竹節(jié)參總皂苷對妊娠期高血壓胎盤滋養(yǎng)細胞氧化應激損傷與過度自噬的影響
2021-07-21 02:28:30張化蓮張珊珊秦紅娟
中草藥 2021年14期
關 琦,張化蓮*,張珊珊,秦紅娟
1.駐馬店市中心醫(yī)院 產(chǎn)科,河南 駐馬店 463000
2.河南省人民醫(yī)院 婦產(chǎn)科,河南 鄭州 450000
妊娠期高血壓疾病(hypertension disorder complicating pregnancy,HDCP)是孕婦常見疾病之一,其發(fā)生與先兆子癇、剖宮產(chǎn)、腦血管疾病、胎兒生長受限、早產(chǎn)以及孕產(chǎn)婦和圍產(chǎn)兒死亡等密切相關[1-2]。HDCP使妊娠復雜化,嚴重危及母嬰安全[3]。HDCP病因和發(fā)病機制復雜,越來越多的研究表明,HDCP與多種因素引起的母胎代謝或免疫平衡有關,而胎盤滋養(yǎng)細胞作為妊娠建立和維持的基礎,其功能異常后會導致胎盤形成受阻,釋放大量炎性因子,使血管內(nèi)皮受損,最終導致子癇前期的發(fā)生[4-5]。
竹節(jié)參Panax japonicus(T.Nees) C.A.Mey.是五加科人參屬植物,竹節(jié)參總皂苷是其主要有效成分,具有鎮(zhèn)痛、抗衰老、抗氧化、抗炎、免疫調(diào)節(jié)以及神經(jīng)保護效應等作用[6-7]。竹節(jié)參總皂苷對Triton WR-1339誘導的高脂血癥小鼠具有一定的調(diào)血脂作用[8],而竹節(jié)參總皂苷在HDCP中的作用至今卻未見報道。人胎盤滋養(yǎng)細胞株HTR-8/SVneo是一種永生化的滋養(yǎng)層細胞系,具有許多與人類原代滋養(yǎng)層細胞相似的特性,目前已成為研究胎盤功能和妊娠相關疾病的主要細胞系。本研究采用亞硝基左旋精氨酸甲酯(N-nitro-L-arginine methylester,L-NAME)干預HTR-8/SVneo細胞模擬HDCP微環(huán)境,探討竹節(jié)參總皂苷對HDCP胎盤滋養(yǎng)細胞氧化應激損傷及自噬的影響,并初步探究其作用機制,從而為HDCP的研究及診治提供實驗基礎。公司;CCK-8試劑盒(批號42022ES34)和Annexin V-FITC/碘化丙啶(PI)細胞凋亡檢測試劑盒(批號40302ES20)購自北京全式金生物科技公司;丙二醛(malondialdehyde,MDA)檢測試劑盒(批號A00312)、超氧化物歧化酶(superoxide dismutase,SOD)檢測試劑盒(批號A00132)、谷胱甘肽(glutataione,GSH)檢測試劑盒(批號A00512)購自南京建成生物工程研究所;TRIzol試劑盒(批號15596-026)、SYBR?Premix Ex TaqTMII試劑盒(批號RR047A)購自日本Takara公司;逆轉(zhuǎn)錄試劑盒(批號050720190702)購自上海生工生物工程有限公司;細胞裂解液(批號P0013)、BCA蛋白定量試劑盒(批號P0010S)、PVDF膜(批號P0016S)購自上海碧云天生物技術有限公司;脂肪酸結合蛋白4(fatty acid binding protein 4,F(xiàn)ABP4)抗體(批號3579S)、微管相關蛋白1輕鏈3-II(microtubule associated protein 1 light chain 3 II,LC3-II)抗體(批號3868S)、LC3-I抗體(批號4599S)、p62抗體(批號3534S)、自噬效應蛋白(Beclin-1)抗體(批號3495S)購自美國CST公司;甘油醛-3-磷酸脫氫酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)抗體(批號TA-1203)、HRP標記的山羊抗兔抗體(批號ZB-2306)購自北京中杉金橋生物有限公司。
1.3 儀器
1 材料
1.1 細胞
HTR-8/SV-neo細胞購自上海瑞鹿生物技術有限公司。
1.2 藥品與試劑
竹節(jié)參總皂苷(質(zhì)量分數(shù)為85.4%)由首都醫(yī)科大學中醫(yī)藥學院中藥化學實驗室提供;L-NAME(批號1115878)購自美國Sigma公司;胎牛血清(批號2166446)、青霉素-鏈霉素(批號20190909)購自美國Gibco公司;DMEM/F12培養(yǎng)基(批號C11995500CP)購自美國Thermo Fisher Scientific
BC-J160S細胞培養(yǎng)箱(上海博迅實業(yè)有限公司醫(yī)療設備廠);低溫高速離心機(德國Heraeus公司);MACSQuant Analyzer流式細胞儀(德國Miltenyi公司);光學顯微鏡(日本Olympus公司);DYY-6C電泳儀(北京六一儀器廠);PCR擴增儀(美國ABI公司);M340651酶標儀(北京中西遠大科技有限公司)。
2 方法
2.1 細胞培養(yǎng)
HTR-8/SV-neo細胞用含10%胎牛血清、青霉素(100 U/mL)、鏈霉素(100 μg/mL)的DMEM/F12培養(yǎng)基,于37 ℃、5% CO2的培養(yǎng)箱中培養(yǎng),每2天換液1次,待細胞融合度達到80%~90%時進行傳代,使用第3~5代細胞進行后續(xù)實驗。
2.2 竹節(jié)參總皂苷對HTR-8/SV-neo細胞活力的影響
取處于對數(shù)生長期的HTR-8/SV-neo細胞,以4×104/mL接種至96孔板,培養(yǎng)12 h。設置對照組、模型組和竹節(jié)參總皂苷(20、40、80 μg/mL)[9]組,模型組和各給藥組加入L-NAME(100 μmol/L)處理48 h;各給藥組再加入相應藥物,對照組加入不含藥物的培養(yǎng)基,培養(yǎng)24 h,加入10 μL CCK-8試劑,孵育1 h,采用酶標儀測定450 nm處的吸光度(A)值。
2.3 竹節(jié)參總皂苷對HTR-8/SV-neo細胞凋亡的影響
按“2.2”項下方法處理細胞和分組,收集細胞,以PBS洗滌2次,加入500 μL 1×Binding Buffer重懸,加入5 μL Annexin V-FITC和PI試劑,室溫避光孵育15 min,采用流式細胞儀檢測細胞凋亡情況。
2.4 竹節(jié)參總皂苷對HTR-8/SV-neo細胞上清液中MDA、SOD和GSH的影響
按“2.2”項下方法處理細胞和分組,收集上清液,按試劑盒說明書檢測細胞上清液中MDA、SOD和GSH水平。
2.5 竹節(jié)參總皂苷對HTR-8/SV-neo細胞FABP4、LC3-Ⅱ、LC3-Ⅰ、p62和Beclin-1蛋白表達的影響
按“2.2”項下方法處理細胞和分組,收集細胞,加入裂解液,10 000×g離心20 min,取上清,采用BCA蛋白定量試劑盒測定蛋白質(zhì)量濃度。蛋白樣品經(jīng)10%十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳,轉(zhuǎn)至PVDF膜,加入5%脫脂牛奶于室溫封閉2 h,分別加入FABP4、LC3-II、LC3-I、p62、Beclin-1和GAPDH抗體(1∶1000),4 ℃孵育過夜;TBST洗滌后,加入HRP標記的山羊抗兔抗體(1∶5000),室溫孵育2 h,加入ECL化學發(fā)光液,采用凝膠成像儀曝光成像,BandScan5.0軟件分析條帶灰度值。
2.6 竹節(jié)參總皂苷對HTR-8/SV-neo細胞FABP4 mRNA表達的影響
按“2.2”項下方法處理細胞和分組,收集細胞,按照試劑盒說明書提取細胞總RNA并合成cDNA,進行qRT-PCR分析。引物序列:FABP4上游引物5’-AAGAGAAAACGAGAGGATGATAAAC-3’,下游 引 物5’-ATGCGAACTTCAGTCCAGGTC-3’;GAPDH上游引物5’-AGGGGTCTACATGGCAACTG-3’,下 游 引 物5’-CGACCACTTTGTCAAGCTCA-3’。
2.7 竹節(jié)參總皂苷對HTR-8/SV-neo細胞LC3表達影響
取處于對數(shù)生長期的HTR-8/SV-neo細胞,以1×104/孔接種至24孔板內(nèi)的無菌玻片上,培養(yǎng)12 h。按“2.2”項下方法分組,收集細胞,PBS洗滌,于4%多聚甲醛中室溫固定20 min,加入0.5%TritonX-100透膜15 min,加入10%山羊血清室溫封閉2 h,加入LC3抗體(1∶150),4 ℃孵育過夜;PBS洗滌,滴加Alexa Fluor 488標記的山羊抗兔IgG抗體,室溫避光孵育1 h;PBS洗滌,滴加DAPI染色10 min;PBS洗滌,封片,于熒光顯微鏡下觀察細胞LC3表達情況。
2.8 統(tǒng)計學分析
采用SPSS 22.0軟件進行數(shù)據(jù)分析。數(shù)據(jù)以±s表示,采用單因素方差分析,組間兩兩比較采用LSD-t檢驗。
3 結果
3.1 竹節(jié)參總皂苷對HTR-8/SV-neo細胞活力的影響
如圖1所示,與對照組比較,模型組細胞活力顯著降低(P<0.05);與模型組比較,各給藥組細胞活力顯著升高(P<0.05)。
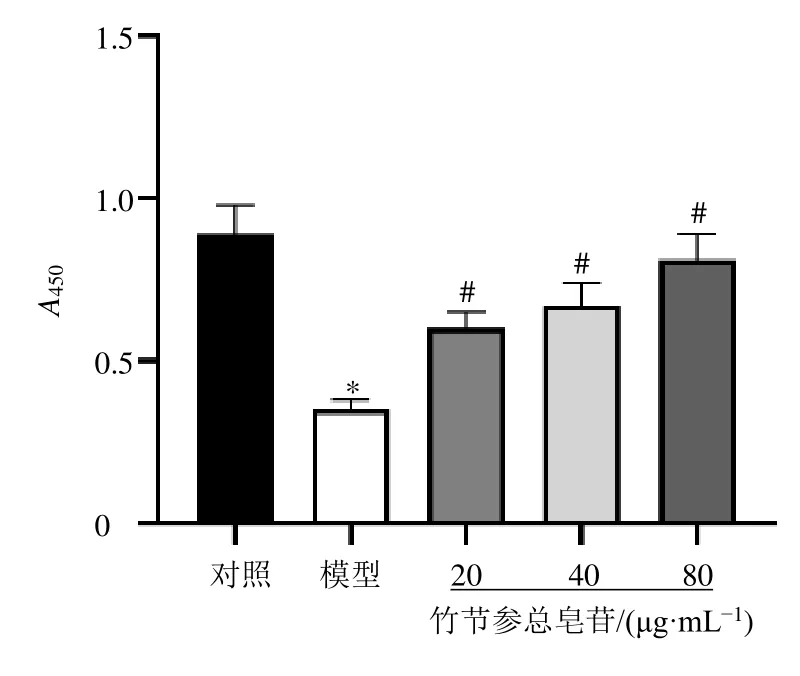
圖1 竹節(jié)參總皂苷對HTR-8/SV-neo細胞活力的影響(±s , n=3)Fig.1 Effect of total saponins from P.japonicus on viability of HTR-8/SV-neo cells (±s , n=3)
3.2 竹節(jié)參總皂苷對HTR-8/SV-neo細胞凋亡的影響
如圖2所示,與對照組比較,模型組細胞凋亡率顯著升高(P<0.01);與模型組比較,各給藥組細胞凋亡率顯著降低(P<0.01)。
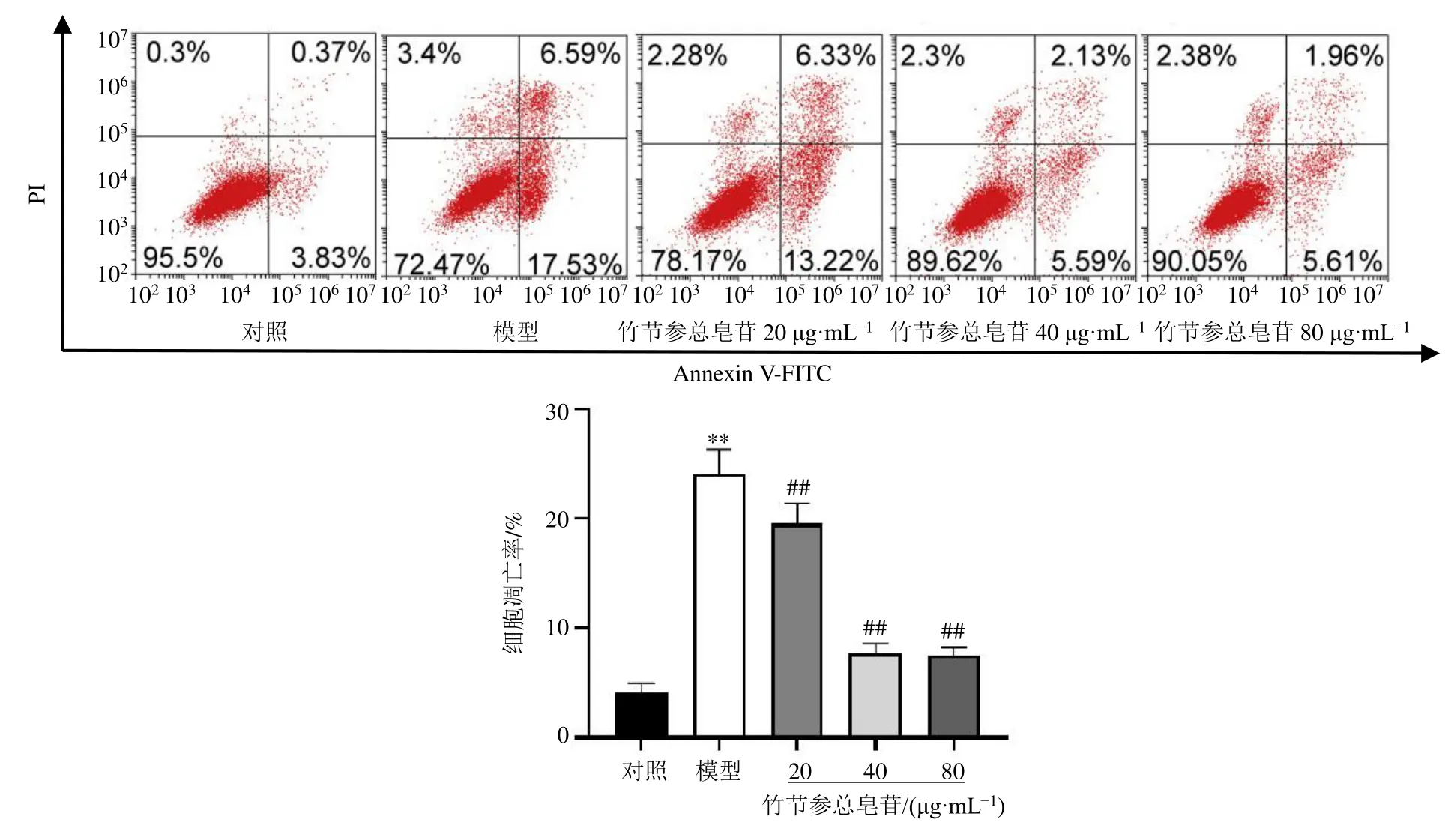
圖2 竹節(jié)參總皂苷對HTR-8/SV-neo細胞凋亡的影響 (±s , n=3)Fig.2 Effect of total saponins from P.japonicus on apoptosis of HTR-8/SV-neo cells (±s , n=3)
3.3 竹節(jié)參總皂苷對HTR-8/SV-neo細胞上清液中MDA、SOD和GSH的影響
如表1所示,與對照組比較,模型組細胞上清液中SOD和GSH水平顯著降低(P<0.01),MDA水平顯著升高(P<0.01);與模型組比較,各給藥組細胞上清液中SOD和GSH水平顯著升高(P<0.01),MDA水平顯著降低(P<0.01)。
表1 竹節(jié)參總皂苷對HTR-8/SV-neo細胞上清液中MDA、SOD和GSH的影響 (±s , n=3)Table 1 Effect of total saponins from P.japonicus on levels of MDA, SOD and GSH in supernatant of HTR-8/SV-neo cells(±s , n=3)

表1 竹節(jié)參總皂苷對HTR-8/SV-neo細胞上清液中MDA、SOD和GSH的影響 (±s , n=3)Table 1 Effect of total saponins from P.japonicus on levels of MDA, SOD and GSH in supernatant of HTR-8/SV-neo cells(±s , n=3)
與對照組比較:**P<0.01;與模型組比較:##P<0.01**P < 0.01 vs control group; ##P < 0.01 vs model group
組別 劑量/(μg·mL-1) MDA/(μmol·L-1) SOD/(μmol·L-1) GSH/(μmol·L-1)對照 — 6.56±0.62 104.01±9.77 112.81±10.21模型 — 25.87±2.31** 54.01±5.51** 64.13±6.76**竹節(jié)參總皂苷 20 17.06±2.01## 69.81±6.49## 72.84±7.62##40 12.21±1.45## 78.60±7.33## 90.61±8.81##80 10.53±0.91## 80.41±7.92## 96.89±9.07##
3.4 竹節(jié)參總皂苷對HTR-8/SV-neo細胞FABP4 mRNA和蛋白表達的影響
如圖3所示,與對照組比較,模型組細胞FABP4mRNA和蛋白表達水平均顯著升高(P<0.05);與模型組比較,各給藥組細胞FABP4mRNA和蛋白表達水平均顯著降低(P<0.05)。
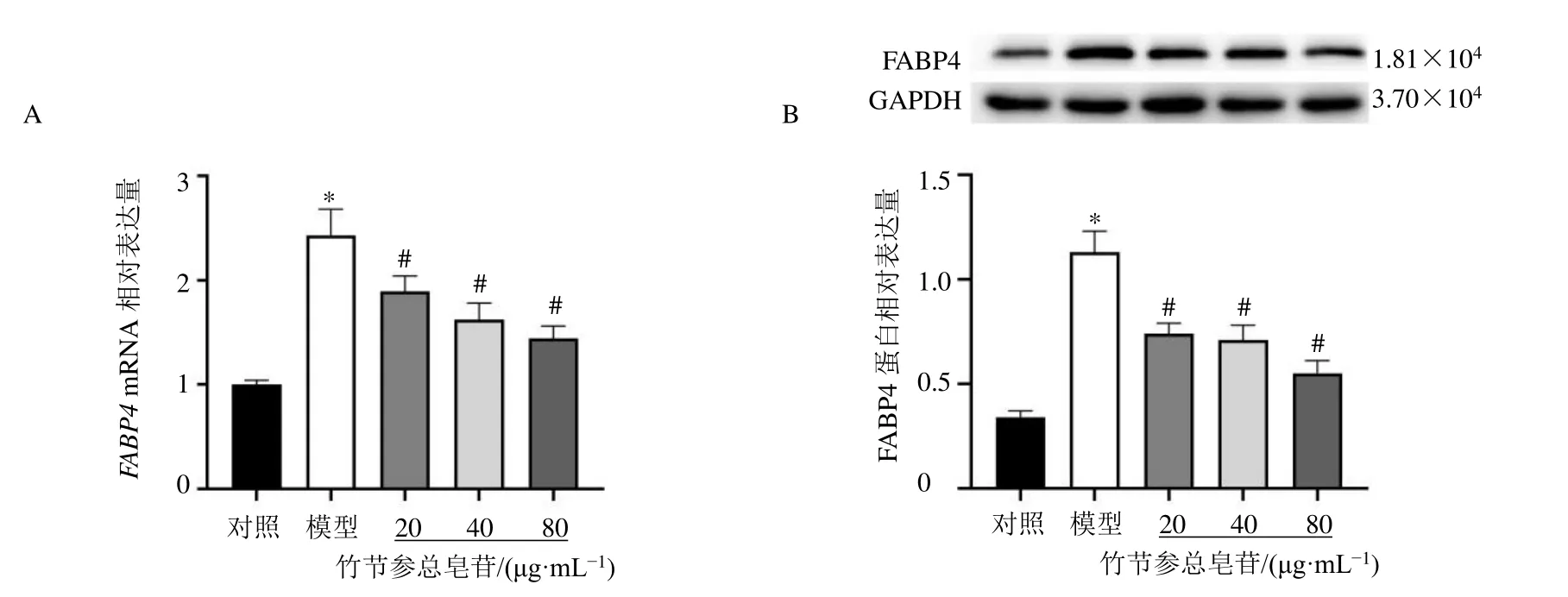
圖3 竹節(jié)參總皂苷對HTR-8/SV-neo細胞FABP4 mRNA (A) 和蛋白 (B) 表達的影響 (±s , n=3)Fig.3 Effect of total saponins from P.japonicus on expressions of FABP4 mRNA (A) and protein (B) in HTR-8/SV-neo cells(±s , n=3)
3.5 竹節(jié)參總皂苷對HTR-8/SV-neo細胞LC3-Ⅰ、LC3-Ⅱ、p62和Beclin-1蛋白表達的影響
如圖4所示,與對照組比較,模型組細胞LC3-Ⅱ和Beclin-1蛋白表達水平均顯著升高(P<0.05),p62蛋白表達水平顯著降低(P<0.05);與模型組比較,各給藥組LC3-Ⅱ和Beclin-1蛋白表達水平均顯著降低(P<0.05),竹節(jié)參總皂苷(40、80 μg/mL)組p62蛋白表達水平顯著升高(P<0.05)。
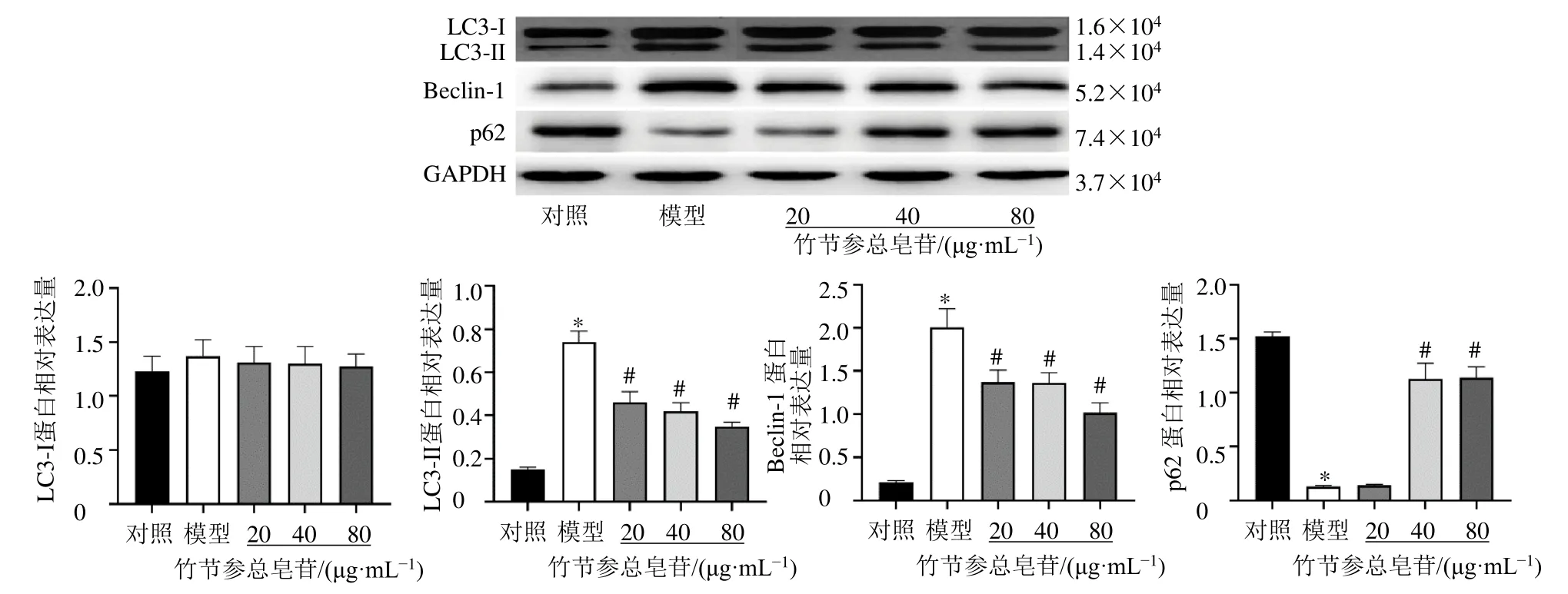
圖4 竹節(jié)參總皂苷對HTR-8/SV-neo細胞LC3-Ⅰ、LC3-Ⅱ、p62和Beclin-1蛋白表達的影響 (±s , n=3)Fig.4 Effect of total saponins from P.japonicus on expressions of LC3-Ⅰ, LC3-Ⅱ, p62 and Beclin-1 in HTR-8/SV-neo cells(±s , n=3)
3.6 竹節(jié)參總皂苷對HTR-8/SV-neo細胞LC3表達的影響
如圖5所示,對照組細胞LC3熒光染色弱,模型組細胞LC3熒光染色較強,各給藥組細胞LC3熒光染色較模型組降低。
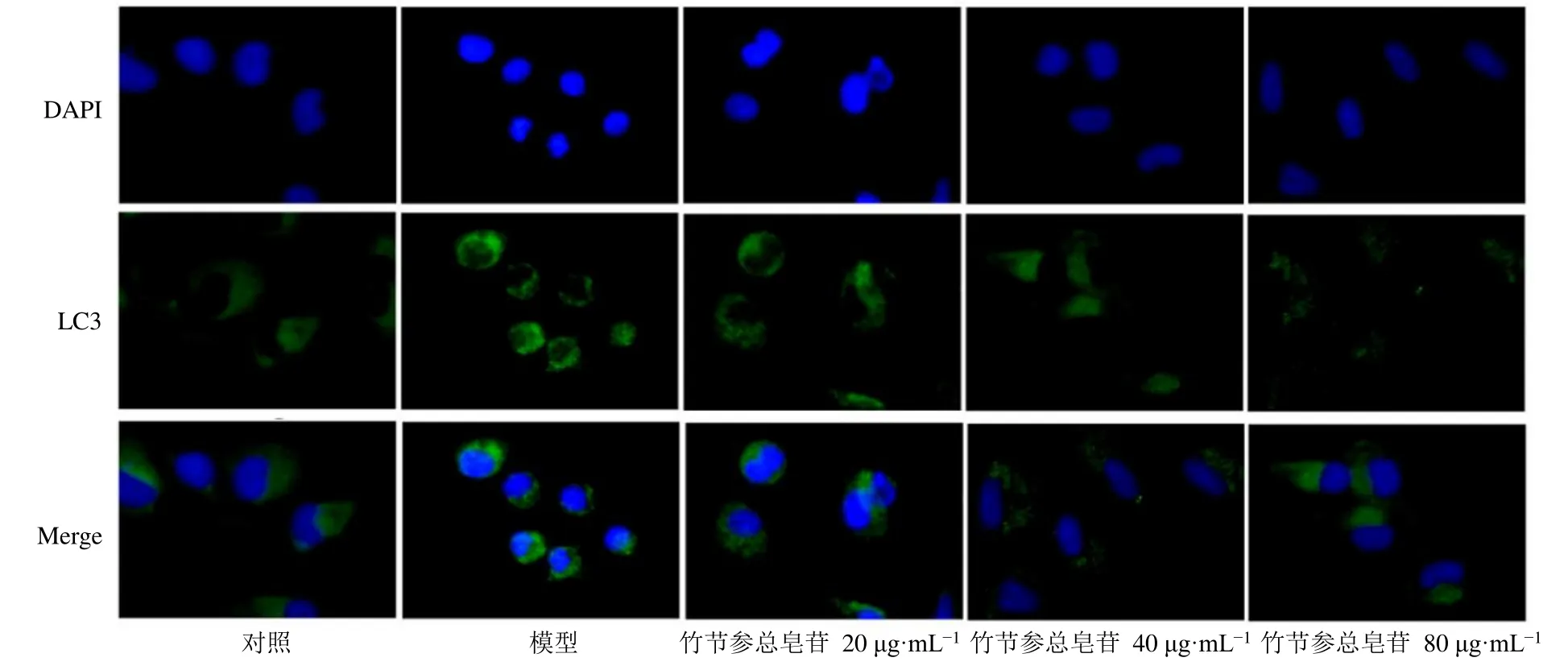
圖5 竹節(jié)參總皂苷對HTR-8/SV-neo細胞LC3表達的影響 (×100)Fig.5 Effect of total saponins from P.japonicus on LC3 expression in HTR-8/SV-neo cells (× 100)
4 討論
近年來HDCP約占妊娠的10%,盡管目前關于HDCP已開展了大量的研究,但HDCP仍然是婦產(chǎn)科中最棘手的難題[2]。HDCP的發(fā)病機制起源于胎盤異常,其中心環(huán)節(jié)是滋養(yǎng)細胞功能異常所致的胎盤生理性障礙。胎盤發(fā)揮著母體與胎兒之間營養(yǎng)物和氧氣交換的作用,從而使胎兒能夠正常生長和發(fā)育。當胎盤功能受損時,胎兒的生長受到影響,從而導致胎兒生長受限[10]。因此,胎盤滋養(yǎng)細胞在HDCP發(fā)病中起著關鍵作用。以一氧化氮合酶抑制劑L-NAME干預人胎盤滋養(yǎng)細胞,可以模擬HDCP的微環(huán)境,從而開展HDCP的相關研究[11-12]。本研究結果顯示,經(jīng)L-NAME處理后,HTR-8/SV-neo細胞活力明顯降低,細胞出現(xiàn)大量的凋亡,表明HDCP中可能會引發(fā)胎盤滋養(yǎng)細胞的生物學功能異常;竹節(jié)參總皂苷能夠提高HTR-8/SV-neo細胞活力,并抑制細胞的異常凋亡現(xiàn)象,表明竹節(jié)參總皂苷對HDCP中的胎盤滋養(yǎng)細胞可能發(fā)揮一定作用。
氧化劑與抗氧化劑的產(chǎn)生以及能力之間的不平衡會導致氧化應激反應。活性氧在組織和器官中的積累會誘發(fā)心血管疾病、神經(jīng)系統(tǒng)并發(fā)癥、癌癥、呼吸系統(tǒng)疾病、類風濕性關節(jié)炎以及與妊娠相關疾病的發(fā)生[13-14]。與正常孕婦相比,患有HDCP的孕婦體內(nèi)氧化應激反應增加。SOD和GSH具有抗氧化物酶性質(zhì),能夠通過協(xié)同清除活性氧和自由基來減輕氧化性損傷;MDA是脂質(zhì)過氧化產(chǎn)物,其含量的高低可以體現(xiàn)氧化反應的狀態(tài)[15]。本研究結果顯示,L-NAME干預后HTR-8/SV-neo細胞上清液中SOD和GSH水平降低,MDA水平升高,表明HTR-8/SV-neo細胞發(fā)生了氧化應激反應;竹節(jié)參總皂苷組細胞上清液中SOD和GSH水平升高,MDA水平降低,表明氧化應激反應被抑制。
自噬是細胞內(nèi)分解代謝的穩(wěn)態(tài)機制,可以將細胞內(nèi)組分轉(zhuǎn)移至溶酶體進行降解和氨基酸循環(huán)。Beclin-1是自噬的關鍵調(diào)節(jié)劑,在其啟動和進程中均起著重要作用,可以通過激活各種細胞信號通路調(diào)控自噬發(fā)生;存在于細胞質(zhì)中的p62能夠與經(jīng)泛素化蛋白結合,并與LC3-II結合形成復合物,在溶酶體內(nèi)發(fā)生降解。自噬是維持細胞應激反應穩(wěn)態(tài)的重要機制之一,能夠在早期胎盤缺氧和營養(yǎng)受限的環(huán)境中對滋養(yǎng)層細胞起到重要的保護作用,但過度的自噬可能會破壞細胞結構并誘導細胞發(fā)生異常死亡[16-17]。然而,自噬是否與妊娠相關疾病的胎盤功能障礙的病理過程有關,是否存在由于氧化應激損傷而導致滋養(yǎng)細胞過度自噬,以及這些過程發(fā)生的潛在機制尚待證實。本研究結果顯示,經(jīng)L-NAME干預后,HTR-8/SV-neo細胞LC3熒光染色增強,LC3-Ⅱ、Beclin-1蛋白表達水平均升高,p62蛋白表達水平降低,表明HTR-8/SV-neo細胞發(fā)生過度自噬;竹節(jié)參總皂苷組LC3-Ⅱ、Beclin-1蛋白表達水平降低,p62蛋白表達水平升高,表明竹節(jié)參總皂苷能夠改善HDCP中胎盤滋養(yǎng)細胞的過度自噬現(xiàn)象。
FABPs是脂質(zhì)分子伴侶的家族成員,能夠通過調(diào)節(jié)幾種脂質(zhì)信號通路來促進系統(tǒng)性代謝。FABP4在細胞和組織中的異常表達與疾病的發(fā)病機制密切相關,通過特異性抑制劑、中和抗體或未知受體拮抗劑對FABP4功能的藥理修飾將是針對肥胖癥、糖尿病、動脈粥樣硬化等多種疾病的新型治療策略[18-19]。Tu等[20]通過檢測妊娠期前3個月患者血漿中FABP4水平發(fā)現(xiàn),妊娠早期較高的FABP4水平與妊娠期糖尿病風險增加有關。在肥胖誘發(fā)的非酒精性脂肪肝中,F(xiàn)ABP4表達顯著增加,過氧化酶體增殖物激活受體 γ(peroxisome proliferator activated receptor γ,PPARγ)具有抑癌基因的作用,而FABP4能夠促進疾病的發(fā)生[21]。本研究結果顯示,經(jīng)L-NAME干預后,HTR-8/SV-neo細胞中FABP4mRNA和蛋白表達水平明顯升高;竹節(jié)參總皂苷顯著抑制FABP4mRNA和蛋白表達水平,表明竹節(jié)參總皂苷可能通過調(diào)控FABP4表達來改善HDCP中胎盤滋養(yǎng)細胞損傷。
綜上所述,竹節(jié)參總皂苷能夠抑制L-NAME干預的胎盤滋養(yǎng)細胞活力降低以及細胞凋亡,并改善細胞的氧化應激損傷與過度自噬,其機制可能與調(diào)控FABP4表達有關。
利益沖突所有作者均聲明不存在利益沖突
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數(shù)理化·七年級數(shù)學人教版(2020年10期)2020-11-26 08:24:50
數(shù)學物理學報(2020年2期)2020-06-02 11:29:24
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
光學精密工程(2016年6期)2016-11-07 09:07:19
學苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
核科學與工程(2015年4期)2015-09-26 11:59:03
中國火炬(2010年8期)2010-07-25 11:34:30