版納植物園龍腦香科植物遷地保護成效與應用探討
2021-07-19 02:14:25肖云學劉光裕
廣西植物 2021年5期
肖云學 劉光裕

摘 要: ?龍腦香科植物廣泛分布于熱帶亞洲,被認為是亞洲熱帶雨林的標志性物種。中國科學院西雙版納熱帶植物園從建園開始就注重龍腦香科植物的研究、收集與保護,并建立了龍腦香科植物專類園。經過幾十年收集,現已成為我國龍腦香科植物重要的保育基地。該文結合歷年的引種保育、物候及生長量觀測等資料,系統整理了版納植物園在龍腦香科植物引種保育和研究利用的工作。結果表明:自建園以來,版納植物園共收集了7屬34種龍腦香科植物,國內的有11種,國外的有28種。國內引種種類長勢良好,除廣西青梅外均能開花結果。國外種類開花結果的有13種。此外,版納植物園部分龍腦香科植物種子生物學特性、植株抗寒及抗病蟲能力的分析,也說明版納植物園的龍腦香科植物的遷地保護是成功的。該文還結合新的研究成果,對龍腦香科植物在造林、生態恢復等方面的應用進行了探討,以期為進一步加強該科植物的引種、保育及研發等工作提供參考。
關鍵詞: 龍腦香科, 遷地保護, 專類園, 物候, 生態恢復
中圖分類號: ?Q948
文獻標識碼: ?A
文章編號: ?1000-3142(2021)05-0843-10
Abstract: ?Dipterocarpaceae plants are typical lowland tropical trees, and are usually regarded as the flagship species of Asia tropical rainforest. Xishuangbanna Tropical Botanical Garden (XTBG) is one most important botanical garden focus on tropical plant collection, conservation and research since 1959. This paper reviews Dipterocarpaceae collection history, research and utilization of at XTBG based on the phenological observation, mass growth, and adaptability. The results showed that 34 species of Dipterocarpaceae from 7 genera had been collected in the garden, among which 11 species from China and 28 species abroad. All the species have flowers and fruit collected from China, except for ?Vatica guangxiensis. There are 13 species that have flowers and fruits collected from overseas. Integrated with seed germination traits, cold and disease resistance data, we concluded that XTBG Dipterocarpaceae plants ex-situ conservation is successful, which may afford a better guide for Dipterocarpaceae plants conservation in the future.
Key words: Dipterocarpaceae, ex-situ conservation, living collection, phenology, ecological restoration
龍腦香科(Dipterocarpaceae)植物是典型的泛熱帶分布科,為東南亞低地濕性雨林的優勢科(李錫文等,2002)。東南亞擁有地球上第三大熱帶雨林,擁有豐富的物種,是全球生物多樣性的熱點區域之一(Mittermeier et al., 2005)。據調查,全世界約有13個屬470種分布在東南亞 (Appanah & Turnbull,1998),且大都樹型高大,是群落中的上層樹種,因此大部分的東南亞熱帶雨林被認為是龍腦香林(Primack & Corlett, 2005)。龍腦香科的木材具有很高的利用價值,在南亞、東南亞許多國家的國際熱帶木材市場中扮演著重要的角色,出口的各類木材中龍腦香科植物約占80% (FAO, 2007)。自20世紀70年代以來,隨著人口的增長,對農業用地的需求也增加,許多熱帶雨林被開墾為農業用地,如在東南亞,油棕的快速擴張,人類活動的破壞,導致雨林大量消失,龍腦香科植物也遭到嚴重的破壞 (Koh et al., 2011)。如今,亞洲的龍腦香科植物的分布、生態及持久性均面臨人為干擾和氣候變化等諸多挑戰(Saatchi et al., 2011)。
中國南部的少部分地區有龍腦香科植物的分布,國產龍腦香科有5屬12種(Li et al.,2007)。中國龍腦香科植物分布的區域,同樣面臨著被砍伐用于木材、森林破碎化等很多問題,導致龍腦香科植物的生存空間縮小,如坡壘(Hopea hainanensis)及青梅(Vatica mangachapoi)等均已經面臨母株減少的困境(鐘義,1983),加之龍腦香科植物自身繁育的缺陷,使得中國的龍腦香科植物大部分種類處于瀕危狀態(孟令增和許再富,2005)。中國龍腦香科植物亟需系統地保護規劃和實踐。此外,龍腦香科植物作為熱帶雨林的關鍵物種,對生態系統的健康及可持續發展發揮著重要作用,這些物種的消失又可以引起其他物種的興衰(McNeely et al., 1990 ; 張知彬, 1994)。因此,保護好龍腦香科植物對熱帶雨林的生態保護有重要意義。
在中國,能夠野外活體保護龍腦香科植物的地方極少,西雙版納是最為重要的一個區域,也是龍腦香科植物保護和利用矛盾最突出的地方。據統計,云南每年從東南亞進口龍腦香科木材約30種(羅良才,2008),中國市場對龍腦香科植物木材需求旺盛,而且有研究證明龍腦香科植物是最適宜熱帶地區生長的木材樹種(姜笑梅等,2010)。假如龍腦香科植物保護能與木材種植無縫對接,能在中國熱帶地區的橡膠林推廣種植龍腦香科樹種,開展替代膠林種植,不僅可以修復西雙版納的荒地、破碎化的森林,而且可以發動群眾更好地保護西雙版納的龍腦香科植物。龍腦香科植物屬于珍稀瀕危植物,且木材材質優良,在植物保護和資源開發上潛力巨大,是中國植物保護事業中值得探索的一種模式。因此,掌握其相關生物學特性和造林技術等基礎資料,是保護和推廣種植能否成功的關鍵因素。
中國科學院西雙版納熱帶植物園(以下稱“版納植物園”)歷來重視龍腦香科植物的引種保育工作,建有約1.7 hm2龍腦香科專類園,收集、保存了國內外的34種龍腦香科植物,并開展了引種栽培、物候觀測、生理生態、遺傳多樣性等研究,成為我國研究龍腦香科植物研究的重要基地。通過一系列研究的開展,掌握了龍腦香科植物遷地栽培的基礎資料,但對于龍腦香科植物種子的長期保存尚未攻克,對造林技術也缺乏足夠的經驗,遷地保護是否成功也缺乏科學的評估報告。本文結合歷年的引種保育情況、物候和生長量等資料以及有關研究結果,系統整理了版納植物園龍腦香科植物收集的種類、園區分布,以及研究利用概況,分析了引種龍腦香科植物在園內的生長適應性,提出引種保育需要努力的方向,為提升龍腦香科植物的引種保育能力及后續的研發推廣種植提供參考。
1 版納植物園龍腦香科植物引種概況
1.1 版納植物園歷年引種的龍腦香科植物
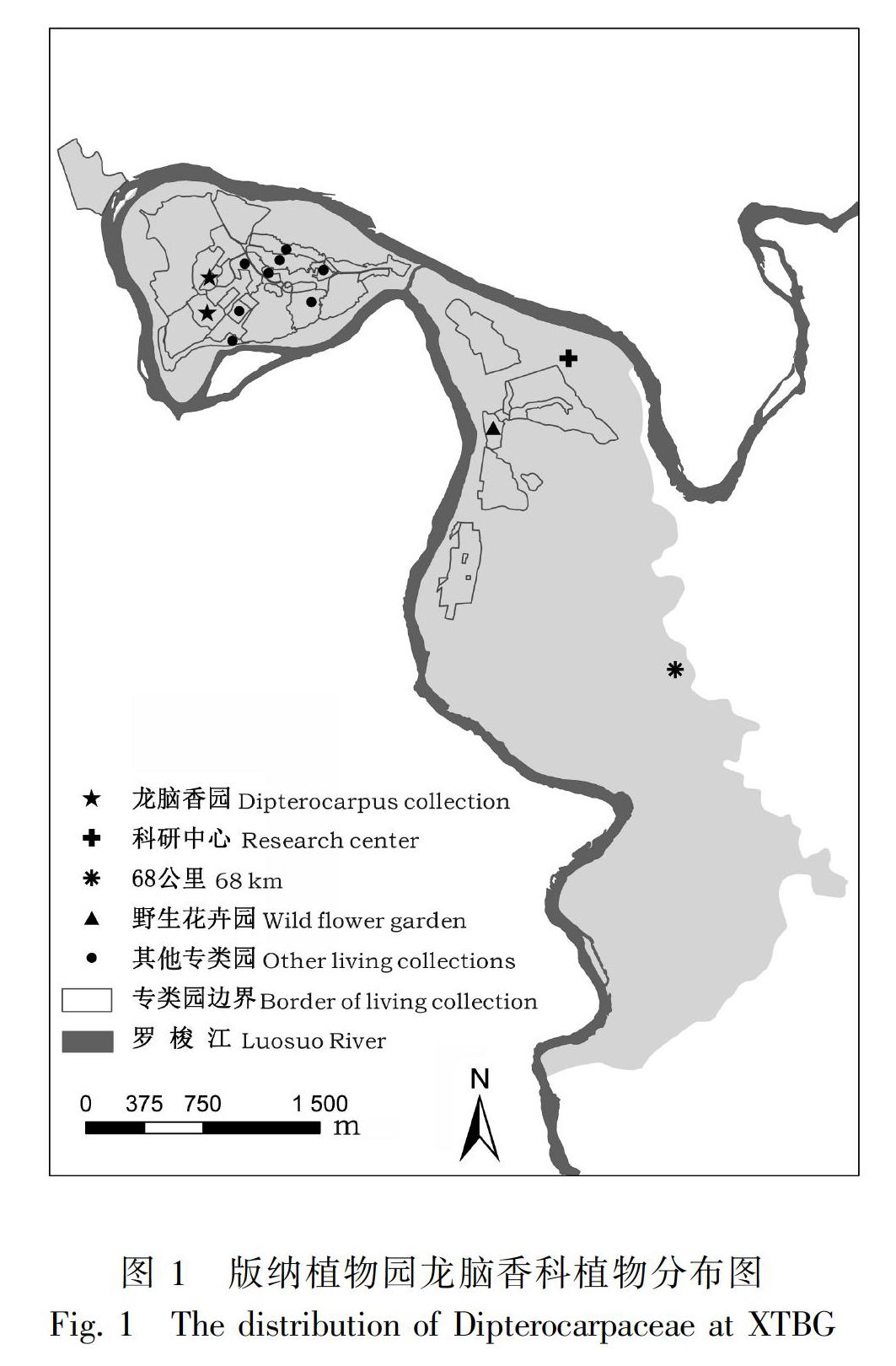
從1959年建園開始,版納植物園就注重龍腦香科植物的收集、引種與保育工作。目前,園內共收集保存了龍腦香科植物7屬34種,主要分布在龍腦香園專類園區中,形成了以龍腦香園、新科研中心植物遷地保護區、野生花卉區及68公里(國道G219景洪至勐醒方向的68 km處)共4個主要分布區(圖1),有些種類零星分散于各專類園區中。龍腦香科植物的引種主要經歷3個階段,此期間共引種7屬22種,詳見表1。
前兩個階段的引種是在建園初期為響應國家植物資源開發利用的號召,以熱帶植物資源開發為目的,有目的地從國內外引進相應的樹種,如羯布羅香(Dipterocarpus turbinatus)、纏結龍腦香(D. intricatus)、望天樹(Parashorea chinensis)等,其目的均以木材開發為主;第三階段是結合版納植物園實施《熱帶植物專類園景觀優化項目》,對龍腦香科等亞洲熱帶典型科屬植物進行收集保護,以熱帶龍腦香科植物資源保育為核心目的,基本引種了中國本土的龍腦香科植物,并調整原有密集植物的空間布局,使園區的規劃、植物種植更加科學合理,并兼顧了遺傳多樣性保育。
1.2 版納植物園龍腦香科引種來源地分析
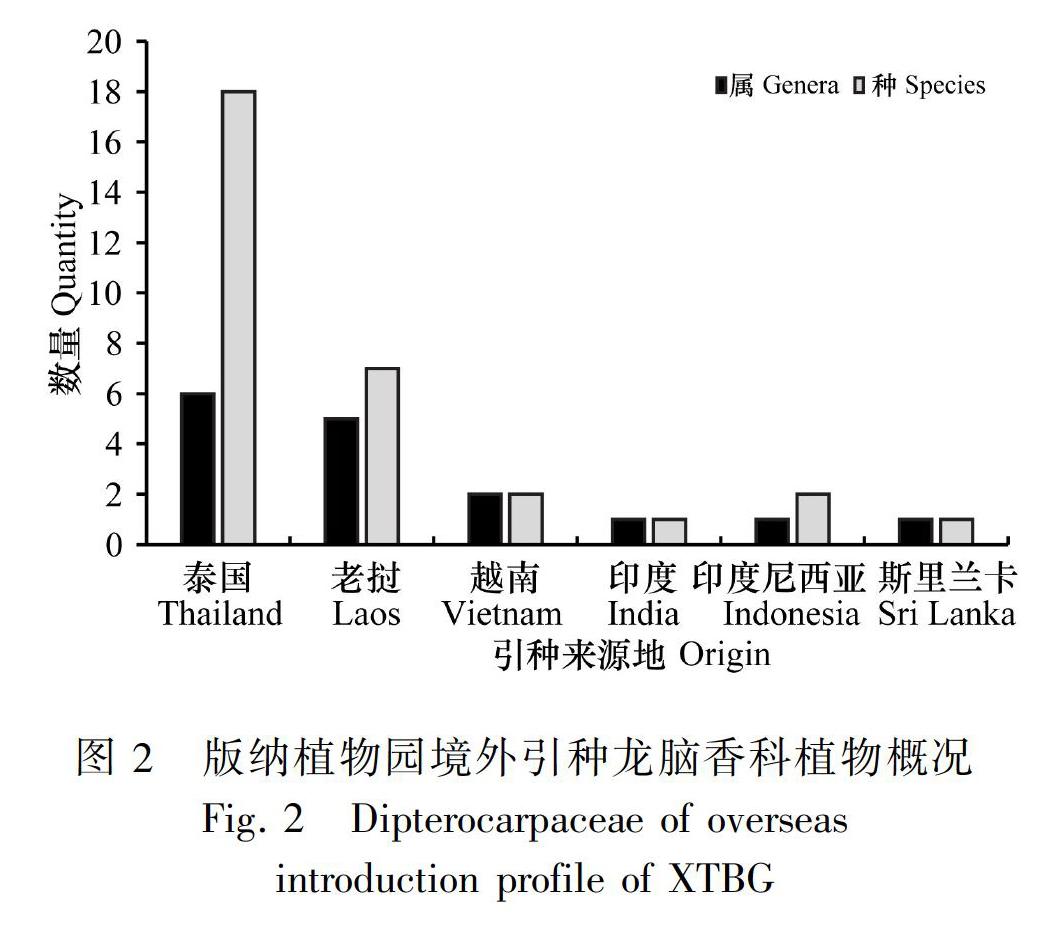
版納植物園內保育的龍腦香科包括了我國除西藏青梅(Vatica lanceifolia)、西藏坡壘(Hopea shingkeng)外的所有國產種,種源地涵蓋了除西藏之外的所有產地。國外的龍腦香科植物主要來自東南亞的泰國、老撾等地,以從泰國、老撾引種的種、屬數量最多,分別為泰國的6屬18種,老撾的5屬7種,其余國家引種較少。與龍腦香科植物的世界分布種類相比較,引種的種、屬偏少,來源地較為單一。分析歷年的引種資料,境外引種詳見圖2。
2 版納植物園引種龍腦香科植物
的生長適應性分析
2.1 引種龍腦香科植物在引種地的生長適應性
版納植物園內保存7屬34種的龍腦香科植物均引自國內外的不同區域,經過多年的精心照料,所有引種的龍腦香科植物在版納植物園內長勢良好,詳見表2。
由表2可知,引種的龍腦香科植物以娑羅雙屬(Shorea)、龍腦香屬(Dipterocarpus)及青梅屬(Vatica)的種類最多,分別為10種、9種、7種,占了種數的76.47%。引自國內的廣西青梅(V. guangxiensis)至今尚無花果,望天樹有花無果外,其余種類均有花果。引自國外的龍腦香科植物有6屬28種,有5種與國產種類相同,其中開花、結果的有13種,不及引種總數的一半。引自國內或者國外的種類,無論開花結果與否,長勢均很好,說明版納植物園是適合龍腦香科植物生長的。
2.2 引種的龍腦香科在引種地的抗逆性
植物從一個生境到另一個生境, 生境中的溫濕度、有害生物的改變會對植物的生長會造成一定的影響。龍腦香科植物作為東南亞熱帶植物區系的一個優勢科,具有較強的熱帶性,種源地的泰國、老撾、越南、斯里蘭卡、印度尼西亞等地與中國云南版納植物園的所在地勐侖鎮相比,是從低緯度到高緯度,低海拔到高海拔,降雨量與熱量高于勐侖鎮。從1959年至今,勐侖地區遭遇過三次大的寒流,第一次是1973年12月下旬至1974年1月上旬,版納植物園的最低溫度是3 ℃;第二次是1999年年底至2000年初,出現百年不遇寒冬,最低溫度為2.2 ℃(許再富和禹平華,1982;張玲等,2001);第三次是2013年12月底至2014年1月初,出現夜間最低溫度2 ℃。應對這些低溫冷害天氣,園內栽培的龍腦香科植物均未表現出明顯的傷害現象,長勢良好。說明版納植物園遷地栽培的龍腦香科植物具有良好的抗寒性。
有害生物是影響植物生長的又一重要因素,植物生境的改變也會影響其對有害生物的抵抗力,植物對有害生物的抗逆性也是植物適應環境的表現。栽植于版納植物園內的龍腦香科植物均是遷地栽培,據筆者多年的觀察以及查閱歷年的檔案資料,目前在引種地的龍腦香林尚無重大病蟲害發生,雖有白蟻、卷葉蛾、大灰象、桑寄生等有害生物危害,但這些有害生物只危害龍腦香科的少部分葉片或枝干,對其生長不構成威脅,說明龍腦香科植物具有良好的抗病蟲能力,在引種地能很好地生長。
2.3 龍腦香科植物種子的萌發與含水量的關系
“種子到種子”是評價植物遷地保護獲得成功的最起碼標準(許再富,1998)。龍腦香科植物的種子通常較大,且不具胚乳,油脂含量低,無休眠期,成熟落地后很快就萌發,種子壽命短,是典型的頑拗性種子(Yan et al., 2007; 李磊等,2016)。因種子無休眠期、不耐存儲、果大、風媒及雨水遠距離傳播受限,成為制約龍腦香引種保育或種群擴散的重要限制因子。有學者對龍腦香科種子的萌發與種子含水量進行過較為系統的研究,結果表明,種子的含水量與種子的萌發率密切相關,隨著種子含水量的降低,種子的萌發率也隨之降低。但不同的種類其萌發率與含水量之間不同(馬信祥和程必強,1990;楊清等,1995;肖來云和普正和,1996;肖來云,1996;楊清等,2008),詳見表3。
由表3可知,不同種類的龍腦香科植物種子萌發的含水量及最適萌發的種子含水量均不同,在自然條件下大都可保存20 d左右,以纏結龍腦香種子在自然條件下貯存的天數最長,可保存50 d以上,而以望天樹保存的天數最短,只有5 d。因此,在野外條件下采集種子時,做好保濕措施在一定程度上可延長種子的壽命,采收后要盡快播種。
3 討論與結論
版納植物園龍腦香科植物遷地保護的實踐證明,版納植物園是適合龍腦香科植物生長的,不僅國產的龍腦香種類能在該園很好地生長,引自泰國、老撾等低緯度、低海拔、高溫區的龍腦香科也能在該園茁壯生長,具有良好的抗寒性、抗病蟲能力。中國的龍腦香科植物通過早期的植物資源開發利用和20世紀80年代后期的定向引種收集得到了有效的遷地保護。此外,在遷地保護的同時還開展了引種栽培、物候觀測、種子萌發特性等方面的基礎研究,積累了較為豐富的科學資料,為進一步開展龍腦香科植物的推廣種植提供了科學依據。目前,國內從事龍腦香科有關研究的單位均到版納植物園采樣及開展相應的研究,龍腦香園作為科研的支撐平臺發揮著越來越重要的作用。
龍腦香科植物在版納植物園的遷地保護獲得成功,除了與當地所處的獨特地理、氣候環境有關外,還與管理人員的精心養護是分不開的。龍腦香科植物在原產地的群落中大都為上層陽性樹種,但其幼苗處于群落下層,光照不足,種間競爭激烈迫使其有較長的蹲苗期。在遷地保護的栽培條件下,對幼苗采取適當遮陰,尤其是在干季對其覆蓋減少灼傷,并輔以清除雜草、肥水管理等措施,解除了龍腦香科幼苗的蹲苗期,促進其生長,這也是該科植物遷地保護獲得成功的重要因素。版納植物園在龍腦香科植物種類的收集、保育及研究方面,也還存在著一些問題需要解決,主要體現在以下幾個方面。
3.1 種群數量不夠,引種潛力大
世界龍腦香科植物有17屬約700種,版納植物園中引種保育了該科植物的7屬34種,屬占比約1/3,但種類還很少。引種來源地的分析結果表明,引種主要來自東南亞的少數幾個國家,對熱帶亞洲分布中心的馬來西亞、印度尼西亞的引種很少甚至沒有,而對熱帶美洲及熱帶非洲的種類尚未進行引種,引種的空間還很大。因該科植物的木質堅硬、樹脂有多種用途,且引種栽植于版納植物園內的種類長勢良好。因此,有必要進行大量引種。這對于提升物種保育能力及生物多樣性保護具有重要意義。
3.2 引種栽培研究較多,木材利用研究方面較少
版納植物園內保存的龍腦香科植物, 在引種栽培、種子繁殖機制等有開展基礎研究,但在木材應用或者其他方面的研究尚屬空白,也沒有將單個種類作為經濟植物進行推廣。建園初期,龍腦香科植物因栽植過密,導致很多種類的莖干增粗受限。作為植物資源戰略儲存庫的版納植物園,對該類植物的引種,不應停留在保育層面,還應在基礎研究、應用研究等方面開展更深入的研究。如研究引種栽培的龍腦香科植物的材質特性、樹脂提香技術、藥用及對生態影響等方面,提升龍腦香科植物的應用價值,真正發揮植物園服務國家、服務社會的功能。
3.3 境外引種的種類長勢良好,但仍有些許種類無花無果
結合歷年的物候觀測資料結果可知來源于國內的龍腦香種類在引種地除廣西青梅外均有開花結果現象,其中的云南娑羅雙(Shorea assamica)隔年有開花現象,且結果率低,有可能是缺少傳粉者,原因有待進一步分析。而來源于國外的種類,長勢良好,未表現出不適的情況,但仍有部分種類無花無果現象,其中的原因較為復雜,可能是生長年限不夠,或許是熱量不足,又或許是微環境的改變,有待于進一步研究和證實。
3.4 引種居群較為單一,遺傳變異研究較少
版納植物園已收集了所有國產龍腦香的種類,但對同一種類不同居群的引種還不充分,如在收集區域方面,尚未引種、保存西藏的種類;在種方面,如東京龍腦香(Dipterocarpus retusus)分布于云南東南部(河口、金平、屏邊、綠春、江城)及西部(盈江那幫壩)等(陶國達等,1990),目前只有引自盈江及綠春,其他地區的尚未引種,河內坡壘(Hopea hongayensis)也存在這一問題。保護植物遺傳基因的多樣性是遷地保護的最終目的(孟令曾和許再富,2005)。自然雜交在植物界非常普遍,有研究表明約有25%的被子植物在其歷史進化歷程中與其近緣種發生了雜交(Mallet, 2007),雜交的結果使物種的遺傳發生了改變,有時還會導致一些稀有居群或物種滅絕(王錚峰和彭少麟,2003)。版納植物園龍腦香園內栽培的龍腦香科植物親緣關系較近,雜交的可能性較高。目前,李巧明和許再富(2001)對版納青梅(Vatica xishuangbannaensis)與廣西青梅、望天樹的遺傳多樣性及居群開展了相關研究,結果發現版納青梅具有較高水平的群體分化,居群內存在一定程度的近交,但居群變異水平很低;望天樹群體內遺傳變異很低,無近交現象。對于其他種類,尤其是同一物種不同來源地及種間一起栽培是否會引起基因交流而導致物種的遺傳發生變異,也缺乏相應的研究。
4 展望
龍腦香科植物是具有重要經濟價值的類群,在我國大都列入保護植物的范疇,引種栽培的國內外種類在版納植物園均生長良好,這為該科植物的遷地保護提供了范例。為進一步提升龍腦香科植物的保育能力及推廣種植,建議從以下幾個方面加強研究。
4.1 合理規劃引種方案,豐富園區的物種
2019年,在版納植物園建園60周年之際召開了國際專家咨詢會,與會專家指出在專類園建設方面,龍腦香園是版納植物園最具特色的專類園,已有一定的研究基礎,后期要加強對龍腦香科植物的引種與研究。因此,結合版納植物園的發展規劃,加大對龍腦香科植物的引種,對國內分布的種類,完善同一種類不同居群的引種,豐富物種的遺傳多樣性,對境外的引種也要盡可能引進不同居群的種類。同時,要擴大龍腦香園的面積,為植物的生長提供足夠的空間。在野外生境考察時,注意對原生境的土壤、伴生物種等資料的采集。對于境外的種類,要了解并掌握其分布區域的環境氣候等因子,結合引種地的生境,確定可引種的種類,并積極創造條件,到境外引種。龍腦香科植物種子大都為頑拗性,在野外引種時以收集小苗最佳,采種時要做好保濕措施。
4.2 改善當前的種植模式,探索群落建園
當前,版納植物園內的龍腦香園以純林營造為主,林下蔭蔽度大,幾乎沒有伴生物種,物種多樣性極低。在野外生境中,該科植物為中上層樹種,且還伴生有其他物種,多個物種間既有競爭又有依賴,彼此共同進化,最終達到一個相對穩定的狀態(張桂萍等,2006)。有學者對云南娑羅雙、版納青梅、望天樹等物種的群落特征進行過系統的研究(朱華,1993;李宏偉,2006),但對群落間物種關聯度的研究較少,探明龍腦香科植物群落間物種的關聯度對建園有重要的參考價值。在引種地,有些種類開花無果,可能是缺乏相應的傳粉者。在遷地栽培時,控制目標物種的數量,樹與樹之間留有足夠的生存空間,適當栽植一些有協同作用的伴生物種,這對于提升龍腦香林內生態系統的穩定性,完善生態系統內的營養循環,減少管理和維護成本具有重要意義。
4.3 夯實龍腦香科的基礎研究,逐步開展應用研究
引種的目的是保育,保育的最終目的是應用。對現有的龍腦香科進一步加強對其生長適應性的研究,掌握繁殖技術和栽培技術,對有果實的種類繼續探索種子的萌發與繁育機制,為進一步的推廣栽培打好基礎。同時,要加強遷地栽培種是否會引起物種遺傳多樣性散失或降低的研究,為遷地保護提供科學的理論指導。開展木材材質、樹脂成分、藥用和香料提取等方面的研究,提升龍腦香科植物的高附加值,為后期的研發提供動力。
4.4 加強龍腦香科植物種子延長保存及造林技術的研究
龍腦香科植物的種子大多為頑拗性種子,不耐失水,不耐儲藏,對于野外采種或種子保存都極為不利。要加強龍腦香種子的延長保存技術的研究,對龍腦香科的種質資源保存有重要意義。另外,在生態植被的恢復過程中對于龍腦香科植物的造林技術,也缺乏足夠的經驗。作為熱帶雨林的關鍵樹種,本身具有很高的經濟價值,研究其造林技術無論是對生態還是當地居民都具有積極的效應。馬來西亞的沙撈月地區,在雨林的恢復過程中就有使用部分龍腦香樹種來作為種植的樹種(Hattori et al., 2013),并積累了豐富的經驗,這為熱帶雨林使用龍腦香樹種作為生態修復及后續的造林、推廣種植提供了案例。
參考文獻:
APPANAH S, TURNBULL JM, 1998. A review of dipterocarps: Taxonomy, ecology, and silviculture [M]. Bogor: CIFOR: 99-114.
FAO, 2007. State of the worlds forests [M]. Food and Agriculture Organisation of the United Nations, Rome: 157.
GUO F, HUANG RF, YU G, 2015. Effects of heat treatment and modified PU coating on the photo-induced discoloration of wood [J]. J Nanjing For Univ (Nat Sci Ed), 39(2): 169-173. [郭飛, 黃榮鳳, 余鋼, 2015. 熱處理和改性 PU 漆對木材光變色性能的影響 [J]. 南京林業大學學報(自然科學版), 39(2): 169-173].
HATTORI D, KENZO T, KENDAWANG JJ, et al., 2013. Rehabilitation of degraded tropical rainforest using dipterocarp trees in Sarawak, Malaysia [J]. Intl J For Res: 1-13.
JIANG XM, YIN YF, LIU B, 2010. Current status, development, and the prospect of wood identification technology [J]. Chin Wood Ind, 24(4): 36-39. [姜笑梅, 殷亞方, 劉波, 2010. 木材樹種識別技術現狀、發展與展望 [J]. 木材工業, 24(4): 36-39.]
KOH LP, JUKKA M, SOO CL, et al., 2011. Remotely sensed evidence of tropical peatland conversion to oil palm [J]. Proc Natl Acad Sci USA, 108: 5127-5132.
LI HW, 2006. A phytocoenological study on Vatica guangxiensis (Dipterocarpaceae) community in Mt.Maocaoshan, Xishuangbanna [J]. Acta Bot Yunnan, 28(2): 162-168. [李宏偉, 2006. 西雙版納茅草山廣西青梅群落學研究 [J]. 云南植物研究, 28 (2): 162-168.]
LI L, MENG ZG, LONG GQ, et al., 2016. Advances on recalcitrant seeds of plants [J]. J Trop ?Subtrop Bot, 24(1): 106-118. [李磊, 孟珍貴, 龍光強, 等, 2016. 植物頑拗性種子研究進展 [J]. 熱帶亞熱帶植物學報, 24(1): 106-118.]
LI QM, XU ZF, 2001. Genetic diversity and population differention of Vatica guangxiensis [J]. Acta Bot Yunnan, 23(2): 201-208. [李巧明, 許再富, 2001. 版納青梅居群的遺傳多樣性和群體分化 [J]. 云南植物研究, 23(2): 201-208.]
LI QM, XU ZF, 2001. Population genetic structure and differentiation of parashorea chinensis (Dipterocarpaceae) [J]. Acta Bot Yunnan, 23(3): 313-320. [李巧明, 許再富, 2011. 龍腦香科植物望天樹的遺傳結構及分化 [J].云南植物研究, 23(3): 313-320.]
LI XW, LI J, ASHTON PS, 2002. Notes on the Dipterocarpaceae of China [J]. Acta Bot Yunnan, 24(4):409-420. [李錫文, 李捷, ASHTON PS, 2002. 中國龍腦香科植物紀要 [J]. 云南植物研究, 24(4): 409-420.]
LI XW, LI J, ASHTON PS, 2007. Flora of China ?[J]. Beijing: Science Press, 13: 10284.
LUO LC,2008. Identification and utilization of imported Dipterocarpaceae wood in Yunnan [J]. J W Chin For Sci, 37(3): 99-107. [羅良才, 2008. 云南進口龍腦香科樹種的木材識別與利用 [J]. 西部林業科學, 37(3): 99-107.]
MA XX, CHENG BQ, 1999. Study on seeds and its germination of 100 tropical plants [J]. Trop Bot Res, 40: 18-30. [馬信祥, 程必強, 1999. 100種熱帶植物種子及發芽研究 [J].熱帶植物研究, 40: 18-30.]
MALLET J, 2007. Hybrid speciation [J]. NAT, 446: 279-283.
MCNEELY JA, MILLER KR, REID WV, 1990. Protecting the worlds biodiversity [M]. Translated by XUE DY, WANG LQ, ZHOU ZJ, et al. Beijing: China Environmental Science Press. [MCNEELY JA, MILLER KR, REID WV, 1990. 保護世界的生物多樣性 ?[M]. 薛達元, 王禮嬙, 周澤江, 等, 譯. 北京:中國環境科學出版社.]
MENG LZ, XU ZF, 2005. The threatened status and ex-situ community conservation approach on Dipterocarpaceae in China [J]. ?Guihaia, 25(1): 8-13. [孟令曾, 許再富, 2005. 中國龍腦香科植物受脅狀況及遷地群落保護探討 [J]. 廣西植物, 25(1): 8-13.]
MITTERMEIER RA, DA FONSECA GAB, RYLANDS AB, et al., 2005. A brief history of biodiversity conservation in Brazil [J]. Conservation Biol, 19(3): 601-607.
PRIMACK RB, CORLETT RT, 2005. Tropical rain forests: An ecological and biogeographical comparison [M]. Blackwell Science, harback, 21.
SAATCHI SS, HARRIS NL, BROWN S, et al., 2011. Benchmark map of forest carbon stocks in tropical regions across three continents [J]. Proc Natl Acad Sci USA, 108: 9899-9904.
TAO GD, 1990. Flora Reipublicae Popularis Sinicae ?[M]. Beijing: Science Press, 50: 113. [陶國達, 1990. 中國植物志 [M]. 北京: 科學出版社, 50: 113.]
WANG ZF, PENG SL, 2003. Plant hybridization and its harmful genetic consequences [J]. Biodivers Sci, 11: ?333-339. ?[王崢峰, ?彭少麟, 2003. 雜交產生的遺傳危害—以植物為例 [J]. 生物多樣性, 11: 333-339.]
XIAO LY, PU ZH,1996. A study on immigration protection rare and endangered plants [J]. Yunnan For Sci Technol, 1: 45-53. [肖來云, 普正和, 1996. 珍稀瀕危植物的遷地保護研究 [J]. 云南林業科技, 1: 45-53.]
XIAO LY,1996. Introduction and cultivation of Dipterocarpus intricatus Dyer [J]. J Plant Resour Environ, 5(1): 63-64. [肖來云, 1996. 纏結龍腦香的引種栽培 [J]. 植物資源與環境, 5(1): 63-64.]
XU ZF, YU PH,1982. The research of the adaptability of the introduced dipterocarps to lower temperature [J]. Acta Bot Yunnan, 4(3): 297-301. [許再富, 禹平華, 1982. 引種的龍腦香科植物對低溫適應性的探討 [J]. 云南植物研究, 4(3): 297-301.]
XU ZF,1998. Principle and methodology of ex situ conservation for rare and endangered plants [M]. Kunming: Yunnan Science and Technology Press. [許再富, 1998. 稀有瀕危植物遷地保護的原理與方法 [M]. 昆明:云南科技出版社.]
YAN XF, CAO M, XU HL, 2007. Effects of desiccation and temperature on the germination of Shorea chinensis (Dipterocarpaceae) seeds [J]. Seed Sci Technol, 35(1): 232-236.
YANG Q, XIAO CF, ZHANG L, et al., 2008. Introduction and cultivation of Dipterocarpus zeylanicus in Xishuangbanna [J]. J N China For Univ, 23(5): 99-102. [楊清, 肖春芬, 張玲, 等, 2008. 錫蘭龍腦香引種栽培研究 [J]. 西北林學院學報, 23(5): 99-102.]
YANG Q, XIAO LY, PU ZH,1995. Introduction and cultivation of Dipterocarpus tuberoulatus [J]. Subtrop Plant Res Comm, 24(2): 41-45. [楊清, 肖來云, 普正和,1995. 小瘤龍腦香的引種栽培 [J]. 亞熱帶植物通訊, 24(2): 41-45.]
ZHANG GP, ZHNG F, RU WM, 2006. Interspecific correlations among dominant populations of ligneous species in Mianshan Mountain of Shanxi [J]. J Chin Ecol, 25(3): 295-298. [張桂萍, 張峰, 茹文明, 2006. 山西綿山植被木本植物優勢種群種間關聯 [J]. 生態學雜志, 25(3): 295-298.]
ZHANG L, XIAO CF, WANG J, 2001. Ex-situ conservation of Hopea chinensis [J]. Guihaia, 21(3): 277-280. [張玲, 肖春芬, 王堅, ?2001. 瀕危植物狹葉坡壘的遷地保護 [J]. 廣西植物, 21(3): ?277-280.]
ZHANG ZB, 1994. Several theoretical bases for biodiversity conservation [M]//Biodiversity committee of the Chinese academy of sciences. Principles and methods of biodiversity. Beijing: China Science and Technology Press: 36-54. [張知彬, 1994. 生物多樣性保護的若干理論基礎 [M]// 中國科學院生物多樣性委員會編.生物多樣性的原理與方法. 北京:中國科學技術出版社: 36-54.]
ZHONG Y, 1983. The succession and protection of plant resources of Hainan Island [J]. Wuhan Bot Res, 1(1): 101-110. [鐘義, 1983. 海南島熱帶植物資源的演變和保護 [J]. 武漢植物學研究, 1(1): 101-110.]
ZHU H, 1993. A phytocoenological study on vatica forest in Xishuangbanna [J]. Guihaia, 13(1): 48-60. ?[朱華, 1993. 西雙版納青梅林的群落學研究 [J]. 廣西植物, 13(1): 48-60.]
ZHU H, WANG H, XIAO WX, 2007. A study on Parashorea chinensis community of tropical rain forest in Gulinqing of Maguan County, SE Yunnan, China [J]. Guihaia, 27 (1):62-70. [朱華, 王洪, 肖文祥, 2007. 滇東南馬關古林箐熱帶雨林望天樹群落的研究 [J]. 廣西植物, 27 (1): 62-70.]
(責任編輯 何永艷)
