不同栽植年限小葉楊與胡楊雜交種(小×胡楊)光合響應特性比較
2021-08-16 01:50:36黃雅茹馬迎賓遲悅春趙英銘劉禹廷
西北林學院學報 2021年4期
關鍵詞:植物
黃雅茹,馬迎賓,遲悅春,趙英銘,褚 洋,孫 非,劉禹廷,張 帥
(1.內蒙古磴口荒漠生態系統國家定位觀測研究站/中國林業科學研究院 沙漠林業實驗中心,內蒙古 磴口 015200; 2.中國林業科學研究院 華北林業實驗中心,北京 102300)
光照對于植物生長發育具有非常重要的影響,如果植物長期生長在不同的光強下,即使是相同的植物,為了適應不同的光環境,其形態指標、生理指標也會發生相應的改變。植物生長發育、生物量的積累、有機物質的積累重要基礎是光合作用[1],光合作用隨著光強的變化而變化。光照強度增加,光合速率也增強,反之亦然,因此光照強度顯著影響植物的光合作用[2-3]。光合作用的生理過程非常復雜,植物自身生理特性和環境因素共同影響光合作用,光合作用的過程時刻受光照、溫度、水分等環境因子的影響,其中,光合作用的先決條件是光照[4]。測定光合作用后,通過公式計算求出光飽和速率(Amax)、光飽和點(LSP)、光補償點(LCP)、表觀量子效率(AQY)、暗呼吸效率(RD)等生理參數[5]。研究表示,通過上述生理參數有助于判定植物光合作用能力和光合作用效率及其受環境變化的影響程度[6-7]。在干旱區,植物的水分生理及光合生理特性與人們采取的植被管理措施密切相關,因此,植物的水分生理及光合特性直接決定了植物的生存和發展狀況[8]。了解荒漠植物在特定生境中的光合以及水分生理特性非常重要。
烏蘭布和沙漠綠洲是內蒙古河套地區防風固沙的重要屏障,目前,防護林體系主要的造林樹種有新疆楊(Populusalbavar.pyramidalis)、二白楊(Populusgansuensis)、小葉楊(Populussimonii)、毛白楊(Populustomentosa)等,但是,綠洲防護林的部分退化引發了一些環境問題及社會矛盾,因此,林業部門對防護林體系的更新與重建非常重視,尤其是樹種選擇問題,是目前急需解決的難題[9]。我國干旱沙漠地區喬木造林樹種非常少,是生態建設中亟待解決的重大技術問題;而用干旱沙漠區特有的樹種材料,通過雜交育種的方式,選育出適宜干旱荒漠區的高抗性新品種是我國的重要課題。小×胡楊(Populussimonii×P.euphratica)是小葉楊(Populussimonii)和胡楊 (Populuseuphratica)進行多次雜交試驗選育出的優良品種;小葉楊(♀)和胡楊(♂)有性雜交后,完全具有雙親融合性狀[10]。前人研究顯示,小×胡楊比小葉楊和胡楊生長迅速,耐鹽堿性強于小葉楊,比胡楊更容易進行繁殖,是干旱沙漠區防護林造林的新品種[10]。目前,有關小×胡楊的研究報道較少,主要包括小×胡楊幼苗抗旱性[10]、育種[10]、引種[10]、硬枝扦插技術[10]、無菌培養[11]方面的研究,而有關小×胡楊氣體交換參數對光強的響應方面的研究報道還很缺乏。鑒于此,本研究以烏蘭布和沙漠東北部不同栽植年限的小×胡楊為研究對象,測定氣體交換參數,分析不同光強對小×胡楊氣體交換參數的影響,闡明其對環境光合特性的適應和生理響應,以期為烏蘭布和沙漠東北部防護林體系的更新及造林樹種選擇提供一定的理論依據,為綠洲前沿生態環境的改善提供理論依據,為進一步評價小×胡楊新品種適應性提供基礎數據。
1 材料與方法
1.1 研究區概況
研究區地理坐標為39°40′-41°00′N,106°00′-107°20′E,地形起伏不大,沙丘高度≤10 m,新月形沙丘或圓錐形沙丘是研究區的主要沙丘類型,沙質主要以細沙為主。研究區屬于中溫帶半干旱大陸性氣候,主要特點是云少,降水少、氣候干燥,風沙活動比較多,風速較高。研究區多年平均降水量約140.3 mm(1954-2005年),沙漠晝夜溫差大,平均氣溫為6.8℃,年日照時間為3 229.9 h。研究區主風向以西風和西北風為主,11月至翌年5月為研究區的風沙季節。土壤類型主要以風沙土為主[12]。人工林主要有檸條錦雞兒(Caraganakorshinskii)、梭梭(Haloxylonammodendron)、花棒(Hedysarumscoparium)等,天然植被主要有油蒿(Artemisiaordosica)、白刺(Nitrariatangutorum)等,綠洲防護林體系主要栽植樹種是楊樹[13]。
1.2 試驗材料
試驗于2018年8月23日、24日進行,試驗材料是小×胡楊(Populussimonii×P.euphratica),是小葉楊(Populussimonii)和胡楊 (Populuseuphratica)進行多次雜交試驗選育出的優良品種,小葉楊(♀)和胡楊(♂)有性雜交后,完全具有雙親融合性狀。試驗以不同栽植年限的小×胡楊為研究對象,栽植立地條件一致,且栽植在同一林地。具體選擇1年、2年、3年生的小×胡楊,對不同栽植年限的小×胡楊的生長狀況進行測定,包括株高、胸徑、冠幅、枝下高。
1.3 研究方法
不同栽植年限的小×胡楊葉片光響應曲線采用Li-6400xt便攜式光合儀進行測定,在晴朗無云的天氣,采用LED紅藍光源,1年、2年、3年生小×胡楊各選擇3株為標準株,選擇向陽無遮光部位的葉片,每株在冠層上、中、下各選擇2片葉片,9:00-12:00進行測定。選取葉片為成熟且無病蟲害的,CO2濃度設置為400 μmol·m-2·s-1,光強設置為1 500、1 200、900、600、300、100、50、0 μmol·m-2·s-1,重復3次。光響應曲線模擬是用光合助手軟件(擬合植物葉片光合有效輻射(PAR)與凈光合速率(Pn)的關系)。計算出小×胡楊的光合生理參數(光飽和速率,光補償點及光飽和點,表觀量子效率等)。非直角模型理論公式[14]為:
(1)
式中,A為表觀量子效率,K為光響應曲線曲角,Q為光合有效輻射,Pnmax為凈光合速率,RD為暗呼吸速率。
1.4 數據處理
數據整理及繪圖使用Excel,采用函數STDEVP計算誤差值。采用SPSS17.0進行多重比較分析。
2 結果與分析
2.1 不同栽植年限小×胡楊生長狀況
3年生小×胡楊株高與胸徑最大,分別是1年生的1.89、1.68倍,不同栽植年限樹高與胸徑存在顯著差異(P<0.05),3年生小×胡楊冠幅也最大,與2年生差異不顯著(P>0.05),與1年生差異顯著(P<0.05)。3年生小×胡楊枝下高最大,與1年、2年生差異不顯著(P>0.05)。
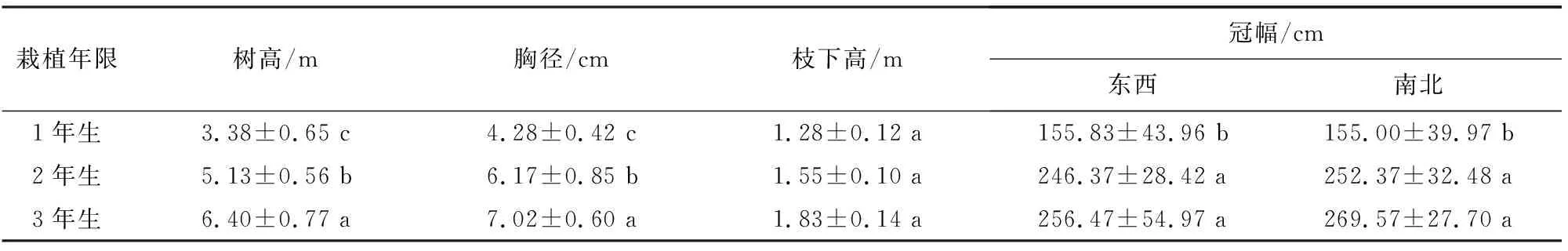
表1 不同栽植年限小×胡楊生長指標
2.2 不同栽植年限小×胡楊凈光合速率光響應特征
植物光合能力的強弱是由光合速率來決定的,在一定的環境條件下,植物葉片的最大光合能力通過最大凈光合速率來體現[14-15]。隨著光合有效輻射的增加,不同栽植年限小×胡楊變化趨勢一致。由圖1可以看出,當光合有效輻射在0~900 μmol·m-2·s-1時,不同栽植年限小×胡楊凈光合速率均隨著光合有效輻射(PAR)的增強而迅速增加,當光合有效輻射(PAR)>900 μmol·m-2·s-1時,不同栽植年限小×胡楊凈光合速率(Pn)隨光合有效輻射(PAR)的增強緩慢增加,光響應曲線變化較為平緩。當光合有效輻射(PAR)為1 500 μmol·m-2·s-1時,曲線處于緩慢上升趨勢,說明小×胡楊還沒有達到光飽和點。經SPSS17.0方差分析與多重比較分析,1年、2年、3年生小×胡楊之間的凈光合速率差異顯著(P<0.05)。不同光合有效輻射,凈光合速率值均表現為3年>2年>1年生。

圖1 小×胡楊凈光合速率光響應曲線
2.3 不同栽植年限小×胡楊光響應特征參數
光飽和速率(Amax)由高到低依次是3年>2年>1年生(表2)。光飽和點(LSP)用來表示植物利用光照強度的能力,光飽和點(LSP)值高,表明強光照射植物時,其生長發育不容易受到抑制,光補償點(LCP)反映了植物利用弱光的能力,其值越小,說明植物利用弱光的能力越強,3年生小×胡楊LSP最高,說明強光照射時3年生小×胡楊生長發育不易受到抑制。1年生小×胡楊LCP最低,說明1年生小×胡楊利用弱光的能力越強。不同栽植年限小×胡楊的表觀量子效率(AQY)從大到小依次是3年>2年>1年生。暗呼吸速率(RD)是指植物在沒有光照條件下的呼吸速率,1年生小×胡楊的暗呼吸速率最大,3年生小×胡楊的暗呼吸速率最小,3年生小×胡楊的光飽和速率(Amax)、光飽和點(LSP)、光補償點(LCP)、表觀量子效率(AQY)均最高,不同栽植年限小×胡楊光飽和速率(Amax)差異達到顯著(P<0.05),光飽和點(LSP)、光補償點(LCP)、暗呼吸速率(RD)不顯著(P>0.05),表觀量子效率(AQY)2年生小×胡楊與3年生小×胡楊之間差異不顯著(P>0.05),均與1年生小×胡楊差異顯著(P<0.05)。
2.4 不同栽植年限小×胡楊蒸騰速率光響應特征
植物水分運輸過程中,蒸騰作用起著主導作用。由圖2可知,隨著光合有效輻射的增加,不同栽植年限小×胡楊蒸騰速率緩慢增加,曲線趨勢變化較為平緩。經SPSS17.0方差分析與多重比較分析,1年、2年、3年生小×胡楊之間的凈光合速率光響應差異不顯著(P>0.05)。不同光合有效輻射,蒸騰速率值均表現為3年>2年>1年生。

圖2 小×胡楊蒸騰速率的光響應
2.5 不同栽植年限小×胡楊氣孔導度和胞間CO2濃度的光響應特征
由圖3可知,隨著光合有效輻射的增加,1年、2年、3年生小×胡楊氣孔導度(Gs)變化趨勢一致,均呈增加趨勢。不同光合有效輻射,氣孔導度(Gs)值均表現為2年>3年>1年生。當光合有效輻射<600 μmol·m-2·s-1,1年、2年、3年生小×胡楊氣孔導度(Gs)增加緩慢,當光合有效輻射>600 μmol·m-2·s-1,隨著光強的增強,1年、2年、3年生小×胡楊氣孔導度(Gs)增加較快。這說明光強的變化對小×胡楊氣孔運動有著非常大的影響。隨著光合有效輻射的增加,1年、2年、3年生小×胡楊氣孔導度(Gs)變化趨勢一致(圖4),均呈下降趨勢。當光合有效輻射<600 μmol·m-2·s-1,隨著光強的增加,1年、2年、3年生小×胡楊胞間CO2濃度(Ci)下降較快,當光合有效輻射>600 μmol·m-2·s-1,隨著光強的增加,1年、2年、3年生小×胡楊胞間CO2濃度(Ci)下降緩慢。
2.6 不同栽植年限小×胡楊水分利用效率光響應特征
在干旱荒漠地區,水分利用效率能夠體現植物適應干旱環境能力的強弱。隨著光合有效輻射的增加,1年、2年、3年生小×胡楊水分利用效率變化趨勢一致,均呈增加趨勢(圖5)。光合有效輻射<600μmol·m-2·s-1,1年、2年、3年生小×胡楊水分利用效率增加迅速,>600 μmol·m-2·s-1時,隨著光強的增強,3年生小×胡楊水分利用效率繼續緩慢上升,但而1年、2年生的小×胡楊水分利用效率趨于平緩甚至下降。

表2 不同栽植年限小×胡楊光合作用光響應曲線的特征參數
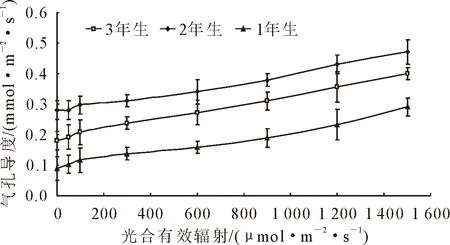
圖3 小×胡楊氣孔導度的光響應

圖4 小×胡楊胞間CO2濃度的光響應
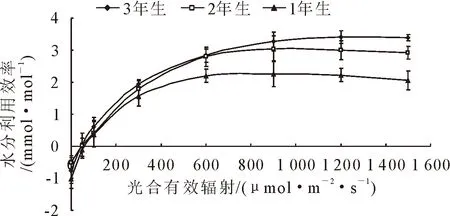
圖5 小×胡楊水分利用效率的光響應
3 結論與討論
3.1 討論
植物生長發育和生存與光有著密切聯系,植物光合作用所需能量的主要來源為光合有效輻射,光合有效輻射是影響植物光合作用重要因素[15]。表征植物光響應特性的指標主要包括光飽和速率(Amax),光飽和點(LSP)、光補償點(LCP)等,也是研究植物光合作用的主要生理指標,同時也是研究植物適應性的重要參數[16]。不同栽植年限小×胡楊的光飽和點(LSP)變化范圍1 722~1 763 μmol·m-2·s-1,比光補償點(LCP)31.22~31.99 μmol·m-2·s-1的變化幅度大(表2),說明不同栽植年限小×胡楊進行光合作用利用強光的能力較強,利用弱光的能力較小。3年生小×胡楊的光飽和速率(Amax)最高,達到28.32 μmol·m-2·s-1,1年生小×胡楊的光飽和速率(Amax)為19.14 μmol·m-2·s-1,說明3年生小×胡楊比1年生小×胡楊利用強光進行光合作用的能力較強。該研究中小×胡楊的光補償點為 31.22~31.99 μmol·m-2·s-1,光飽和點(LSP)為1 722.33~1 763.14 μmol·m-2·s-1,小×胡楊的光補償點和光飽和點比小葉楊[17](光補償點為28.97 μmol·m-2·s-1,光飽和點為1 680.60 μmol·m-2·s-1)的高,這表明小×胡楊比小葉楊對強光的適應能力更好、更喜光;小×胡楊的光補償點比胡楊[18]的光補償點(22.54 μmol·m-2·s-1)高,小×胡楊的光飽和點比胡楊[18](光飽和點為2 265.87 μmol·m-2·s-1)的低,說明小×胡楊對光強的利用范圍還有所不足。
植物在黑暗條件(無光照)的呼吸速率稱為暗呼吸速率[19]。研究顯示荒漠植物能夠很好的適應干旱區的環境,都具有獨特的生理機制,例如暗呼吸速率(RD)一般較高[20-22]。學者們認為有的植物存在光抑制現象,隨光強的增加,光合速率達到最大,光強繼續增強,光合速率呈下降趨勢[23];有的植物隨光強的增加,光合速率達到最大,光強繼續增強[24],光合速率保持平穩(一定范圍內),本研究當光合有效輻射(PAR)為1 500 μmol·m-2·s-1時,曲線處于緩慢上升趨勢,說明小×胡楊沒有達到光飽和點。下一步需要繼續增加光合有效輻射(PAR),探究小×胡楊達到光飽和點后的光響應特性。
光合有效輻射(PAR)0~1 500 μmol·m-2·s-1,1年、2年、3年生小×胡楊胞間CO2濃度隨著光合有效輻射(PAR)的增加變化規律一致,均呈下降趨勢。這是由于儀器葉室內的CO2濃度不變,胞間CO2濃度(Ci)越低,相應的具有越大的CO2濃度梯度值,CO2進入葉片內更為有利,更好地提供源物質,從而增強光合作用,提高光合能力的潛力。凈光合速率(Pn)與氣孔導度(Gs)的光響應顯示,1年、2年、3年生小×胡楊氣孔導度(Gs)變化趨勢一致,均呈增加趨勢。同時期的凈光合速率(Pn)變化較為迅速,說明在光和有效輻射(PAR)0~1 500 μmol·m-2·s-1下,CO2進入小×胡楊葉片后轉化速率快,提高了小×胡楊光合速率。
1年、2年、3年生小×胡楊氣孔導度及蒸騰速率變化趨勢相似。隨著氣孔導度增加,1年、2年、3年生小×胡楊的蒸騰速率逐漸增加。小×胡楊蒸騰速率(Tr)與氣孔導度(Gs)呈正相關。這與趙輝等[5]的研究結果一致。蒸騰速率(Tr)隨著光強的增加而迅速增加,光強逐漸增強,氣孔導度(Gs)逐漸增加,CO2進入葉片的阻力減小,植物葉片內CO2濃度增加,植物體內水分會大量散失(通過氣孔),導致蒸騰速率(Tr)上升較快。氣孔能夠自動調節開閉程度,使植物能夠更加適應環境變化。
烏蘭布和沙漠環境干旱少雨,光照強烈,荒漠植物生長發育的主要限制因子是水分,植物的水分利用效率越大,說明抗旱能力越強[25]。因此,為了適應沙漠干旱高溫特殊環境條件,植物必須具有較高的光合速率和水分利用效率[26-27],本研究不同栽植年限小×胡楊水分利用效率的光響應特性相似,隨著光強的增加,水分利用效率逐漸增加,不同栽植年限小×胡楊凈光合速率(Pn)隨著光和有效輻射(PAR)的增加而持續增加,蒸騰速率(Tr)隨著光和有效輻射(PAR)的增加而緩慢增加,表明小×胡楊凈光合速率隨光強的增加而增大,從而對水分的利用效率也越高。
研究顯示,影響植物對環境的適應性的因子非常多(植物根系、光合生理),植物自身特征也是關鍵因素[28]。因此,植物的適應性需要綜合考慮所有的影響因子。本研究發現1年、2年、3年生小×胡楊具有高光合速率、高水分利用效率及相對較低的蒸騰速率,對烏蘭布和沙漠東北緣環境的適應性較好。本研究只對1年、2年、3年生小×胡楊光合生理參數進行了探討,今后還需要對小×胡楊進行更加詳細的研究,才能對小×胡楊的適應性進行綜合評價。
3.2 結論
在光合有效輻射(PAR)0~1 500 μmol·m-2·s-1范圍內,不同栽植年限小×胡楊凈光合速率均隨著光合有效輻射(PAR)的增強而增加,當光合有效輻射(PAR)為1 500 μmol·m-2·s-1時,曲線處于緩慢上升趨勢,小×胡楊還沒有達到光飽和點。在光合有效輻射(PAR)0~1 500 μmol·m-2·s-1范圍內,不同栽植年限小×胡楊氣孔導度(Gs)隨光合有效輻射(PAR)的增加而增加,蒸騰速率隨著有效光輻射的增強而逐漸增加,變化趨勢都較緩慢平穩,蒸騰速率對光合有效輻射(PAR)的變化并不敏感,而胞間CO2濃度(Ci)均呈逐步下降的趨勢。當光合有效輻射<600 μmol·m-2·s-1,1年、2年、3年生小×胡楊水分利用效率增加迅速,當光合有效輻射>600 μmol·m-2·s-1,隨著光強的增強,3年小×胡楊水分利用效率繼續緩慢上升,但1年、2年生小×胡楊水分利用效率趨于平緩甚至下降。
1年、2年、3年生小×胡楊具有高光合速率、高水分利用效率及相對較低的蒸騰速率,對烏蘭布和沙漠東北緣環境的適應性較好。需要指出的是,今后還應該對小×胡楊生理生態特性進行深入細致的研究,才能綜合評價小×胡楊的適應性。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
