。以乳酸菌厭氧發酵制得的發酵豆粕與普通豆粕相比,在魚類的生長性能、消化吸收及對魚類消化生理結構的影響方面都有積極效應。魚類飼料中不同工藝處理后的發酵豆粕能替代16%~40%的魚粉(Barneset al, 2015; Yaghoubiet al,2016),豆粕處理及發酵工藝的不同對抗營養因子的去除能力也不一樣,芽孢桿菌有氧發酵工藝對大分子蛋白的降解能力優于傳統的乳酸菌厭氧發酵(表1),對豆粕抗營養因子的處理能力有顯著提高。
發酵豆粕替代魚粉的潛力與發酵豆粕產品生產工藝及養殖對象的食性有密切關系。本實驗研究芽孢桿菌有氧發酵在大口黑鱸飼料中替代魚粉的潛力,養殖實驗結束后發現,20%~30%替代組均表現出較佳的生長性能。Azarm等(2014)研究發現,能用乳酸菌厭氧發酵豆粕替代 40%的魚粉飼喂黑鯛(Acanthopagrus schlegeli)。Barnes等(2015)發現,能用乳酸菌厭氧發酵豆粕替代 35%的魚粉飼喂虹鱒(Oncorhynchus mykiss)。Zhang等(2014)研究發現,能用γ射線照射后的豆粕替代 16%的魚粉飼喂日本花鱸(Lateolabrax japonicus)。Kokou等(2012)研究發現,能用生物處理豆粕替代近40%的魚粉飼喂金頭鯛(Sparus aurata)。芽孢桿菌有氧發酵豆粕替代魚粉的研究目前也有相關報道,如在虹鱒、凡納濱對蝦和巖魚(Moniruzzamanet al, 2018; van Nguyenet al, 2018; Leeet al, 2016 )替代魚粉的量分別為30%、25.36%和40%。
本研究20%~30%實驗組比對照組顯著提高了增重率,可能是由于發酵后大分子蛋白質被大量分解為多肽和小肽,提高了吸收效率。游離氨基酸的吸收是逆濃度梯度的轉運,要通過不同的Na+轉運系統才能完成(Matthews, 1980),且游離氨基酸在吸收時有相互競爭,需要消耗較多的能量。小肽的吸收不通過水解,而是通過特殊的轉運系統,完整地進入血液中(Jalkanenet al, 2004)。因此,小肽比游離氨基酸更有利于魚類吸收,能提高魚類高蛋白質的合成效率。此外,植物蛋白替代魚粉最大的不利因素是缺乏一些必需氨基酸 (Kaderet al, 2012; Zhanget al, 2014; Joetal, 2017),導致在吸收時會影響氨基酸在體內合成蛋白質的效率,而以小肽的形式直接吸收可能會避免這種缺陷,其具體的吸收合成過程還有待于進一步開展相關的生理實驗來證實。
3.2 芽孢桿菌有氧發酵豆粕對大口黑鱸幼魚血清生化指標的影響
魚類的血液指標能充分反映魚體的生理和健康狀態(Maita, 2007),生理或病理因素都會導致它們的變化。血清白蛋白主要反映機體營養能力,而球蛋白與免疫能力相關。實驗結果顯示,魚粉替代量在30%以內時,實驗組血清白蛋白略有升高,說明適宜的替代魚粉對機體的營養能力沒有影響。但替代組的球蛋白顯著高于對照組,表明替代后導致機體免疫系統活躍。
谷丙轉氨酶、谷草轉氨酶是機體分布最廣、活性最強的轉氨酶,通常作為評估肝臟健康狀況的指示酶(Krajnovic-Ozretic, 1991)。魚類與哺乳動物一樣,當組織或器官受到功能性損傷時,某些特有的細胞酶會釋放到血液里而被檢測出來(Boucket al, 1975)。正常情況下,轉氨酶主要存在于肝臟。肝功能障礙或肝細胞受損時,谷丙轉氨酶和谷草轉氨酶會釋放到血液中,引起血清轉氨酶活性的升高(Kaplan, 1979)。實驗組谷丙轉氨酶與谷草轉氨酶隨著替代量的增加而升高且均顯著高于對照組,表明大口黑鱸攝食替代魚粉飼料后,肝臟產生了應激。養殖魚蝦投喂植物蛋白含量高的飼料容易出現營養性脂肪肝,主要是因為植物蛋白含有大量的抗營養因子(吳莉芳等, 2006; 陳曉明等, 2015)。本實驗魚沒有發現有明顯的綠肝、花肝等病變,可能是因為相對于養殖生產實驗飼養時間較短,且芽孢桿菌有氧發酵后清除了大量的抗營養因子。但長期應激是否產生肝功能障礙等不良影響,還有待于進一步進行中試驗證。這也提示在實際生產時,芽孢桿菌有氧發酵豆粕替代魚粉的量要從生長、生理及病理等多個因素綜合評測。
3.3 芽孢桿菌有氧發酵豆粕對大口黑鱸幼魚形體指標的影響
本研究的形體指標中實驗組肝體比有明顯下降的趨勢,但與對照組無顯著差異,這與Kokou 等(2012)和Yaghoubi 等(2016)的研究結果相似,這可能是因為實驗時間較短所致(Qiuet al, 2018)。肝體比增加與許多因素有關,如必需氨基酸低也會導致養殖對象肝體比的增大(Espeet al, 2010)。而本研究中,肝體比沒有增加,說明發酵豆粕中豐富的小分子量蛋白質、小肽等可以彌補必需氨基酸少的缺陷。本研究20%和30%組的臟體比顯著低于對照組,這一結果要優于王新霞(2009)的研究結果,其研究表明,加州鱸飼料中添加10%以內的發酵豆粕,能顯著降低臟體比和肝體比,降低了脂肪在體內的沉積,從而降低脂肪肝的發生率,而本研究20%和30%組臟體比均為下降。這可能是因為本研究飼料的營養組成中小肽含量豐富,以小肽吸收能促進幼魚的快速生長,對能量的需求加大,促使飼料中的糖類更多地用于能量消耗,而轉化為脂肪儲存相對較少。這種推斷需要進一步研究糖的代謝來證實。同時,小肽能促進“脂質代謝”(Drevon, 2005),也會減少脂肪在內臟的沉積,致使臟體比下降。
3.4 芽孢桿菌有氧發酵豆粕對大口黑鱸幼魚腸道組織結構的影響
腸道絨毛高度、密度以及杯狀細胞的數量等組織形態學指標反映魚類腸道的發育和健康程度,通常作為評估魚類腸道的消化吸收能力的依據(聶國興等,2007; 王壇等, 2019)。植物蛋白替代魚粉會導致腸道組織結構發生變化。芽孢桿菌有氧發酵豆粕替代魚粉后,盡管適宜替代量(20%和30%替代)促進了大口黑鱸的生長性能,但對腸道組織形態學研究表明,40%的魚粉替代引起了前腸肌肉層厚度的下降,損害了小腸的完整性。鄭銀樺等(2015)用混合植物蛋白質替代50%魚粉后,引起了大口黑鱸腸道肌層變薄,絨毛完整度下降等不良影響,與本研究結果相似。本實驗切片觀察還發現,替代組前腸絨毛長度和寬度均顯著優于對照組,推測絨毛面積的增加促進了營養物質的吸收,從而促進了生長,這與Yamamoto 等(2010)在虹鱒中的研究結果相一致。趙柳蘭等(2018)研究認為,大口黑鱸前腸皺襞密集而細長,中性黏液細胞多,是大口黑鱸營養物質消化吸收的主要場所,肌肉層厚度下降會影響前腸蠕動的能力,對前腸物理性消化功能會產生不良影響,但絨毛面積增加而引起的吸收功能的提高,彌補了物理消化減弱這個缺陷,這也是20%和30%組生長性能優于對照組的一個極為重要的原因。同時,由于大口黑鱸為有胃魚,胃的蠕動減少了腸道蠕動的壓力,因此,營養物質經過胃和前腸充分的消化吸收作用以后,中后腸結構的變化對營養物質消化吸收影響就相對有限。杯狀細胞分泌黏液協助消化和保護胃腸道組織免受損傷(Cerezuelaet al, 2013;Khosraviet al, 2015)。本研究表明,魚粉替代對前腸杯狀細胞沒有不良影響,但替代量達到40%時,后腸杯狀細胞數量明顯減少。Bakke-McKellep 等(2007)研究發現,在豆粕型日糧中添加益生元和益生菌后,通過微生物的調控可以改變杯狀細胞的數量。飼料中的芽孢桿菌對虹鱒小腸的杯狀細胞的形成有促進作用(Ramoset al, 2017),本研究所用的發酵豆粕含有豐富的芽孢桿菌,這可能是導致前腸杯狀細胞不受影響的主要原因。與之相反,Khosravi 等(2015)研究認為,大豆濃縮蛋白會導致杯狀細胞數量的減少,可能是大豆濃縮蛋白對抗營養因子的去除能力有限所致。
4 結論
芽孢桿菌有氧發酵豆粕能顯著增加飼料小分子蛋白、肽類的含量,適量替代魚粉能促進大口黑鱸的生長,且能降低臟體比。但同時引起了轉氨酶活性的升高,增加了營養應激程度,長期飼喂會使生長受阻。替代量太高會引起腸組織結構的損傷,從而影響消化吸收。因此,在本研究中,綜合考慮生長、形體指標和消化道組織結構等多方面因素,大口黑鱸飼料中芽孢桿菌有氧發酵豆粕替代20%的魚粉能達到理想的效果。
