脂肪酶的作用機理及分泌調節研究進展
2021-08-09 02:00:36閆昭明劉霜莉黃華山陳清華
動物營養學報 2021年7期
閆昭明 劉霜莉 黃華山 陳清華*
(1.湖南農業大學動物科學技術學院,長沙 410128;2.山東隆科特酶制劑有限公司,臨沂 276400)
畜牧業中抗生素的不規范使用,導致了病原微生物對抗生素的耐藥性及畜產品中抗生素殘留等問題。自2020年起,我國施行飼料端全面“禁抗”新舉措。因此尋找良好的抗生素替代物將是目前的熱點研究工作。其中,飼用酶制劑在實際生產中已經得到了廣泛的應用。飼用不同種類酶制劑有助于提高動物對飼糧的養分消化率[1],從另一個角度也側面反映了飼糧營養價值的提高[2]。飼用酶制劑有助于加速消化道內營養物質的降解,并降低抗營養因子所帶來的負面影響,降低食糜黏度同時促進谷物飼糧中非淀粉多糖(non-starch polysaccharides,NSP)的消化并產生低聚糖,進而影響腸道內微生物的數量及組成[3]。動物內源可分泌三大類消化酶,分別為蛋白酶、淀粉酶、脂肪酶。其中蛋白酶和淀粉酶的研究較為完整和深入,但對于脂肪酶尤其是對于其分泌及消化的機制性研究還需要進一步完善。本文就脂肪酶的分類、作用機理、分泌調節進行綜述,以期為脂肪酶的應用提供參考。
1 脂肪酶的種類及作用機理
脂質、碳水化合物、蛋白質共同構成機體的產熱供能體系,也被稱為健康所必需的“三大營養素”[4],分別具有獨特的生理功能,同時是動物維持正常生命活動的根本保證。脂質除是動物機體內的能量來源外,還是質膜及細胞膜的重要組成部分,在細胞內信號傳導的過程中發揮介質的作用[5]。甘油三酯(triglyceride,TG)是動物機體內脂類物質的重要組成成分,主要儲存于脂肪組織中[6]。動物機體對于能量的攝入和消耗之間的平衡關系在一定程度上決定了生物體結構和功能的完整性。當能量攝入量大于消耗量時,未能被消耗的能量將會轉化為脂肪酸(fatty acid,FA),并通過發生酯化反應生產TG儲存于體內。而當能量消耗量較大時,TG也可發生脂解反應再次將FA釋放出來[7]。
脂肪酶又稱三酰基甘油酰基水解酶,是酯酶的一個分類,具有水解TG使之生成甘油和FA的能力。脂肪酶在自然界中來源廣泛,動物、植物及微生物均具有生產脂肪酶的能力,其中動物脂肪酶多存在于消化道及胰腺等器官中[8]。大多數種類脂肪酶的最適存活溫度在30~50 ℃,且適宜偏堿性的外部pH環境,活性也受金屬離子及有機溶劑的影響[9]。目前,對脂肪酶的研究主要集中于脂肪甘油三酯脂肪酶(adipose triglyceride lipase,ATGL)、激素敏感脂肪酶(hormone-sensitive lipase,HSL)、單酰甘油脂肪酶(monoacylglycerol lipase,MGL),這3種酶在機體內脂肪及非脂肪組織中對脂質代謝及分解具有明確的功能[10]。在TG的水解過程中,這3種酶表現出“順序”催化的功能。即ATGL首先將TG水解為二酰甘油(diacylglycerol,DAG)和1分子FA;再由HSL繼續催化,將DAG轉化為單酰甘油(monoacylglycerol,MAG)和1分子FA;最后由MGL發揮催化作用,將MAG分解為甘油和FA[11];為保證機體內水解平衡性,DAG和MAG還可通過二酰基甘油酰基轉移酶(diacylglycerol acyltransferase,DGAT)和單酰基甘油酰基轉移酶(monoacyglycerol acyltransferases,MGAT)發生酯化過程再次生成TAG[12],具體過程如圖1所示。
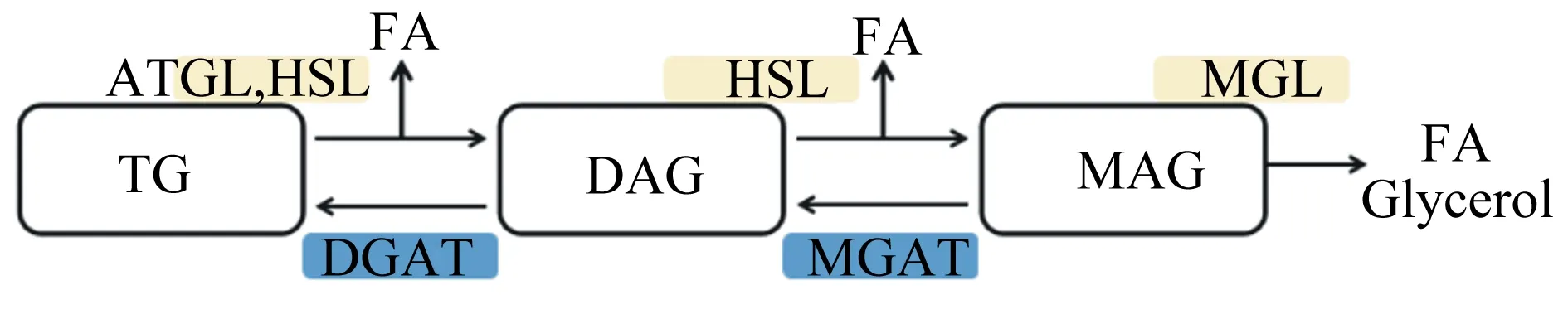
TG:甘油三酯 triglyceride;DAG:二酰甘油 diacylglycerol;MAG:單酰甘油 monoacylglycerol;FA:脂肪酸 fatty acid;Glycerol:甘油;ATGL:脂肪甘油三酯脂肪酶 adipose triglyceride lipase;HSL:激素敏感脂肪酶 hormone-sensitive lipase;MGL:單酰甘油脂肪酶 monoacylglycerol lipase;DGAT:二酰基甘油酰基轉移酶 diacylglycerol acyltransferase;MGAT:單酰基甘油酰基轉移酶 monoacyglycerol acyltransferases。
1.1 ATGL
在2004年首次發現Patatin-like磷脂酶的結構域2(PNPLA2),為簡化其描述,也稱為ATGL[13]。其主要功能為開始TG的水解過程,作為限速酶控制TG的水解速度[14],并被認為是哺乳動物機體內脂滴與脂質周轉過程中重要的水解酶[15]。通過ATGL免疫熒光發現其主要定位區域為細胞脂滴表面,說明其本質是一種脂滴蛋白[16]。
G0/G1開關基因2(G0/G1 switch gene 2,G0S2)與比較基因識別-58(comparative gene identification-58,CGI-58)這2種調節蛋白共同參與動物機體內ATGL的活化[17]。G0S2是一種重要的蛋白質,可通過調節細胞周期由G0期向G1期發生轉變,進而表現為對細胞增殖活動的調控作用。同時其可作為一種內源選擇性抑制劑抑制ATGL活性[18],其機制可描述為G0S2的疏水結構區與ATGL中Patatin結構域以非競爭方式結合[19],形成ATGL-G0S2復合體,從而減少TG的分解。CGI-58是ATGL發揮水解TG功能的輔助激活因子[20],其本身不具有水解酶活性。在小鼠試驗中發現,CGI-58數量的增加可提高TG水解酶的活性,但將ATGL基因敲除后卻未表現出類似效果,說明CGI-58是通過特異性激活ATGL進而促進TG的水解,且CGI-58的數量也決定了ATGL的活性上限[21]。
過氧化物酶體增殖物激活受體-γ(peroxisome proliferator-activated receptor-gamma,PPAR-γ)在脂肪生成和脂肪組織發育的過程中是一種關鍵的調控因子。在ATGL水解TG的過程中,所產生的脂肪酸可作為信號分子調控由PPAR-γ介導的酯化耦聯反應,以達到防止在脂質水解過程中出現內質網應激及TG的過量水解,維持脂肪細胞的結構及功能完整性[16]。此外,G0S2是PPAR-γ的靶基因,因其啟動子區域包括PPAR-反應元件(PPAR-response element,PPRE),PPAR-γ與PPRE發生特異性結合以上調G0S2基因的表達水平,進而對ATGL活性產生影響。G0S2在人和小鼠脂肪細胞形成過程中表達量顯著上調,提示G0S2在負調控ATGL活性的同時也參與了機體內脂肪形成[22]。
1.2 HSL
HSL在機體內脂肪動員(TG的逐步水解,其產物經血液運輸至不同組織被氧化利用的過程)的過程中發揮作用,即調動儲存的脂肪用于氧化供能[23]。其作為一種中性脂肪酶,對底物具有特異性的催化作用,主要分布于脂肪組織中,在肌肉及腎上腺、睪丸、胰腺等組織中也有少量表達[24]。多種抗脂解激素如胰島素、前列腺素,脂解激素如胰高血糖素、腎上腺素、促腎上腺皮質激素、促甲狀腺激素,可作用于HSL并發生磷酸化或去磷酸化反應從而調控其活性,其具有2個磷酸化位點,位點1(調節位點)可被蛋白激酶A(protein kinase,PKA)和糖原合酶激酶(glycogen synthase kinase,GSKS)磷酸化,外源應用胰島素可通過腺嘌呤核苷三磷酸(adenosine triphosphate,ATP)依賴的方式活化環磷酸腺苷(cyclic adenosine monophosphate,cAMP)磷酸二酯酶,以加速cAMP水解的形式抑制PKA活性,從而降低HSL活性[25]。近年來研究表明,白細胞介素-4(interleukin-4,IL-4)可通過增加cAMP數量,增強PKA的活性,使HSL的絲氨酸563及660位點發生磷酸化,以提高HSL活性,磷酸化HSL(p-HSL)可由細胞基質向脂滴表面發生轉移同時表現出解脂的功能[26]。位點2(基礎位點)磷酸化受AMP-活化蛋白激酶(AMP-activated protein kinase,AMPK)和Ca2+-鈣調蛋白依賴性蛋白激酶Ⅱ(calcium-CaM-dependent protein kinase Ⅱ,CaMK Ⅱ)共同調控[27]。
1.3 MGL
MGL屬于絲氨酸水解酶家族,是TG實現完全水解過程中的一種關鍵酶,也是MAG降解的限速酶[28]。在組織水平上,MGL主要存在于大腦、心臟、脂肪組織、腎臟、睪丸、卵巢等中并表現出特異性;在細胞層面上,MGL主要存在于細胞質、質膜及脂滴中[29]。對其底物選擇性的研究發現,相較于飽和脂肪酸形式的底物,MGL對不飽和脂肪酸形式的底物的水解活性較強,底物脂肪酸的不飽和程度對于MGL活性也有顯著影響,MGL對于花生四烯甘油的水解效率高于棕櫚酰甘油[30]。
MGL可將內源性大麻素(2-arachidonylglycerol,2-AG)水解為花生四烯酸(arachidonic acid,AA),AA作為一種二十烷酸衍生物的活性物質,具有豐富的生理功能,目前MGL-(2-AG)-AA水解代謝途徑已逐步成為研究熱點[31]。也有研究表明,MGL與多種病理及生理過程的發生有關,癌癥細胞中MGL水解TG的過程中誘導如溶血磷脂酸、前列腺素、溶血磷脂及溶血磷脂乙醇胺等信號分子的表達,其中溶血磷脂酸及前列腺素等可誘發腫瘤的發生和發展[32],提高生殖系統癌癥、黑色素瘤的發生率及轉移性,同時影響腸道功能的正常表達[33]。有研究表明,敲除MGL基因可通過抑制肝臟中關鍵的脂肪生成酶,如固醇調節元件結合蛋白-1c(sterol regulatory element binding protein-1c,SREBP-1c)、PPAR-γ2、二脂酰甘油酰基轉移酶-1(diacylgycerol acyhransferase-DGAT-1)的表達,從而促進FA氧化并防止小鼠肝臟脂肪變性的發生,但小鼠性腺中白色脂肪生成量增加,并未對脂肪分解產生影響[34]。
2 脂肪酶的分泌調節
動物消化道內主要包括舌脂肪酶、胃脂肪酶和胰脂肪酶3種,胃脂肪酶在脂質水解的過程中發揮輔助作用,并對胰脂肪酶的分泌起調控作用[35]。就分泌部位而言,舌脂肪酶及胃脂肪酶也可統稱為十二指腸前脂肪酶,胰脂肪酶稱為十二指腸內脂肪酶。由胰腺分泌的胰脂肪酶可直接影響小腸對于FA的吸收,故被認為是最重要的內源脂肪酶[9]。同時,胰脂肪酶分泌能力的強弱是衡量動物對脂肪消化能力高低的重要指標[36]。由肝細胞分泌產生的含有膽汁酸、甘油膽酸等成分的膽汁可對脂肪進行乳化,從而起到輔助脂肪酶消化的作用。
影響脂肪酶分泌的因素是多方面且復雜的。動物不同生理階段及健康狀況下脂肪酶活性均有差別,母豬哺乳期對脂質的利用效率最高[37]。畜禽均表現出“為能而食”的特點,添加油脂可提高豬飼糧能量水平。Liu等[38]通過應用不同能量水平飼糧飼喂斷奶仔豬后發現,高能飼糧組仔豬十二指腸及空腸內脂肪酶活性較高,提示飼糧能量水平與豬機體內脂肪酶的分泌存在交互效應。以上試驗結果在肉雞能量攝入與脂肪酶分泌的研究中得到了驗證[39]。此外,某些重金屬如銅元素可作為脂肪酶的激活劑,并通過抑制消化道內有害菌生長、繁殖而提高動物腸道健康水平,促進營養物質的利用[40]。克曌地[41]探究不同金屬離子在體外條件下對脂肪酶水解能力的影響,發現Sn2+、Fe3+、Fe2+、Ca2+、Mg2+均可促進脂肪酶水解能力的提高,但Zn2+則會降低脂肪酶的活性。激素對脂肪酶的分泌調節研究較為深入,HSL被認為是機體內脂解過程中受激素調控的主要因子,但對豬ATGL基因激素敏感性的研究發現,ATGL在激素誘導的脂肪水解過程中也發揮了作用。相較于自由采食,在限飼條件下,豬機體內胰島素水平較低,胰島素以負反饋調控形式促進ATGL的產生[42]。同時,飼糧組成及加工工藝、環境因素等都會對脂肪酶的分泌和脂質的消化吸收產生影響。
目前對于外源酶補充是否影響動物內源酶分泌的觀點主要分為2類:第1類即少量添加外源酶有提高機體消化酶水平的作用,但過量補充則抑制胰內源酶的分泌;第2類則不支持“反饋性抑制”理論,原因在于外源補充酶多由細菌或真菌在體外發酵生產,其結構和性質與內源酶存在區別[43]。為探究內、外源酶制劑是否存在交互分泌機制,應先探究酶制劑分泌的方式。以胰脂肪酶為例,胰腺內腺泡細胞核中基因發生轉錄作用生成mRNA,并通過翻譯、修飾、包膜作用形成酶原顆粒,后經高爾基體參與細胞分泌活動,以胞吐形式將酶釋放至胰管中,并進入十二指腸發揮酶解功能[44]。但目前,對于外源添加脂肪酶是否可以在轉錄及翻譯水平上調控動物機體內源脂肪酶的分泌還鮮有報道。
在畜禽的養殖實踐中,Liu等[38]在斷奶仔豬低能飼糧中添加150 mg/kg的脂肪酶后發現,其雖未能影響仔豬生長性能,但可在提高養分表觀消化率的基礎上同時提高十二指腸及回腸的脂肪酶活性,并對腸道內蛋白酶活性也有提高效應。Chen等[45]在仔豬飼糧中添加200 mg/kg的未包被脂肪酶或包被脂肪酶后也得到了類似的結果,且包被脂肪酶的效果更佳。包被的主要功能是可進一步保證酶的結構完整及生理活性,將脂肪酶在特定部位進行釋放,使之免受胃蛋白酶的消化從而降低損失[46]。Hu等[47]以肉雞為試驗動物,在低能飼糧中添加的脂肪酶也可促進胰脂肪酶活性的提高,并達到提高飼料轉化率及生產性能的目的。幼齡動物對脂質的消化能力較弱,主要與消化系統未能完全發育有關。斷奶應激是導致仔豬生長受限的主要原因,這一階段仔豬的營養物質來源從液體飼糧向固體飼糧過渡,仔豬脂肪酶、蛋白酶的分泌減少并引起腹瀉的發生[48]。動物消化道脂肪酶活性隨著日齡不斷增加,對脂質的利用效率得以提升[49],主要原因是對于飼糧中飽和脂肪酸的消化率增加[50]。3周后,仔豬脂肪酶分泌量達到正常水平,故仔豬階段外源補充脂肪酶的有效期為仔豬斷奶后3周內[51]。
3 小結與展望
脂肪酶在畜牧業中的應用具有提高動物生產性能、緩解飼料資源短缺、利于環境保護等功能,在仔豬生產中的應用更表現出降低腹瀉率的效果[52],是防治仔豬斷奶腹瀉的良好選擇之一。目前通過微生物發酵的手段生產脂肪酶的技術(液體深層發酵)已經逐步成熟,也已實現對曲霉、假單胞酶、根霉等高效發酵菌種的篩選工作。但是脂肪酶多適宜在堿性環境下發揮生理功能,而動物消化道內主要為酸性環境。故還需通過發酵菌種改造、基因改良工程、發酵工程及后期工藝處理等方面進行進一步優化。
目前研究者已形成的共識是使用酶制劑對動物的生長是有益的,伴隨單胃動物的不斷生長,酶制劑的功能從補充內源缺乏[53]向通過微生物路徑介導而對機體產生有益作用[54]。但在不同動物、不同生理階段中應用不同酶制劑(單一或復合)所得到的結果并不統一,脂肪酶的應用亦是如此。故對于脂肪酶的分泌、作用機制及在機體內與激素、其他營養物質之間的互作效應還值得進一步研究。
