基于SSR標記的海南栽培檳榔遺傳多樣性分析
2021-07-20 14:32:28齊蘭王世正黃麗云周煥起劉立云
熱帶作物學報 2021年5期
齊蘭 王世正 黃麗云 周煥起 劉立云


摘 ?要:檳榔是海南重要的熱帶特色經濟作物,具有很高的藥用價值,被列為“四大南藥”之首。但其基礎研究落后,缺乏對種質資源的系統研究,遺傳多樣性尚不明確。利用SSR分子標記技術對海南島58份栽培檳榔資源進行遺傳多樣性分析,旨在明確檳榔栽培種的親緣關系,為檳榔雜交育種、功能基因的挖掘提供理論依據。結果表明:從500對SSR引物中篩選出32對多態性好、條帶清晰的引物,在58份種質資源中共檢測到79個等位基因,每個SSR位點2~5個,平均2.4688個,其中有效等位變異占觀測等位變異的58.92%。各引物PIC值變幅為0.0169~0.5969,平均值為0.2254,Shannon信息指數(I)為0.0496~1.2552,平均值為0.4496;Neis遺傳多樣性指數(Nei)變幅為0.0171~0.6492,平均值為0.2596,說明所選SSR引物在供試樣品中檢測等位基因遺傳多樣性偏低。利用UPGMA構建58份栽培檳榔的遺傳聚類樹狀圖,第V組材料遺傳多樣性最豐富,其他分組群體遺傳多樣性較低,聚類結果說明海南栽培檳榔品種無明顯的區域劃分,表明海南檳榔不同地區間的種苗交流頻繁,總體遺傳多樣性水平較低,該研究為檳榔遺傳育種親本選擇提供依據,也為今后引進種質增補差異資源提供參考。
關鍵詞:檳榔;SSR標記;聚類分析;遺傳多樣性
中圖分類號:S792.91 ? ? 文獻標識碼:A
Abstract: Arecanut (Areca catechu L.) is an important tropical economic and social importance plant in Hainan Prov-ince. It has high medicinal value and is listed as the first of the four south China medicinal plants. Areca is mostly planted in underdeveloped areas, and the basic research is weak. The germplasm diversity of areca from Hainan remains unclear due to lack of systemic research of germplasm. We analyzed the genetic diversity among 58 arecanut accessions from Hainan by SSR molecular marker technology to clarify the genetic relationship and provide a theoretical basis for the hybrid breeding and functional gene mining. The results showed that 32 primers with high polymorphism and clear bands were screened out from 500 SSR primers. A total of 79 alleles were detected from the 58 arecanut germplasm resources, and there were 2-5 loci per SSR, with an average of 2.4688 alleles at each marker, and the number of effective allele was accounted for 58.92% of the observed one. The range of PIC was 0.0169-0.5969, with an average of 0.2254. The Shannon index (I) of all primers ranged from 0.0496 to 1.2552, with an average of 0.4496. Neis genetic diversity index (Nei) was ranged from 0.0171 to 0.6492, with an average of 0.2596. The genetic diversity of alleles detected by SSR primers was low. UPGMA Cluster analysis showed that the genetic diversity of group V was the richest. Genetic analysis showed that the 58 arecanut were separated from the effect of regional isolation. The conclusion of frequent exchanges among different regions of arecanut in Hainan and lower genetic diversity in Hainan cultivars would provide theoretical guidance for the selection of parents in the genetic breeding of arecanut, while reference for collecting germplasm to supplement different resources in the future.
Keywords: arecanut; SSR markers; clustering analysis; genetic diversity
DOI: 10.3969/j.issn.1000-2561.2021.05.014
檳榔(Areca catechu L.)是棕櫚科檳榔屬多年生常綠植物,原產于馬來西亞,我國主要栽培于海南、云南以及臺灣等地,其中海南產量占全國總產的99%(未計臺灣省)[1]。檳榔是海南重要的熱帶特色經濟作物,具有很高的藥用價值,被列為“四大南藥”(檳榔、砂仁、益智、巴戟)之首。含有多種人體所需的營養元素和有益物質,檳榔原果的主要成分為酚類、多糖、脂肪、粗纖維和生物堿,檳榔種子總生物堿含量達0.3%~ 0.6%,主要為檳榔堿,并含有少量檳榔次堿,是驅蟲的有效成分,還具有消積導滯、行氣利水、降血壓的功能[2-3]。
我國檳榔栽培歷史悠久,尤其是在海南,形成了一些各具特色的地方品種,不同類型的檳榔在產量、品質、果實形狀等方面存在很大差異[4],但是檳榔種質資源基礎研究相對薄弱,對種質資源的多樣性缺乏系統的比較和研究,存在種質混雜,同名異物或者同物異名等情況[1]。隨著檳榔產業規模的不斷擴大,農戶在引種時偏向于栽培商品喜好型品種,栽培品種單一,一些特有種質正在逐步消失,遺傳基礎可能存在逐年變窄的趨勢。種質資源的收集、保存和鑒定評價對作物新品種選育具有重要作用,種質資源遺傳多樣性是檳榔遺傳育種的基礎和前提,因此,對來源于不同生態區域的檳榔種質資源開展系統遺傳多樣性研究,對檳榔種質資源評價和創新利用具有重要意義。
DNA分子標記目前廣泛應用于作物種質資源遺傳多樣性研究,其中SSR標記具有共顯性、多態性豐富、可重復性好等優點,已被廣泛應用于水稻[5]、小麥[6]、甘薯[7]、綠豆[8]、新麥草[9]等大田作物及牧草,以及椰子[10-11]、油棕[12]等棕櫚科植物的遺傳多樣性評價研究。我國檳榔種質資源的遺傳多樣性研究較少,任軍方[13]利用ISSR標記對海南保亭的檳榔栽培居群進行遺傳多樣性分析,結果顯示海南保亭的檳榔多樣性水平較低。
本研究調查收集海南不同地區、不同果形的58份栽培檳榔資源,利用SSR分子標記技術進行海南栽培檳榔遺傳多樣性研究,明確目前海南栽培檳榔的遺傳基礎,闡明不同材料間的親緣關系,為我國檳榔種質資源鑒定評價、遺傳育種與改良奠定基礎。
1 ?材料與方法
1.1 ?材料
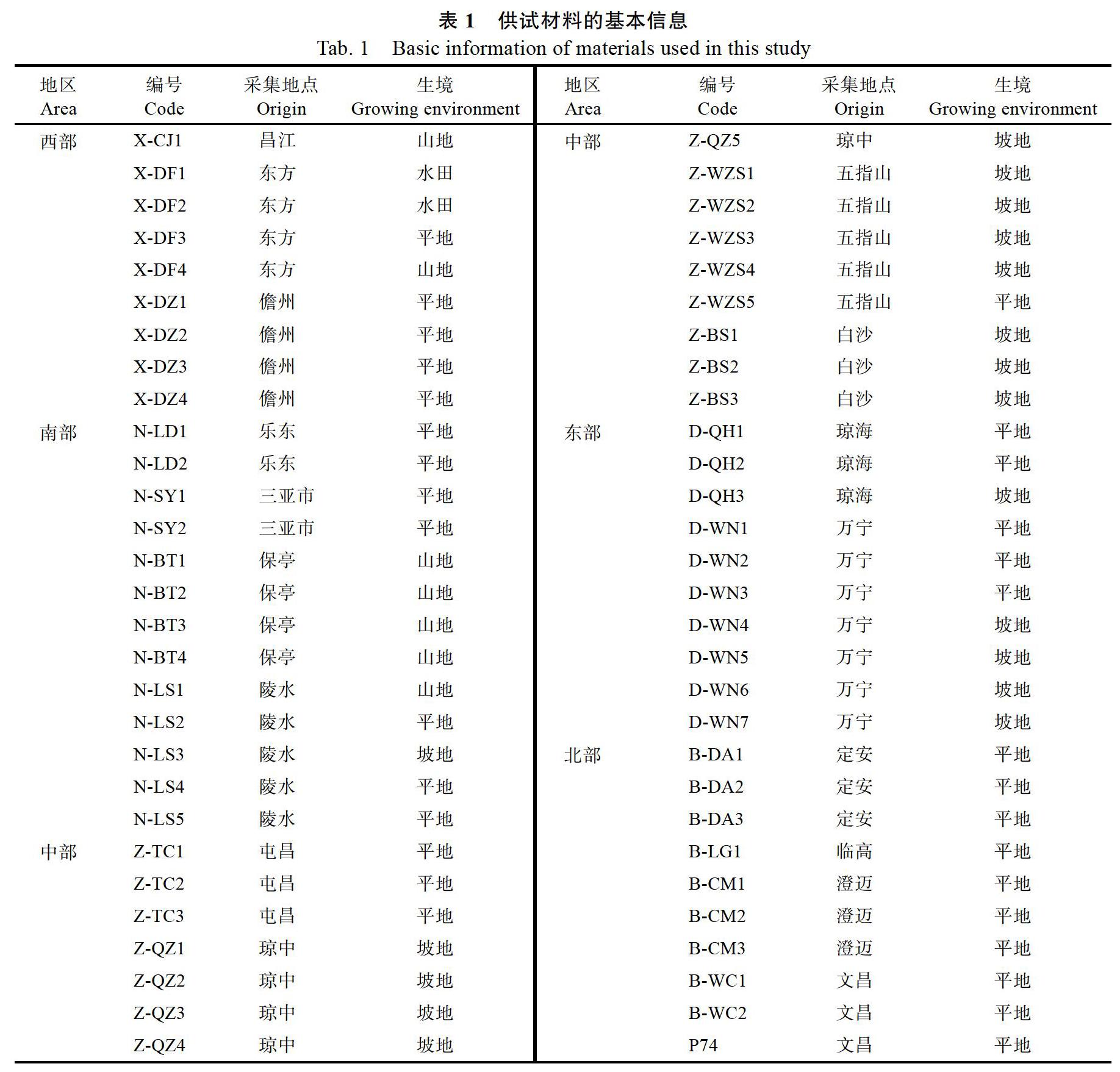
試驗材料共58份,來自海南省各市縣栽培檳榔園,選取檳榔樹株型、果型中有差異的作為試驗樣品,其中P74為從臺灣引進的檳榔種質資源,栽培在中國熱帶農業科學院椰子研究所檳榔種質資源圃,資源圃內編號為P74,其余樣品編號規則:按照各市縣在海南省的地理位置分為:西部地區(X)、中部地區(Z)、南部地區(N)、北部地區(B)、東部地區(D),加上樣品采集地市縣名稱拼音首字母縮寫,樣品基本信息見表1。
1.2 ?方法
1.2.1 ?DNA的提取 ?采集檳榔成齡植株葉片,裝入自封袋,置于放冰袋的泡沫盒帶回實驗室。葉片剪碎后,放入研缽加液氮磨樣,采用植物基因組DNA快速提取試劑盒[生工生物工程(上海)股份有限公司]提取葉片基因組DNA,風干后,加入100 μL TE緩沖液溶解。吸取1 μL DNA原液,利用NanoDrop 2000微量紫外分光光度計檢測提取DNA質量和濃度,工作液濃度稀釋至20 ng/μL,DNA原液和工作液儲存于?20 ℃冰箱備用。
1.2.2 ?引物篩選 ?利用12份地理來源不同,形態差異較大的材料進行引物篩選,從500多對SSR引物序列中篩選特異性好,條帶清晰的引物32對,用于海南栽培檳榔品種基因型檢測(表2)。本研究用到的SSR引物是利用‘熱研一號檳榔品種的葉片、雌花和果實,通過轉錄組測序技術開發的SSR引物,引物序列由中國熱帶農業科學院椰子研究所楊耀東博士提供,引物由生工生物工程(上海)股份有限公司合成。
1.2.3 ?PCR反應和產物檢測 ?SSR分子標記試驗PCR反應體系參考齊蘭等[14]的方法,PCR反應程序為:94 ℃預變性5 min;94 ℃變性30 s,58 ℃退火復性30 s,72 ℃延伸30 s,33個循環;72 ℃延伸10 min,最后設置4 ℃保存。用已經篩選出的多態性高且穩定性好的SSR引物進行SSR-PCR擴增。擴增產物用8%聚丙烯酰胺凝膠電泳檢測,銀染顯色,條帶清晰后在膠片觀察燈上進行拍照。
1.3 ?數據處理
對照Marker(D 2000)的分子量大小根據條帶有無來記錄跑膠結果,將SSR引物擴增出的DNA片段按照從大到小的順序依次排列,相同大小的分子量遷移處,有帶記為“1”,無帶記為“0”,缺失數據記為“9”,建立0-1形式的二進制數字矩陣,通過DataFormater軟件將0-1數據轉換成Popgen、Powermarker等軟件所需的數據模式[15]。
采用Popgene 1.32軟件[16]計算檳榔種質資源的觀測等位基因數(Na)、有效等位基因數(Ne)、期望雜合度(He)和觀測雜合度(Ho)、Shannons信息指數(I)和Neis遺傳多樣性指數(Nei)等遺傳參數。并計算分組群體間的遺傳多樣性和遺傳距離;利用Powermarker V3.25軟件[17]計算每對引物的多態性信息含量(polymorphic information content, PIC);采用Ntsys 2.11軟件[18],利用Clustering的SAHN程序中的非加權類平均法(UPGMA)進行聚類分析,通過Tree-plot程序生成遺傳聚類圖。
2 ?結果與分析
2.1 ?SSR標記多態性分析
檳榔DNA提取原液通過微量紫外分光光度計檢測其濃度,各檢測樣品DNA濃度范圍為610~1053 ng/μL,OD260/280比值介于1.8~2.0之間,說明提取樣品DNA的純度高,滿足SSR-PCR擴增要求。
利用篩選的32對SSR引物,對58份檳榔種質資源進行基因型檢測,結果顯示,32對引物共檢測到79個等位基因,每個SSR位點2~5個,平均2.4688個,其中有效等位變異占觀測等位變異的58.92%。觀測雜合度(Ho)變幅為0.0000~ 0.7586,平均值為0.1886,說明所檢測等位基因位點雜合度稍低;期望雜合度(He)變幅為0.0172~0.6549,平均值為0.2618,PIC值變幅為0.0169~0.5969,平均值為0.2254,中度及以上多態性信息位點(PIC>0.25)有13個。Shannons信息指數(I)的范圍為0.0496~1.2552,平均為0.4496;Neis遺傳多樣性指數(Nei)變幅為0.0171~0.6492,平均值為0.2596,說明SSR引物在供試樣品中檢測等位基因遺傳多樣性偏低(表3)。
2.2 ?不同檳榔種質資源遺傳聚類分析
利用SSR標記檢測基因型數據進行聚類分析,構建親緣關系聚類圖(圖1)。從聚類分析樹狀圖可以看出,遺傳相似系數矩陣(GS)變化范圍為0.63~0.94。在遺傳相似系數為0.70處可以將海南栽培檳榔與臺灣種檳榔區分開。在遺傳相似系數0.82處(圖1中參考線所示),根據離散程度,將供試樣本分為5個組,其中第I組劃分為2個亞組,I-1和I-2組,將未聚類在一起的與其他樣品親緣關系較遠的4份材料劃為第Ⅴ組,分別是來自中部瓊中、五指山,北部定安以及1份臺灣種質。
第I組分為2個亞組,I-1組主要包括中部、北部和東部的材料,其中還有3份來自南部陵水的材料。亞組I-2組的材料來源相對分散,大多數是來自西部和中部地區的樣本,該組的材料來源相對復雜,占供試材料的2/3,基本覆蓋了海南島各地的樣品。其中有幾份來自不同地區的樣本
遺傳距離很接近,遺傳相似系數達到0.95,推測這些材料可能來自相同的親本,說明海南栽培檳榔在不同地區的種苗交流很頻繁。第Ⅱ組和第Ⅲ組包含的材料相對分散,無明顯的區域性劃分,第IV組的3份材料來自南部的陵水和樂東。第V組的材料1份來自臺灣,另外3份與其他海南本地種資源遺傳差異較大,這4份材料各自成為1個分支,推測可能是早期從周邊東南亞國家引進的種質。
2.3 ?基于SSR標記的檳榔分組群體遺傳多樣性與遺傳距離
根據SSR標記基礎上的遺傳聚類結果,將58份海南栽培檳榔種質資源分為5組,由于第I組
成員較多,明顯分為2個亞組,因此按照6個分組,即I-1、I-2、Ⅱ、Ⅲ、IV和V組,利用Popgene 1.32軟件分析檳榔種質分組群體內的遺傳多樣性參數(表4)。對分組群體SSR標記多態性分析結果顯示,第V組的觀測等位基因數(Na)最高(2.2188),最低為第IV組(1.5312);有效等位基因數(Ne)最高為第V組(1.8001),最低為第Ⅱ組(1.2909);Shannons指數(I)最高的是第V組(0.6106),最低為第Ⅱ組(0.2753);Neis遺傳多樣性指數(Nei)最高為第V組(0.3828),最低為第Ⅱ組(0.1704);觀測雜合度(Ho)和期望雜合度(He)分別為0.1328~0.3047和0.1818~0.4375,均為第V組最高。結果表明,第V組的遺傳多樣性最豐富,其他幾組遺傳多樣性參數值稍低,說明海南本地栽培檳榔群體遺傳多樣性水平偏低。
利用Popgene 1.32軟件計算6個分組群體間的遺傳一致度和遺傳距離,群體I-1和I-2組間的遺傳距離最小(0.0192),遺傳一致性系數最大(0.9810);群體IV和群體V的遺傳距離最大(0.1423),遺傳一致性系數最小(0.8673);總體來講不同分組群體的遺傳距離差異不大(表5),表明海南檳榔不同分組群體遺傳差異較小,需要引進國外種質資源以拓寬其遺傳基礎。遺傳一致度和遺傳距離分別是從相同和相反方面度量群體間遺傳關系的遠近,遺傳距離系數大,相應的遺傳一致性系數小,材料間的親緣關系比較遠;因此開展雜交育種時建議選擇親緣關系遠的組合,更有利于產生雜種優勢。
3 ?討論
檳榔是熱帶亞熱帶地區一種重要的經濟作物,栽培地區多為不發達地區,基礎研究薄弱。印度是最早對檳榔開展系統性研究的國家,自1957年開始先后從中國等世界各地收集檳榔種質,已經達到117份,保存在印度的維塔檳榔實驗站[19]。國內外已開展檳榔種質資源遺傳多樣性的分析研究,劉慧娟[20]利用10對RAPD標記對海南20個檳榔主要種植縣市的74份樣品進行聚類分析,在遺傳相似系數為0.78左右檳榔群體被分為2組,第1組包括印度、柬埔寨樣品,還有部分中國云南與海南的樣品等,第2組樣品包括來自中國云南和海南樣品,說明海南栽培檳榔與周邊東南亞國家存在種苗交換現象。任軍方[13]利用ISSR標記對海南保亭的檳榔居群進行遺傳多樣性分析,結果顯示保亭檳榔12個居群總的遺傳多樣性Neis指數為0.2878,表明海南保亭檳榔的多樣性水平比較低,可能是由于采樣區域太小。臺灣學者Hu等[21]開發了9對SSR引物,對臺灣屏東縣的36份檳榔材料進行了群體遺傳分析,預期雜合度和觀測雜合度分別為0.71~0.94和0.00~0.88。Sankaran等[22]利用11對RAPD標記對10份印度檳榔種質材料進行遺傳多樣性分析,PIC值范圍為0.17~0.60,UPGMA聚類分析顯示野生種檳榔與栽培種檳榔遺傳差異大。印度農作物研究中心(Central Plantation Crops Research Institute, CPCRI)的Bharath等[23]利用RAPD標記對60份檳榔種質資源進行遺傳多樣性分析,遺傳相似系數介于0.68~0.93之間,印度本地收集材料聚類在一起,其他國家收集資源聚類在一起,聚類分析結果與地理來源有顯著相關性。RAPD標記技術的一個缺點是實驗重復性差,反應條件的變化常常導致擴增結果帶形的變化,結果可靠性較低[20]。而SSR標記具有重復性高,可靠性強且品種間多態性豐富等特點。Ramaswamy等[24]利用檳榔葉片開展轉錄組測序,在檳榔基因組上鑒定到6853個含SSR位點的區域,可開發更多的SSR標記用于檳榔種質資源研究。
本研究篩選了32對多態性SSR引物,在58份栽培檳榔材料中共檢測到79個等位基因,每個SSR位點2~5個,平均2.4688個。PIC值變幅為0.0169~0.5969,平均值為0.2254。Shannons信息指數(I)范圍為0.0496~1.2552,平均為0.4496;Neis遺傳多樣性指數平均值為0.2596,說明本研究所用SSR引物在供試樣品中檢測等位基因遺傳多樣性偏低,需要更多的SSR標記來評估更多的遺傳資源。
根據SSR分子標記進行UPGMA遺傳聚類分析,從聚類分析圖可以看出,遺傳相似系數矩陣(GS)變化范圍為0.63~0.94。在遺傳相似系數為0.70處可以將海南栽培檳榔與臺灣檳榔種區分開,表明海南本地檳榔與臺灣檳榔種的遺傳差異較大。第V組的材料1份來自臺灣,另外3份與其他海南本地種資源遺傳距離較遠,該組的Shannons信息指數和Neis遺傳多樣性指數均為最高,表明該組樣品間遺傳差異較大。海南在20世紀90年代引種過東南亞部分國家或地區的檳榔品種,后期由于果實品質不能滿足市場需求,在海南的種植面積越來越少,本次采集的這3份檳榔材料是否為從國外引種遺留下來的資源有待于進一步驗證。遺傳分析結果顯示海南栽培檳榔無明顯的地域聚類,說明海南島檳榔栽培種苗交流比較頻繁,且海南栽培檳榔遺傳基礎相對狹窄,其原因可能是農民選種時喜歡商品性較好的品種,主栽優良品種由于長時間的人工馴化、不同地區間資源共享,導致遺傳多樣性水平降低。因此,在今后的育種工作中應積極引進國內外的檳榔種質資源以拓寬其遺傳背景。本研究明確了海南栽培檳榔品種的遺傳多樣性差異,為檳榔種質資源保存利用及遺傳育種、改良、育種親本選擇提供理論指導,也為檳榔種質資源保護和今后引進種質增補差異資源提供參考。
參考文獻
[1] 晏小霞, 王祝年, 王建榮. 檳榔種質資源研究概況[J]. 中國熱帶農業, 2008(5): 34-36.
[2] 鄭錦星, 李忠海, 袁列江, 等. 檳榔生理效應研究進展[J]. 食品科技, 2006(9): 302-305.
[3] 張春江, 呂飛杰, 陶海騰. 檳榔活性成分及其功能作用的研究進展[J]. 中國食物與營養, 2008(6): 50-53.
[4] 黃麗云, 劉立云, 李 ?艷, 等. 海南主栽檳榔品種鮮果性狀評價[J]. 熱帶作物學報, 2014, 35(2): 313-316.
[5] 從夕漢, 施伏芝, 阮新民, 等. 東南亞62個秈型水稻親本SSR遺傳多樣性分析[J]. 核農學報, 2016, 30(5): 859-868.
[6] 李正玲, 胡 ?琳, 王會偉, 等. 河南省同名小麥地方品種SSR遺傳多樣性分析[J]. 麥類作物學報, 2016, 36(5): 564-570.
[7] 聶立圓, 李愛賢, 秦 ?楨, 等. 基于SSR標記的132份甘薯種質指紋圖譜的構建及遺傳多樣性分析[J]. 植物遺傳資源學報, 2018, 19(5): 904-911.
[8] Chen H L, Qiao L, Wang L X, et al. Assessment of genetic diversity and population structure of mungbean (Vigna ra-diata) germplasm using EST-based and genomic SSR mark-ers[J]. Gene, 2015, 566(2): 175-183.
[9] 張 ?晨, 云 ?嵐, 李 ?珍, 等. 新麥草種質的SSR遺傳多樣性及群體結構分析[J]. 植物遺傳資源學報, 2019, 20(1): 48-59.
[10] Xia W, Xiao Y, Liu Z, et al. Development of gene-based simple sequence repeat markers for association analysis in Cocos nucifera[J]. Molecular Breeding, 2014, 34(2): 525-535.
[11] Xiao Y, Luo Y, Yang Y D, et al. Development of microsa-tellite markers in Cocos nucifera and their application in evaluating the level of genetic diversity of Cocos nucifera[J]. Plant Omics, 2013, 6(3): 193 -200.
[12] 周麗霞, 吳 ?翼, 肖 ?勇. 基于SSR分子標記的油棕遺傳多樣性分析[J]. 南方農業學報, 2017, 48(2): 216-221.
[13] 任軍方. 利用ISSR標記對海南保亭檳榔遺傳多樣性的研究[D]. 海口: 海南大學, 2010.
[14] 齊 ?蘭, 黃麗云, 王 ?澤, 等. 正交設計優化檳榔SSR-PCR反應體系及引物篩選[J]. 分子植物育種, 2019, 17(4): 1264-1269.
[15] 樊文強, 蓋紅梅, 孫 ?鑫, 等. SSR數據格式轉換軟件DataFormater[J]. 分子植物育種, 2016, 14(1): 265-270.
[16] Yeh F C, Boyle T J. Population genetic analysis of co-dominant and dominant markers and quantitative traits[J]. Belgian Journal of Botany, 1997, 129: 157.
[17] Liu K, Muse S V. Powermarker: An integrated analysis environment for genetic marker data[J]. Bioinformatics, 2005, 21(9): 2128-2129.
[18] Rohlf F J. NTSYS-pc: Numerical taxonomy and multi-variate analysis system[M]. New York: Exeter Software, Setauket, 1992.
[19] Balasimha D, Rajagopal V. Arecanut[M]. Kasaragod: COCRI, 2004.
[20] 劉慧娟. 海南檳榔種質遺傳多樣性與檳榔黃化病發生關系研究[D]. 海口: 海南大學, 2010.
[21] Hu C H, Huang C, Hung K H. Isolation and characterization of polymorphic microsatellite loci from Areca catechu (Arecaceae) using PCR-based isolation of microsatellite arrays[J]. Molecular Ecology Resources, 2009, 9(2): 658-660.
[22] Sankaran M, Chandrasekar P, Singh D R, et al. Assessment of genetic diversity among arecanut accessions by using RAPD markers[J]. Indian Journal of Horticulture, 2013, 70(3): 428-430.
[23] Bharath B G, Ananda K S, Rijith J, et al. Studies on genetic relationships and diversity in arecanut (Areca catechu L.) germplasm utilizing RAPD markers[J]. Journal of Plantation Crops, 2015, 43(2): 117-125.
[24] Ramaswamy M, Smita N, Naganeeswaranb A, et al. Tran-scriptome sequencing and de novo assembly in arecanut, Areca catechu L elucidates the secondary metabolite path-way genes[J]. Biotechnology Reports, 2018, 17: 63-69.
責任編輯:謝龍蓮
