甜菜氮代謝相關(guān)蛋白BvAMT3-3基因的克隆及生物信息學(xué)分析
2021-08-09 09:36:56李佳佳劉銘曦馬龍彪劉大麗
黑龍江大學(xué)自然科學(xué)學(xué)報 2021年3期
關(guān)鍵詞:分析
魏 多, 李佳佳, 高 卓,2, 劉銘曦,馬龍彪, 興 旺, 劉大麗
(1.黑龍江大學(xué) 現(xiàn)代農(nóng)業(yè)與生態(tài)環(huán)境學(xué)院 省高校甜菜遺傳育種重點實驗室, 哈爾濱150080;2.黑龍江大學(xué) 生命科學(xué)學(xué)院, 哈爾濱 150080; 3.哈爾濱師范大學(xué) 生命科學(xué)與技術(shù)學(xué)院, 哈爾濱 150025)
0 引 言
作為世界第二大糖料作物,甜菜是除了甘蔗之外糖的主要來源之一,世界上約35%的糖是由甜菜供應(yīng)而來的[1]。甜菜屬于耐寒作物,在中國的種植區(qū)域主要位于東北三省、新疆和內(nèi)蒙古等地。由于耕地的過度使用以及作物的不規(guī)范化生產(chǎn),導(dǎo)致甜菜在種植過程中常常遭受氮脅迫等環(huán)境逆境的限制。植物在生長的過程中無時無刻不需要氮元素,并通過將無機氮轉(zhuǎn)化為有機氮的方式來吸收氮素[2]。合理施用氮肥可以有效提高作物產(chǎn)量,但施用過度會導(dǎo)致病蟲害增加和環(huán)境污染等問題[3-5]。基于此,使用銨基肥料和硝化抑制劑可減輕氮肥對環(huán)境的影響[6],根系對養(yǎng)分的吸收和運輸能力與不同植物器官的需求相匹配,這也進(jìn)一步?jīng)Q定了植物能否健康茁壯地生長。如果營養(yǎng)素可利用性下降或非生物性脅迫抑制了根系的正常功能,則會導(dǎo)致植物營養(yǎng)不足,降低它們的生態(tài)競爭力和農(nóng)業(yè)生產(chǎn)力[7]。
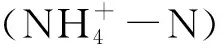
本文在對甜菜AMTs基因的前期研究中,通過對受氮逆境脅迫的甜菜進(jìn)行轉(zhuǎn)錄組測序發(fā)現(xiàn),在參與甜菜氮逆境脅迫相關(guān)的眾多基因中,BvAMT3-3基因的表達(dá)極其顯著。因此,利用RT-PCR的方法克隆了甜菜BvAMT3-3基因,并通過qPCR分析氮脅迫下目的基因的表達(dá)特性,結(jié)合生物信息學(xué)數(shù)據(jù)庫和生物軟件對BvAMT3-3的基因結(jié)構(gòu)和編碼的蛋白質(zhì)特點進(jìn)行了分析和預(yù)測,為進(jìn)一步了解甜菜AMTs基因以及甜菜中銨鹽的吸收機制奠定基礎(chǔ),對提高甜菜氮素的利用效率具有重要意義。
1 材料與方法
1.1 植物材料
供試甜菜為國家甜菜種質(zhì)資源中期庫提供的780016B/12優(yōu)。種子通過蛭石培養(yǎng)萌發(fā)后,將兩片子葉期的甜菜幼苗培養(yǎng)于N濃度為10 mmol·L-1的Hoagland營養(yǎng)液中,培養(yǎng)兩周后,取甜菜幼葉組織用于RNA的提取。
1.2 實驗方法
1.2.1 RNA的提取及cDNA的合成
利用Invitrogen公司的Trizol reagent提取甜菜新鮮材料的總RNA[17],并根據(jù)北京TransGen公司的反轉(zhuǎn)錄試劑盒合成cDNA,于-20 ℃保存?zhèn)溆谩?/p>
1.2.2BvAMT3-3基因的克隆
以甜菜總RNA反轉(zhuǎn)錄的cDNA為模板,利用KOD-Plus-Neo高保真酶(TOYOBO)PCR擴增該基因CDS全長。根據(jù)甜菜BvAMT3-3基因的堿基序列設(shè)計引物如下:
BvAMT3-3-F (5′-3′): CGGAA TTCATG GGA GAC ATA TTG CCA CCC
BvAMT3-3-R (5′-3′): GCTCT AGATTA TAC AAC TTG AGT TGC ACC AA
PCR產(chǎn)物電泳檢測后,將回收產(chǎn)物與載體pEASY Blunt連接,轉(zhuǎn)入T1感受態(tài)細(xì)胞。加入含LB的培養(yǎng)基后倒平板并搖置涂勻,37 ℃搖菌過夜后,篩選出陽性克隆的菌落進(jìn)行PCR檢測,選擇電泳檢測結(jié)果正確的菌液送上海生工生物工程公司進(jìn)行測序。
1.2.3 生物信息學(xué)分析
在NCBI數(shù)據(jù)庫中搜索甜菜BvAMT3基因家族成員;同時BvAMT3-3的開放閱讀框和CDS通過NCBI的ORF finder分析得到;利用NCBI保守結(jié)構(gòu)域數(shù)據(jù)庫預(yù)測該基因所編碼蛋白的保守結(jié)構(gòu)域。通過PSORT對甜菜AMT3-3蛋白進(jìn)行亞細(xì)胞定位;通過TMHHM分析跨膜結(jié)構(gòu)域數(shù)目。利用Prot-Param和ProtScale分析蛋白質(zhì)理化性質(zhì),SignalP 5.0 Server進(jìn)行信號肽預(yù)測分析。利用MEGA7軟件將甜菜AMT2家族的基因序列與其他植物的AMT2成員進(jìn)行聚類分析和Neighbor-Joining進(jìn)化樹的構(gòu)建。
1.2.4 植物培養(yǎng)及逆境處理
甜菜種子于蛭石中培養(yǎng)一周,待種子萌發(fā)后,選取生長良好的甜菜幼苗移栽入N濃度為0、1.5和10 mmol·L-1的Hoagland營養(yǎng)液中,N0為缺氮,N1.5為低氮,以10 mmol·L-1生長的甜菜幼苗為對照。取處理0、24和48 h時間點植株的相同部位葉片作為樣品,然后將甜菜幼苗移至正常培養(yǎng)條件,取恢復(fù)生長24和48 h的植株樣品,經(jīng)研磨后用于測量相關(guān)基因的表達(dá)量。
1.2.5BvAMT3-3基因的實時熒光定量PCR分析
根據(jù)甜菜BvAMT3-3基因(XM_010669058)堿基序列信息設(shè)計特異引物,以Trizol法提取的甜菜葉片RNA為模板,以BvGAPDH(NC_024800)基因為內(nèi)參,利用SuperReal PreMix Plus (TianGen)進(jìn)行qRT-PCR擴增,擴增時采用SYBR Green為標(biāo)記物。每個樣品3次重復(fù),利用2-△△CT方法對目的基因表達(dá)的數(shù)據(jù)進(jìn)行統(tǒng)計和分析。

表1 相關(guān)qRT-PCR引物序列及產(chǎn)物大小
2 結(jié)果與分析
2.1 甜菜BvAMT3-3基因的克隆
以甜菜總RNA反轉(zhuǎn)錄合成的cDNA為模板,用帶有EcoRI和Xbal酶切位點的特異性引物進(jìn)行RT-PCR擴增,獲得了一條大約1 420 bp的BvAMT3-3擴增片段,如圖1所示。
將BvAMT3-3基因經(jīng)載體連接,Kana陽性克隆篩選后,對所得的陽性克隆進(jìn)行質(zhì)粒提取,雙酶切電泳檢測。得到了大小為3 928 bp的載體pEASY-Blunt條帶和大小為1 427 bp的目的基因片BvAMT3-3的條帶,如圖2所示。
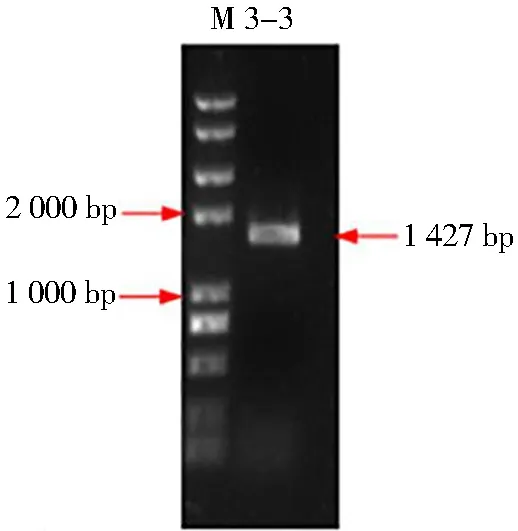
M: Trans 2K Plus II Marker(TransGen)
2.2 甜菜BvAMT3-3基因的序列分析
甜菜BvAMT3-3基因的CDS全長為1 427 bp,編碼475個氨基酸,如圖3所示。根據(jù)ProtParam分析計算,這個基因所編碼蛋白的分子量(Molecular weight)為51.055 kDa,等電點(Theoretical pI)為8.98。蛋白的不穩(wěn)定指數(shù)(Instability index)為29.61,屬于穩(wěn)定蛋白。根據(jù)SignalP 5.0分析,該蛋白沒有信號肽,不屬于分泌蛋白。對甜菜AMT蛋白二級結(jié)構(gòu)進(jìn)行預(yù)測分析,結(jié)果表明BvAMT3-3蛋白以α-螺旋為主,占43.79%,其次為無規(guī)則卷曲,占34.53%,而延伸鏈和β-轉(zhuǎn)角所占比例較小,如表2所示。亞細(xì)胞定位預(yù)測BvAMT3-3定位于內(nèi)質(zhì)網(wǎng)膜結(jié)構(gòu)上。


圖3 甜菜BvAMT3-3基因序列信息

表2 BvAMT3-3蛋白的二級結(jié)構(gòu)分析和亞細(xì)胞定位
2.3 BvAMT3-3蛋白的親水性和疏水性預(yù)測
用 ProtScale 軟件對蛋白質(zhì)進(jìn)行疏水性/親水性分析,正值越大表示越疏水,負(fù)值越大表示越親水,介于+0.5~-0.5之間的主要為兩性氨基酸。結(jié)果表明,BvAMT3-3蛋白的疏水區(qū)域大于親水區(qū)域,如圖4所示,預(yù)測該蛋白為疏水蛋白,這與ProtParam分析結(jié)果一致。

圖4 BvAMT3-3蛋白親疏水性預(yù)測
2.4 BvAMT3-3蛋白跨膜結(jié)構(gòu)域預(yù)測
運用TMHMM Server 2.0對BvAMT3-3蛋白的跨膜結(jié)構(gòu)域進(jìn)行預(yù)測。結(jié)果表明,BvAMT3-3含有11個跨膜螺旋區(qū),屬于跨膜蛋白,這與AMTs基因家族含有11個跨膜結(jié)構(gòu)域的結(jié)果相一致,如圖5所示。
2.5 甜菜BvAMT3-3蛋白保守結(jié)構(gòu)域分析
利用NCBI保守結(jié)構(gòu)域數(shù)據(jù)庫CD-Search預(yù)測該基因所編碼蛋白的保守結(jié)構(gòu)域。結(jié)果顯示,甜菜BvAMT3-3蛋白存在一個銨轉(zhuǎn)運保守結(jié)構(gòu)域,位置在25~443 aa,這保證了AMT在轉(zhuǎn)運銨鹽中發(fā)揮穩(wěn)定的作用。
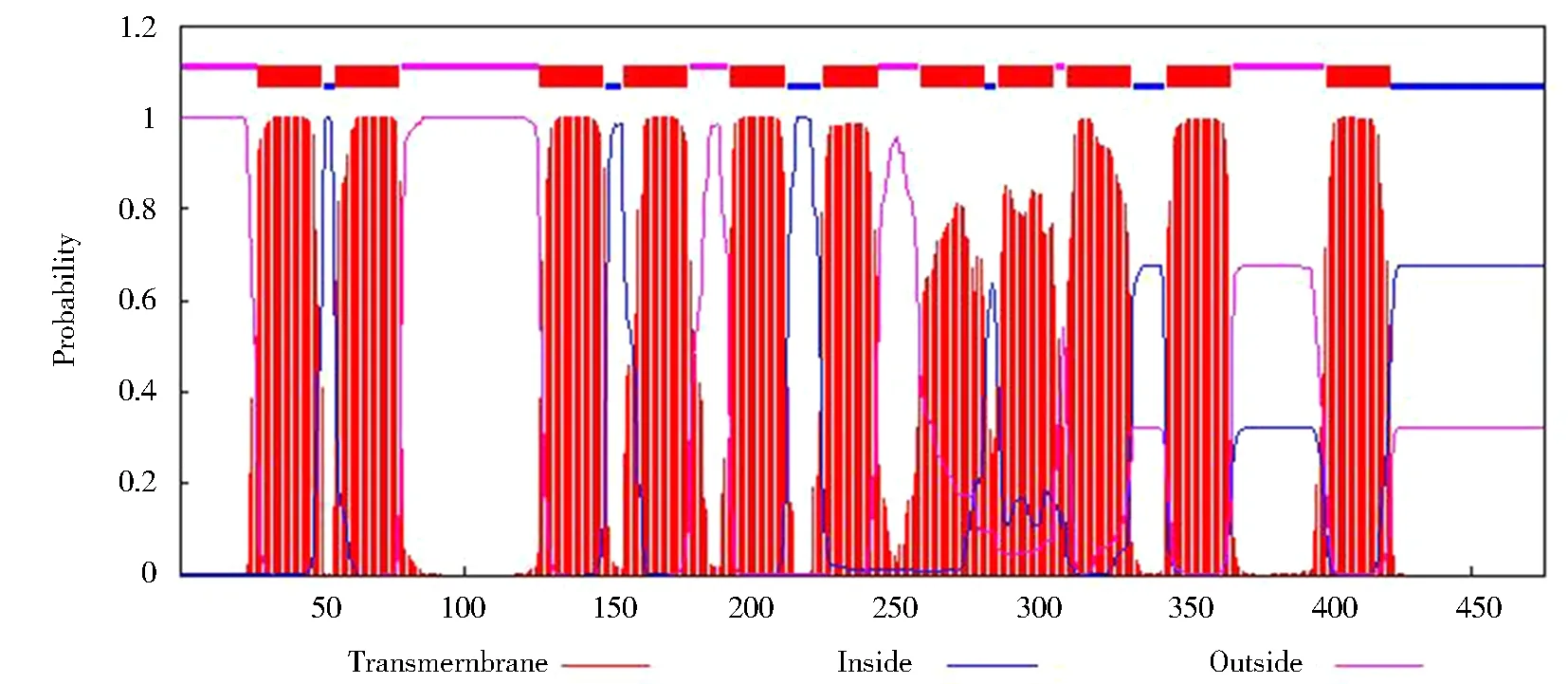
圖5 BvAMT3-3蛋白的跨膜螺旋區(qū)分析
2.6 甜菜BvAMT3-3蛋白三級結(jié)構(gòu)預(yù)測
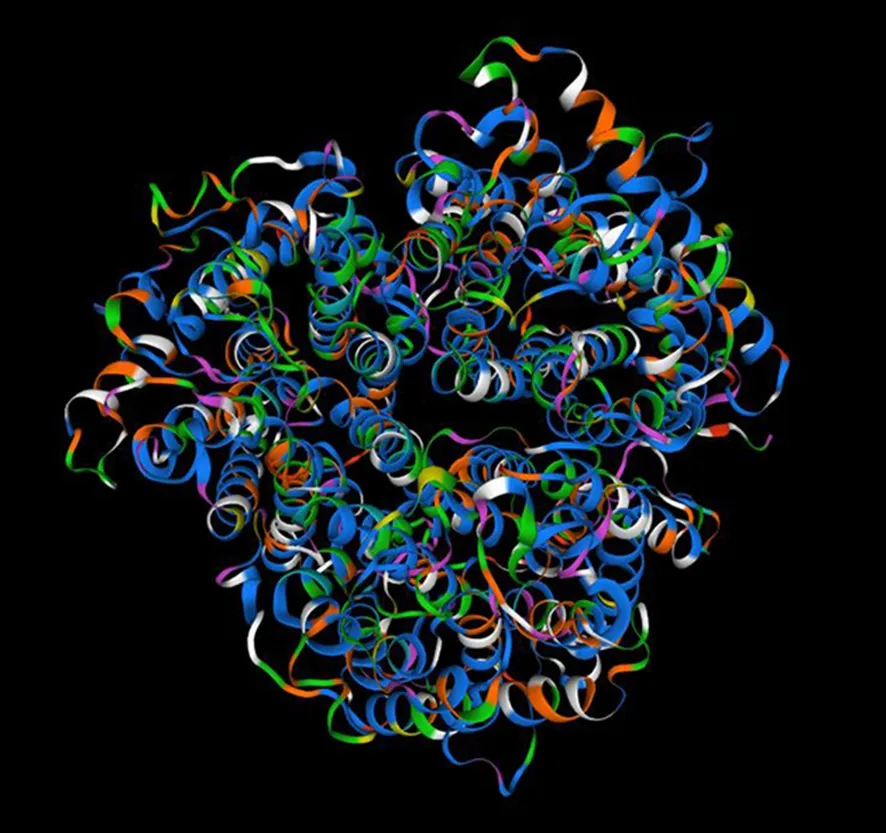
圖6 BvAMT3-3蛋白的三級結(jié)構(gòu)預(yù)測圖Fig.6 Prediction for tertiary structure of BvAMT3-3 protein
將甜菜BvAMT3-3的氨基酸序列提交到SWISSMODEL在線軟件,采用同源建模法預(yù)測該蛋白的三級結(jié)構(gòu),結(jié)果如圖6所示。結(jié)果顯示,該蛋白的三級結(jié)構(gòu)主要由無規(guī)則卷曲、延伸鏈和α-螺旋構(gòu)成,與二級結(jié)構(gòu)的預(yù)測結(jié)果基本一致。
2.7 甜菜BvAMT3-3基因的結(jié)構(gòu)及進(jìn)化樹分析
外顯子-內(nèi)含子的數(shù)量和結(jié)構(gòu)可以在一定程度上反映一些基因家族在進(jìn)化過程中的變化情況[18]。通過NCBI數(shù)據(jù)庫進(jìn)行BLAST搜索到5個甜菜AMT基因家族成員,利用GSDS 2.0對5個AMT基因結(jié)構(gòu)進(jìn)行分析。結(jié)果表明,BvAMT3-3基因含有2個內(nèi)含子,BvAMT1-1和BvAMT1-4均含有1個內(nèi)含子,而BvAMT1-2和BvAMT1-3均沒有內(nèi)含子,如圖7所示。對來自甜菜、菠菜、水稻、谷子等6個物種中的12個AMT3基因進(jìn)行系統(tǒng)進(jìn)化分析發(fā)現(xiàn),甜菜BvAMT3-3基因與菠菜SoAMT3-1基因位于同一進(jìn)化分支,如圖8所示。

圖7 甜菜AMT基因結(jié)構(gòu)特征

圖8 甜菜與其他物種之間的AMT3家族進(jìn)化樹
2.8 甜菜BvAMT3-3基因在低氮脅迫下的表達(dá)特性分析
AMT基因是參與植物氮逆境脅迫應(yīng)答的主要基因之一,它調(diào)控了植物體內(nèi)銨轉(zhuǎn)運蛋白的合成。在對甜菜進(jìn)行氮脅迫處理前,各組甜菜中BvAMT3-3的表達(dá)量幾乎基本處于相同水平,甜菜葉片部位的相對表達(dá)量在1.54左右,根部的表達(dá)量在1.27左右。當(dāng)缺氮(N0)和低氮(N1.5)處理24 h后,BvAMT3-3在葉片和根部的表達(dá)量均表現(xiàn)出明顯的上調(diào),并且隨著處理時間的增加,表達(dá)量呈現(xiàn)逐步上升的趨勢,且缺氮條件比低氮更容易誘導(dǎo)BvAMT3-3的表達(dá)。其中,葉片和根部的BvAMT3-3表達(dá)量均在48 h時達(dá)到最大,分別為0 h的4.62和3.32倍,可以看出N0處理更能誘導(dǎo)BvAMT3-3的表達(dá)。同時,該基因在葉片的表達(dá)比根部更為活躍,如圖9所示。恢復(fù)處理24和48 h后,BvAMT3-3表達(dá)量較脅迫處理時有所下降,但仍維持在較高水平。這說明即便恢復(fù)正常生長條件,氮逆境造成的細(xì)胞內(nèi)反應(yīng)還依舊存在,并且很有可能通過BvAMT3-3介導(dǎo)合成銨轉(zhuǎn)運蛋白,來調(diào)控氮代謝平衡。

3 討 論

4 結(jié) 論
本文利用RT-PCR克隆了基因BvAMT3-3,與生物信息學(xué)分析的結(jié)果相結(jié)合,發(fā)現(xiàn)該基因編碼的蛋白可能會作為轉(zhuǎn)運蛋白參與到甜菜對氮素的吸收和轉(zhuǎn)運網(wǎng)絡(luò)中。當(dāng)受到氮逆境脅迫時,BvAMT3-3基因表達(dá)量隨處理時間的增長呈現(xiàn)上升趨勢,表達(dá)的水平遠(yuǎn)大于未受到氮脅迫處理時的水平,表明該基因參與了甜菜對銨態(tài)氮吸收循環(huán)再利用的過程。甜菜受到氮逆境脅迫時,BvAMT3-3基因可能會參與到逆境脅迫的應(yīng)答網(wǎng)絡(luò)中,可能通過直接或間接的作用降低外界銨態(tài)氮含量的變化對細(xì)胞的損害,從而提高甜菜抵抗氮逆境脅迫的能力。研究結(jié)果有助于下一步研究不同氮水平條件下的基因表達(dá)特征、染色體定位和啟動子順式作用元件,同時也為加深植物吸收氮素的生理和分子機制的研究奠定基礎(chǔ)。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設(shè)計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(社會科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
