種植密度對油菜正反交組合產量與倒伏相關性狀的影響
2021-07-19 09:39:22婁洪祥姬建利劉暑艷尹羽豐周廣生
作物學報 2021年9期
關鍵詞:產量
婁洪祥 姬建利 蒯 婕 汪 波 徐 亮 李 真 劉 芳 黃 威 劉暑艷 尹羽豐 王 晶,* 周廣生
種植密度對油菜正反交組合產量與倒伏相關性狀的影響
婁洪祥1姬建利1蒯 婕1汪 波1徐 亮2李 真1劉 芳3黃 威4劉暑艷5尹羽豐6王 晶1,*周廣生1
1華中農業大學植物科學技術學院 / 農業農村部長江中游作物生理生態與耕作重點實驗室, 湖北武漢 430070;2青海省農林科學院, 青海西寧 810016;3全國農技推廣中心油料處, 北京 100125;4黃岡市農業科學院, 湖北黃岡 438000;5荊州農業科學院, 湖北荊州 434000;6襄陽市農業科學院, 湖北襄陽 441057
高產抗倒不協調是油菜生產中最主要的矛盾, 嚴重制約著油菜生產效益的進一步提高。在高密度、機械化生產模式下, 這種矛盾尤為突出。受母系遺傳的影響, 雜交油菜的正、反交組合在產量與倒伏相關性狀上常表現出明顯差異, 但種植密度對不同雜交組合這些關鍵性狀的影響尚不明確。本研究針對浙油50、華航901及其正交(華航901×浙油50)與反交(浙油50×華航901)組合設置5個種植密度, 考察其產量及倒伏相關性狀并計算其雜種優勢, 研究種植密度對親本及正、反交組合產量與倒伏相關性狀的影響, 為當前高密度直播生產模式下雜交油菜親本的選配提供依據。結果表明, 隨種植密度增加, 親本及正、反交組合的實際產量和經濟系數均呈先增后降的趨勢, 且無論正、反交組合, 其實際產量的動態變化規律均與父本更為接近; 倒伏指數在兩點隨種植密度的變化規律不同, 但無論正、反交組合, 其倒伏指數的動態變化規律均與母本相一致。在西寧試點, 雜交組合產量均具有明顯的平均雜種優勢, 且正交組合比反交組合具有更大的雜種優勢; 正交組合具有一定的抗倒性雜種優勢, 但是反交組合無抗倒性雜種優勢。綜上所述, 高密度直播生產模式下, 雜交油菜親本選配推薦方案以高產品種為父本, 抗倒性強的品種為母本進行組配, 可以獲得高產、抗倒性強且更適宜密植的雜交油菜品種。
油菜; 正反交組合; 種植密度; 產量; 倒伏
油菜(L., AACC, 2=38)是世界四大油料作物之一, 廣泛種植于加拿大、中國、印度、澳大利亞和法國等國家, 近10年來全球種植面積已超過3500萬公頃[1]。我國是世界上第二大油菜生產國, 僅次于加拿大, 常年種植面積超過650萬公頃, 約占全球油菜種植面積的20%[2]。油菜是我國食用植物油市場的第一大油源, 常年供應的菜籽油占國產植物油的47%左右[3]。近年來, 雖然在新品種選育、生產和加工等方面穩步發展, 但與其他大田作物一樣, 油菜產業同樣面臨著產量低、種植密度低、機械化程度較低、肥料施用高和人工成本較高的“三低兩高”等瓶頸問題, 導致其增長乏力[4-5]。眾所周知, 自第一次“綠色革命”至今, 世界三大糧食作物——水稻、玉米和小麥的株型不斷矮化且種植密度不斷提高, 密植已成為提高作物產量的重要栽培措施之一[6-11]。自20世紀90年代至今, 育種家們也在不斷尋找油菜矮桿突變體并鑒定了一些控制油菜矮桿的基因位點, 為油菜“矮化革命”奠定基礎[12]。另一方面, 栽培學家們不斷優化油菜種植密度和其他措施以協調油菜密植、高產與抗倒之間的矛盾[13-14]。研究表明, 在我國長江流域地區合理密植可以改變油菜的個體-群體關系, 平衡個體發育與群體競爭, 提高油菜光能利用率, 從而提高產量并保持抗倒特性[15-16]。高密度種植可能是未來油菜高產栽培的常規措施, 也是目前油菜栽培研究的熱點[17-18], 但是大幅增加種植密度后, 油菜倒伏風險顯著增加, 因此培育并篩選高產、抗倒且耐密的油菜品種迫在眉睫[19-20]。
由于雜種優勢的存在, 作物雜交品種較常規品種通常具有高產、穩產、抗逆等特性[21]。油菜具有很強的雜種優勢, 目前在我國油菜生產上主要使用的品種為雜交品種, 占油菜種植總面積的70%以上, 且推廣面積逐年增加, 并逐漸延伸至飼用油菜和生物能源油菜等領域[22-24]。雜交油菜品種不論在籽粒大小和粒重等產量性狀方面, 還是在株高和分枝數等株型性狀上均表現出較強的雜種優勢[12,25]。值得注意的是, 作物的雜種優勢也受種植密度的影響, 不同種植密度下雜種優勢潛力存在顯著差異[26-28]。另外, 在雜交品種選育過程中, 選擇配合力強的親本非常重要, 同時在不同栽培措施下正、反交組合之間的產量與倒伏相關性狀差異研究不容忽視[29-31]。已有研究表明, 正、反交組合在植物光合作用和內源激素調控上存在一定差異, 從而影響了正、反交組合雜交油菜的產量與品質[32-33]; 在一定程度上, 油菜的籽粒大小和籽粒重量等產量性狀和含油量均存在母系遺傳特征[25,34-36]。因此, 在油菜育種工作中應充分考慮各性狀的遺傳規律, 選配適宜的父母本材料以獲得高產、高油且抗倒的優質油菜雜交品種。例如, 在配制油菜的高光效雜交組合時應盡量選用高值親本做母本, 利用雜種優勢來提高雜種F1代的光合相關性狀[32]。
綜上, 前人關于種植密度對于正、反交油菜組合產量和倒伏相關性狀的相關研究較少。本研究通過分析不同種植密度下油菜正、反交組合的產量和倒伏相關性狀的變化趨勢及其雜種優勢, 研究油菜高產、抗倒、耐密植理論和雜交油菜父母本選配規律以適應機械化生產, 旨在為油菜雜交種親本選配以及雜種優勢利用提供參考。
1 材料與方法
1.1 試驗地點與材料
試驗分別于2016—2017年在湖北武漢及2017年在青海西寧2個試驗點進行, 以2個常規油菜品種浙油50和華航901及其正反交后代為研究對象。浙油50為高產、高含油量品種, 華航901為抗倒性較強的品種。以華航901×浙油50為正交組合, 浙油50×華航901為反交組合, 通過人工去雄、授粉獲得F1代雜交種。2016—2017年武漢試驗點播期為2016年9月29日, 收獲期為2017年5月1日。2017年西寧試驗點的播期為2017年4月5日, 收獲期為2017年8月10日。
1.2 試驗設計
采用完全隨機區組試驗設計, 2個油菜親本及其正反交F1雜種均設置了5個不同的種植密度, 分別為15萬株hm-2(D1)、30萬株hm-2(D2)、45萬株hm-2(D3)、60萬株hm-2(D4)、75萬株hm-2(D5), 各處理均設3個重復, 每個重復廂寬2 m, 溝寬0.2 m, 行距0.25 m, 小區面積20 m2。2016—2017年武漢試驗點的原始土壤含全氮0.9 g kg-1、堿解氮24.27 mg kg-1、速效磷14.08 mg kg-1、速效鉀127.96 mg kg-1、有機質15.77 g kg-1; 2017年西寧試驗點的原始土壤含全氮2.23 g kg-1、堿解氮41.34 mg kg-1, 速效磷170.25 mg kg-1、速效鉀374.51 mg kg-1、有機質36.71 g kg-1。各個小區施肥用量一致, 整地時施用750 kg hm-2復合肥(N∶P2O5∶K2O=15%∶15%∶15%)和7.5 kg hm-2硼肥作基肥, 苗期施用225 kg hm-2尿素(含氮量為46%)作苗肥, 薹期施用150 kg hm-2尿素作薹肥。出苗后間苗, 在四至五葉期根據種植密度定苗。兩點試驗其他田間管理均同常規。
1.3 測定指標與方法
1.3.1 產量與產量構成因子 在油菜角果成熟期,每小區連續取樣10株, 考察單株角果數、每角果粒數、千粒重和單株產量, 以小區實收記實際產量。
經濟系數(harvest index, %) = 實際籽粒產量(g)/實際生物量(g)×100。此處實際籽粒產量為各小區實際收獲油菜籽粒產量, 實際生物量為各小區實際收獲總生物量。
1.3.2 倒伏相關指標 在油菜角果成熟期, 每小區連續取樣10株, 考察株高、有效分枝高度、根頸粗和地上部鮮重和根鮮重等基本性狀, 去除縮頸段后, 將有效分枝以下平均分成4段(從基部往上依次標注為1、2、3和4段), 采用YYD-1莖稈強度測定儀(浙江杭州托普儀器有限公司)測定其抗折力。
倒伏指數(lodging index, cm g g-1) = 高度(cm)×地上部鮮重(g)/抗折力(g)。此處高度與地上部鮮重為對應莖段至植株頂端的高度與鮮重, 抗折力為該段中部10 cm處抗折力。
1.3.3 雜種優勢指數 雜種優勢(heterosis)是指油菜雜種第1代(F1)在株型、產量及抗倒性方面均比親本優越的現象。以上產量與倒伏相關性狀均可計算其雜種優勢。
油菜雜種優勢相關指標計算公式: 雜種優勢指數(heterosis index, %) = F1/MP×100, MP = (P1+P2)/2。式中, F1為雜交種, P1、P2分別為父本和母本, MP為親本平均值。
1.4 數據處理與分析
采用Microsoft Excel 2018記錄與整理數據, 利用IBM SPSS 25.0進行數據統計分析, 使用Origin 2018進行數據可視化和繪圖。采用LSD法(Least Significant Difference, 最小顯著性差異法)進行檢驗來比較處理間差異顯著性, 顯著性水平為0.05。
2 結果與分析
2.1 種植密度對產量性狀的影響
2.1.1 產量與產量構成因子 在兩點試驗中, 隨種植密度的增加, 單株角果數和每角果粒數均呈下降趨勢, 但下降幅度變化由快向緩, 且各品種變化趨勢一致。在武漢試點, 同一密度下正交組合(華航901×浙油50)的單株角果數最多, 顯著多于反交組合(浙油50×華航901) (<0.05) (圖1-A, B); 在西寧試點, 正交組合的平均每角果粒數顯著多于反交組合(<0.05) (圖1-C, D)。另外, 千粒重在兩點試驗中隨密度變化規律不明顯(圖1-E, F)。正、反交組合及其父、母本的實際產量隨種植密度增加均呈現出先上升后下降的趨勢, 但是不同材料最高產量對應的種植密度存在差異。武漢試點條件下, 浙油50、華航901及正交組合的最大實際產量均出現在D3密度下, 分別為3082.7、2717.9和3182.3 kg hm-2, 而反交組合則出現在D4密度下, 為3055.2 kg hm-2。西寧試點條件下, 浙油50及正交組合最大實際產量均出現在D3密度處, 分別達到4138.6 kg hm-2和4573.7 kg hm-2, 而華航901及反交組合最大實際產量均出現在D4密度下, 分別為3963.1 kg hm-2和4422.8 kg hm-2。無論正、反交, 雜交組合實際產量隨密度變化的趨勢均與父本相一致, 盡管在武漢試點反交組合規律不如正交組合明顯(圖2-A, B)。
隨種植密度增加, 生物產量與經濟系數均呈先增加后降低的趨勢, 且各品種兩點試驗變化趨勢一致(圖2-C~F)。以武漢試點為例, 正交組合的經濟系數在D3密度時最高(28.07%); 而反交組合在D4密度時達到最大值(24.75%)。另外, 從正、反交組合及其父、母本的比較分析可知, 在實際產量和生物產量方面, 正交組合與其父本浙油50的具體數值及動態變化規律一致; 而在西寧試點反交組合也與其父本華航901的變化規律更為接近。在經濟系數方面, 兩點試驗中正交組合和其母本華航901的各密度下平均經濟系數分別高于反交組合和其母本浙油50。在各密度下, 雖然浙油50的平均生物量和平均實產均大于華航901, 但浙油50的生物量與華航901相對增幅比大于實產的相對增幅比, 所以華航901的平均經濟系數大于浙油50。正、反交組合規律亦是如此, 所以正交組合的平均經濟系數大于反交組合(圖2-E, F)。方差分析表明, 除單株角果數受品種與密度互作影響不顯著外(>0.05), 年份(點)、品種和密度對產量及產量構成因子的影響均達極顯著水平(<0.01), 且它們的互作影響也達顯著水平(<0.05) (附表1)。
D1、D2、D3、D4和D5分別表示種植密度為1.5×105、3.0×105、4.5×105、6.0×105和7.5×105株hm–2。ZY50、HH901、HH901×ZY50和ZY50×HH901分別代表浙油50、華航901、華航901×浙油50和浙油50×華航901。
D1, D2, D3, D4, and D5 indicate the planting densities of 1.5×105, 3.0×105, 4.5×105, 6.0×105, and 7.5×105plants hm–2, respectively. Zheyou 50, Huahang 901, Huahang 901×Zheyou 50, and Zheyou 50×Huahang 901 are two parents and their reciprocal hybrids, respectively.
縮寫和處理同圖1。Abbreviations and treatments are the same as those given in Fig. 1.
2.1.2 正、反交組合產量性狀的雜種優勢 分析發現, 千粒重與每角果粒數在正反交組合中均沒有表現出明顯的雜種優勢現象(數據略)。在武漢試點, 正交組合(華航901×浙油50)單株角果數的雜種優勢指數顯著高于反交組合(浙油50×華航901) (<0.05), 正交組合的平均雜種優勢指數達到116.8%, 而反交組合(90.2%)低于100%, 無雜種優勢。另外, 在D3密度下正交組合的雜種優勢指數達到最高(128.5%) (圖3-A)。正交組合單株角果數表現出更強的雜種優勢可能與其與母本華航901在不同密度條件下相比父本具有更多的單株角果數有關, 因為從具體數值與動態變化規律上看, 在D1密度下盡管浙油50比華航901有更多的單株角果數, 但隨密度增加而急劇下降, 在D2及更高密度下, 華航901在武漢試點則具有更多的單株角果數, 在西寧試點也與浙油50相當(圖1-A, B)。在西寧試點, 正、反交組合均表現出較強的雜種優勢, 且在低密度下正交組合雜種優勢高于反交組合, 而在高密度下反交組合雜種優勢高于正交組合(圖3-B)。
縮寫和處理同圖1。不同小寫字母表示相同品種密度間差異達顯著水平(< 0.05)。
Abbreviations and treatments are the same as those given in Fig. 1. Values followed by different lowercase letters are significantly different among different densities in the same variety at< 0.05.
在單株產量方面, 正交組合的平均雜種優勢指數顯著高于反交組合(<0.05)。以武漢試點為例, 正交組合的平均雜種優勢指數達到107.3%, 而反交組合為85.3%, 無雜種優勢。另外, 在D2密度下, 正交組合單株產量的雜種優勢指數達到最高(120.8%)。與此同時, 在兩點試驗中, 正交組合在D1、D2密度時表現出較高的雜種優勢, 而反交組合在D4密度時最高。但是, 在武漢試點時反交組合并無明顯的單株產量雜種優勢, 僅在西寧試點時D3、D4密度下表現出一定的雜種優勢(圖3-C, D)。在實際產量方面, 正交組合的平均雜種優勢指數顯著高于反交組合(<0.05)。以西寧試點為例, 正交組合的平均雜種優勢指數達到111.8%, 而反交組合為99.4%, 無雜種優勢。另外, 在武漢試點正交組合的實際產量雜種優勢指數隨密度的增加呈先上升后下降的趨勢, 在D3密度時最高, 而在兩試驗點中反交組合最高雜種優勢指數均處于D4密度下(圖3-E, F)。
在兩點試驗中, 正交組合經濟系數的雜種優勢指數顯著高于反交組合(<0.05)。正交組合各密度下的平均雜種優勢指數分別達到104.7.7% (武漢試點)和105.7% (西寧試點), 而反交組合分別為92.7%和94.4%, 無雜種優勢。與其他產量性狀相比, 兩親本在經濟系數方面展現出較高的穩定性, 但是正交組合具有較強的雜種優勢, 而反交組合經濟系數顯著低于正交組合(<0.05), 也低于中親值, 無雜種優勢。反交組合的經濟系數比正交組合低, 可能由其生物產量大、籽粒產量低所致(圖3-G, H)。
2.2 種植密度對倒伏相關性狀的影響
2.2.1 主要農藝性狀 隨種植密度增加, 株高、根頸粗、地上部鮮重和根鮮重均呈減小趨勢, 在D5密度時達到最小值, 各品種兩點試驗變化趨勢一致(圖4-A, B, E~J); 有效分枝高度隨密度的增加而先增后降, 各品種兩點試驗變化趨勢一致(圖4-C, D)。比較發現, 在同一密度下正交組合(華航901×浙油50)的根頸粗和根鮮重在各品種中最大, 而華航901的株高和地上部生物量低于浙油50。正交組合的株高略低于反交組合(浙油50×華航901), 而其地上部鮮重在D1、D2密度下高于反交組合, 隨密度增加至D3以上時逐漸低于反交組合(圖4-A~J)。方差分析表明, 兩點試驗的品種和密度對油菜成熟期各農藝性狀的影響均達極顯著水平(<0.01), 且它們的互作影響也達極顯著水平(附表2和附表3)。
2.2.2 倒伏相關性狀 莖稈抗折力隨種植密度的增加呈下降趨勢, 在D5密度時達到最小值, 各品種兩點變化規律一致。比較發現, 在同一密度下浙油50平均抗折力大于華航901, 而反交組合(浙油50×華航901)大于正交組合(華航901×浙油50), 只有在武漢試點時正交組合在D1、D2密度下大于反交組合(圖4-K, L)。由圖5-A和B可知, 兩點試驗的平均倒伏指數隨密度的變化規律不同。在武漢試點, 隨密度的增加各品種倒伏指數呈下降趨勢; 在西寧試點, 隨密度增加各品種倒伏指數呈先降后增, 在D5密度時倒伏指數達最大。比較分析發現, 父、母本及正、反交組合在兩點試驗中表現出不同規律。在武漢試點, 倒伏指數表現出華航901>正交組合>浙油50>反交組合的特點, 而在西寧試點則表現出華航901<正交組合<浙油50<反交組合的趨勢。可以看出, 正交組合倒伏指數在武漢與其母本華航901相近, 且遠高于其父本浙油50與反交組合, 抗倒性較差。相反, 在西寧試點, 華航901與正交組合的倒伏指數則遠低于浙油50與反交組合, 表現出較好的抗倒性。綜合兩點試驗, 雜種組合的倒伏指數隨密度的變規律與母本相一致(圖5-A, B)。方差分析表明, 年份(點)、品種和密度對油菜各段倒伏指數的影響均達極顯著水平(<0.01), 且它們的兩兩互作與三者互作影響也達極顯著水平(附表4)。
縮寫和處理同圖1。Abbreviations and treatments are the same as those given in Fig. 1.
莖稈節間最大倒伏指數可以衡量油菜在各密度下的倒伏風險。兩點試驗的最大倒伏指數表現出不同的規律, 但同樣地, 無論正交還是反交組合均與母本相一致(圖5-C, D)。通過分析有效分枝以下4段的平均倒伏指數可知, 從第1段至第4段倒伏指數呈先上升后下降的趨勢, 兩點試驗一致。例如在西寧試點時, 父、母本及雜交組合在D4、D5高密度種植時, 第2~3段比第1、4段的倒伏指數更大。因此, 油菜莖稈中段為易倒伏部位, 存在倒伏風險較高(圖5-E, F)。
2.2.3 正、反交組合倒伏相關性狀的雜種優勢
油菜倒伏數越高越易倒伏, 因此當倒伏指數的雜種優勢指數低于100%時, 說明雜交組合的平均倒伏指數低于中親值, 在抗倒性上具有雜種優勢。隨種植密度增加, 父、母本及雜交組合地上部鮮重與抗折力的雜種優勢指數呈降低趨勢, 各品種兩點試驗變化趨勢一致(圖6-A~D)。而倒伏指數的雜種優勢指數隨密度變化規律不明顯。在武漢試點, 正交組合高于反交組合13.0%; 而在西寧試點, 反交組合高于正交組合36.8%。因此, 在武漢試點, 反交組合相較于正交組合擁有更好的抗倒性; 而在西寧試點, 正交組合則擁有較好的抗倒性(圖6-E, F)。通過分析4段倒伏性狀的雜種優勢指數可知, 在同一密度下從第1段至第4段抗折力雜種優勢指數呈上升趨勢, 兩點試驗一致。另外, 在西寧試點時第1段至第4段倒伏指數雜種優勢指數呈下降趨勢, 而在武漢試點其變化趨勢不明顯(附表5)。
P1、P2、P3和P4分別表示除去縮頸段后將主莖平均分為4段, 從地面往上第1、2、3、4段。圖中不同字母表示各處理差異達顯著水平(< 0.05)。其他縮寫和處理同圖1。
P1, P2, P3, and P4 indicate the first, second, third and fourth part from bottom to the top of the main stem. Values followed by different letters are significantly different among different treatments at< 0.05. Other abbreviations and treatments are the same as those given in Fig. 1.
縮寫和處理同圖1。不同小寫字母表示相同品種密度間差異達顯著水平(< 0.05)。
Abbreviations and treatments are the same as those given in Fig. 1. Values followed by different letters are significantly different among different densities in the same variety at< 0.05.
2.3 雙親及正反雜交后代最佳種植密度的擬合分析
在兩點試驗中, 采用多項式曲線擬合分析各品種實際產量與密度之間的聯系, 獲得了各品種最高產量下的合理密度范圍。通過擬合分析得出, 浙油50與華航901獲得高產的合理密度范圍分別為44.0~52.3萬株hm-2與47.1~49.7萬株hm-2, 而正交組合(華航901×浙油50)與反交組合(浙油50×華航901)的高產密度范圍則分別為42.8~49.4萬株hm-2與49.5~53.7萬株hm-2, 預測在西寧試點正、反交組合在最適密度下獲得的最高產量, 分別為4381.7 kg hm-2和4208.1 kg hm-2。在合理種植密度范圍近似的情況下, 雜交油菜具有更高的產量, 所以雜交組合后代比雙親更耐密植(圖7-A, B)。在倒伏相關性狀方面, 兩點間變化規律不同, 在武漢試點時倒伏指數隨密度增加而顯著降低(<0.05), 而在西寧試點則呈現出先增后降的趨勢。擬合分析發現, 由于各品種兩點的倒伏指數差異較大, 所以以西寧試點(正交組合與母本華航901倒伏指數較低)為標準。分析結果表明, 正交組合與反交組合最小倒伏指數的合理密度分別為49.9萬株hm-2與32.3萬株hm-2, 倒伏指數分別為1.86 cm g g-1和2.97 cm g g-1, 而浙油50與華航901分別為41.2萬株hm-2與47.0萬株hm-2, 倒伏指數分別為2.38 cm g g-1和1.69 cm g g-1(圖7-C, D)。說明, 浙油50、華航901和正交組合(華航901×浙油50)可以通過優化密度良好地協調產量與倒伏的矛盾, 合理密度范圍分別為41.2~52.3萬株hm-2、47.0~49.7萬株hm-2和42.8~49.9萬株hm-2。然而, 反交組合在抗倒伏和產量的合理密度范圍差異極大, 并不能很好地通過優化種植密度來緩解高產與抗倒之間的矛盾。
3 討論
3.1 合理密植協調油菜高產與抗倒性
種植密度是一種重要的栽培措施, 影響著油菜全生育時期的群體結構與光能利用效率, 通過改變油菜的源庫流平衡最終影響籽粒產量[37]。種植密度影響油菜各農藝性狀的表現, 包括株高、分枝高度和分枝角度等株型性狀, 單株角果數、每角果粒數和籽粒重等產量構成因子, 還包括莖粗、莖稈抗折力等倒伏相關性狀。在單株產量隨密度增加而降低的情況下, 群體產量的變化規律顯得尤為重要, 也可作為衡量一個油菜品種是否適宜密植的關鍵指標。優化油菜種植密度可以適當地調節個體生長與群體競爭之間的矛盾, 從而優化高產與抗倒之間的矛盾[15]。然而, 前人研究多集中于常規品種或生產上推廣的雜交品種, 關于種植密度協調正、反交組合的高產與抗倒的研究鮮有報道。
縮寫同圖1。**表示1%顯著水平。
Abbreviations are the same as those given in Fig. 1.**Significant at the 1% probability level.
在本研究中, 隨種植密度增加, 單株角果數、每角果粒數與單株產量逐漸降低, 而千粒重變化規律不明顯。雖然單株產量隨密度增加而顯著降低, 但是實際產量隨密度增加則表現出先增加后降低的趨勢。綜合分析實產和密度之間的關系發現, 各品種獲得高產的合理密度范圍多集中在40~50萬株hm-2之間, 而雜交油菜較親本具有更高的產量。因此, 我們推測本研究中正、反交組合比雙親具有更好的耐密性, 在合理密植條件下正交組合(華航901×浙油50)優于反交組合(浙油50×華航901)的產量表現。在抗倒性方面, 隨密度的增加在西寧試點各品種的倒伏指數表現出先下降后上升的趨勢, 而在武漢試點呈降低趨勢, 在高密度下降幅變緩。倒伏指數受環境因素影響較大, 所以兩點試驗具體數值和變化規律差異較大。正交組合(華航901×浙油50)的最小倒伏指數的合理密度在其高產密度范圍附近, 可通過合理密植獲得高產與抗倒的平衡。然而, 反交組合(浙油50×華航901)在抗倒和高產的合理密度范圍差異較大, 并不能通過優化種植密度來緩解高產與抗倒之間的矛盾。
3.2 正、反交油菜的雜種優勢表現
在油菜雜交選育過程中, 性狀多數受基因組中核基因的控制, 目標性狀遺傳差異較大的親本配對通常會獲得雜種優勢。前人研究發現, 無論正反交, 雜交油菜籽粒的千粒重和大小均表現出顯著的超親優勢, 在雜交育種中可通過合理養分分配來提高粒重達到增產效果[38]。在本研究中, 我們以雜種優勢指數作為衡量雜種優勢的重要指標, 考察了包括產量和倒伏相關等多個油菜重要生產性狀, 全面對比了浙油50與華航901兩親本和正、反交組合后代的雜種優勢表現。在產量方面, 正交組合(華航901×浙油50)的單株產量與實際產量均表現出優于中親值的雜種優勢, 而反交組合(浙油50×華航901)平均雜種優勢指數低于100%, 無雜種優勢。另外, 在兩點試驗中, 正交組合的經濟系數雜種優勢顯著高于反交組合, 且反交組合無雜種優勢, 且各密度水平下規律一致。由此可知, 正交組合相較于反交組合表現出更為優秀的產量雜種優勢。在抗倒伏方面, 可能由于生長環境的差異, 試驗各品種油菜倒伏指數在年際間產生較大差異, 從而正、反交組合倒伏指數雜種優勢大小也在兩點間各不相同。在武漢試點, 正、反交組合皆表現出雜種優勢, 且反交組合更高; 而在西寧試點, 正交組合擁有較高的雜種優勢, 而反交組合并不保持雜種優勢。
3.3 雜交油菜的遺傳效應與親本選配
前人研究表明, 粒重受油菜母本基因型的控制, 可能的原因是在角果發育時期, 角果作為源在光合作用和干物質轉運方面發揮著重要作用, 向作為庫的種子運輸有機質[25]。近年來, 研究人員也發現含油量具有母體遺傳特征, 這與角果皮的光合作用及細胞質效應緊密相關[34,36]。本研究中, 在產量構成因子方面, 父、母本及正、反交組合的單株角果數與每角果粒數均隨密度增加而顯著降低, 且無論正反交, 雜種后代均與母本更為接近, 反映出一定的母體效應; 而千粒重的變化缺乏規律; 相應地, 單株產量隨密度增加而降低。然而, 在兩點試驗中實際產量則隨密度增加呈現出先增后降的趨勢, 且不論正反交, 其變化規律均與父本相一致, 這顯然不符合母體遺傳的特征, 反映出實際產量隨種植密度變化的復雜性, 具體原因尚需要進一步研究。王瑞等[39]的研究也表明, 甘藍型黃籽油菜產量性狀的遺傳主要由加性效應和顯性效應共同控制, 母體效應影響較小。但是在抗倒性方面, 盡管不同試驗點間規律存在差異, 但是不論正反交, 其倒伏指數動態變化規律均與母本相一致(圖5-A~D)。因此, 我們認為在油菜生產中可以選擇含油量高、抗倒品種作為母本, 高產品種作父本選配雜交組合, 以選育并獲得高產、抗倒且耐密的優質雜交油菜品種。當然, 我們也注意到, 由于研究材料或者考察指標的不同, 倒伏性狀的遺傳規律可能不同。例如, 以單株抗壓力反映的倒伏性狀的遺傳分析表明, 不同組合正反交F1差異不顯著, 未見明顯的細胞質效應[40]。
4 結論
在本試驗中, 浙油50、華航901和其正、反交組合的單株角果數、每角果粒數和單株產量隨種植密度增加呈降低趨勢, 而實際產量呈先增后降的趨勢。正交組合(華航901×浙油50)的實際產量和經濟系數顯著高于反交組合(浙油50×華航901)。擬合分析發現, 正交組合的合理密度范圍為42.8~49.9萬株hm-2, 在此密度下可以良好地協調產量與倒伏之間的矛盾。在相同密度下, 正交組合比反交組合有更大的實產雜種優勢, 而在倒伏性狀方面正交組合與其母本華航901隨密度變化規律相似。因此, 為了實現油菜高效生產, 可選擇高產品種為父本, 含油量高、抗倒性強的品種為母本進行雜交組合, 以此作為雜交油菜的親本選配原則。需要指出的是, 由于類似研究在油菜中還鮮有報道, 且本研究相關結果也僅基于兩個油菜品種間的正反交獲得, 不能排除不同親本的差異對正交和反交效應的影響。因此, 相關結論推廣到其他品種時還需要設計更多的正反雜交試驗加以驗證。
[1] 劉成, 黃杰, 冷博峰, 馮中朝, 李俊鵬. 我國油菜產業現狀, 發展困境及建議. 中國農業大學學報, 2017, 22(12): 203–210.
Liu C, Huang J, Leng B F, Feng Z C, Li J P. Current situation, development difficulties and suggestions of Chinese rape industry., 2017, 22(12): 203–210 (in Chinese with English abstract).
[2] Food and Agriculture Organization (FAO) of the United Nations. FAO Statistical Databases in 2018. [2020-09-20]. http://www. fao.org.
[3] 劉成, 馮中朝, 肖唐華, 馬曉敏, 周廣生, 黃鳳洪, 李加納, 王漢中. 我國油菜產業發展現狀、潛力及對策. 中國油料作物學報, 2019, 41: 485–489.
Liu C, Feng Z C, Xiao T H, Ma X M, Zhou G S, Huang F H, Li J N, Wang H Z. Development, potential and adaptation of Chinese rapeseed industry., 2019, 41: 485–489 (in Chinese with English abstract).
[4] 熊秋芳, 文靜, 李興華, 沈金雄. 中國油菜科技創新與產業發展. 中國農業科技導報, 2014, 16(3): 14–22.
Xiong Q F, Wen J, Li X H, Shen J X. Technological innovation and industrial development of rapeseed in China., 2014, 16(3): 14–22 (in Chinese with English abstract).
[5] 李小勇, 周敏, 王濤, 張蘭, 周廣生, 蒯婕. 種植密度對油菜機械收獲關鍵性狀的影響. 作物學報, 2018, 44: 278–287.
Li X Y, Zhou M, Wang T, Zhang L, Zhou G S, Kuai J. Effects of planting density on the mechanical harvesting characteristics of semi-winter rapeseed., 2018, 44: 278–287 (in Chinese with English abstract).
[6] 安偉, 樊智翔, 郭玉宏, 米小紅, 徐瀾. 玉米品種的增產潛力與改良方向. 山西農業大學學報, 2003, 23: 386–388.
An W, Fan Z X, Guo Y H, Mi X H, Xu L. Utilization and quality improvement of high starch maize., 2003, 23: 386–388 (in Chinese with English abstract).
[7] 李豪圣, 宋健民, 劉愛峰, 程敦公, 王西芝, 杜長林, 趙振東, 劉建軍. 播期和種植密度對超高產小麥‘濟麥22’產量及其構成因素的影響. 中國農學通報, 2011, 27(5): 243–248.
Li H S, Song J M, Liu A F, Cheng D G, Wang X Z, Du C L, Zhao Z D, Liu J J. Effect of sowing time and planting density on yield and components of ‘Jimai22’ with super-high yield., 2011, 27(5): 243–248 (in Chinese with English abstract).
[8] 劉浩, 王加峰, 孫大元, 郭濤, 劉永柱, 王慧, 陳志強. 水稻矮稈基因研究進展. 廣東農業科學, 2013, 40(22): 139–144.
Liu H, Wang J F, Sun D Y, Guo T, Liu Y Z, Wang H, Chen Z Q. Research progress of dwarf genes in rice., 2013, 40(22): 139–144 (in Chinese with English abstract).
[9] 王成璦, 王伯倫, 張文香, 趙磊, 趙秀哲, 高連文. 栽培密度對水稻產量及品質的影響. 沈陽農業大學學報, 2004, 35: 318–322.
Wang C A, Wang B L, Zhang W X, Zhao L, Zhao X Z, Gao L W. Effect of planting density on grain yield and quality of rice., 2004, 35: 318–322 (in Chinese with English abstract).
[10] 楊國虎, 李新, 王承蓮, 羅湘寧. 種植密度影響玉米產量及部分產量相關性狀的研究. 西北農業學報, 2006, 15(5): 57–60.
Yang G H, Li X, Wang C L, Luo X N. Study on effects of plant densities on the yield and the related characters of maize hybrids., 2006, 15(5): 57–60 (in Chinese with English abstract).
[11] 周江明, 趙琳, 董越勇, 徐進, 邊武英, 毛楊倉, 章秀福. 氮肥和栽植密度對水稻產量及氮肥利用率的影響. 植物營養與肥料學報, 2010, 16: 274–281.
Zhou J M, Zhao L, Dong Y Y, Xu J, Bian W Y, Mao Y C, Zhang X F. Nitrogen and transplanting density interactions on the rice yield and N use rate., 2010, 16: 274–281 (in Chinese with English abstract).
[12] Li H T, Li J J, Song J R, Zhao B, Guo C C, Wang B, Zhang Q H, Wang J, King G J, Liu K D. An auxin signaling genecontributes to improved plant architecture and yield heterosis in rapeseed., 2019, 222: 837–851.
[13] Ma N, Yuan J Z, Li M, Li J, Zhang L Y, Liu L X, Naeem M S, Zhang C L. Ideotype population exploration: growth, photosynthesis, and yield components at different planting densities in winter oilseed rape (L.)., 2014, 9: e114232.
[14] Kuai J, Sun Y Y, Zhou M, Zhang P P, Zuo Q S, Wu J S, Zhou G S. The effect of nitrogen application and planting density on the radiation use efficiency and the stem lignin metabolism in rapeseed (L.)., 2016, 199: 89–98.
[15] 蒯婕, 王積軍, 左青松, 陳紅琳, 高建芹, 汪波, 周廣生, 傅廷棟. 長江流域直播油菜密植效應及其機理研究進展. 中國農業科學, 2018, 51: 4625–4632.
Kuai J, Wang J J, Zuo Q S, Chen H L, Gao J Q, Wang B, Zhou G S, Fu T D. Effects and mechanism of higher plant density on directly-sown rapeseed in the Yangtze River Basin of China., 2018, 51: 4625–4632 (in Chinese with English abstract).
[16] 趙永國, 程勇, 陸光遠, 徐勁松, 付桂萍, 鄒錫玲, 喬醒, 劉清云, 張學昆. 密植條件下國審冬油菜品種主要性狀特點與差異. 中國油料作物學報, 2015, 37: 285–290.
Zhao Y G, Cheng Y, Lu G Y, Xu J S, Fu G P, Zou X L, Qiao X, Liu Q Y, Zhang X K. Characteristics and variation of winter rapeseed (L.) cultivars under high density., 2015, 37: 285–290 (in Chinese with English abstract).
[17] 宋稀, 劉鳳蘭, 鄭普英, 張學昆, 陸光遠, 付桂萍, 程勇. 高密度種植專用油菜重要農藝性狀與產量的關系分析. 中國農業科學, 2010, 43: 1800–1806.
Song X, Liu F L, Zheng P Y, Zhang X K, Lu G Y, Fu G P, Cheng Y. Correlation analysis between agronomic traits and yield of rapeseed (L.) for high-density planting., 2010, 43: 1800–1806 (in Chinese with English abstract).
[18] 王寅, 魯劍巍. 中國冬油菜栽培方式變遷與相應的養分管理策略. 中國農業科學, 2015, 48: 2952–2966.
Wang Y, Lu J W. The transitional cultivation patterns of winter oilseed rape in China and the corresponding nutrient management strategies., 2015, 48: 2952–2966 (in Chinese with English abstract).
[19] Diepenbrock W. Yield analysis of winter oilseed rape (L.): a review., 2000, 67: 35–49.
[20] Hu Q, Hua W, Yin Y, Zhang X, Liu L, Shi J, Zhao Y, Qin L, Chen C, Wang H Z. Rapeseed research and production in China., 2017, 5: 127–135.
[21] Huang X H, Yang S H, Gong J Y, Zhao Q, Feng Q, Zhan Q L, Zhao Y, Li W J, Cheng B Y, Xia J H, Chen N, Huang T, Zhang L, Fan D L, Chen J Y, Zhou C C, Lu Y Q, Weng Q J, Han B. Genomic architecture of heterosis for yield traits in rice., 2016, 537: 629–633.
[22] 傅廷棟. 中國油菜生產和品種改良的現狀與前景. 安徽農學通報, 2000, 6(1): 3–10.
Fu T D. Current status and prospects of rape production and variety improvement in China., 2000, 6(1): 3–10 (in Chinese with English abstract).
[23] 烏蘭, 馬偉杰, 義如格勒圖, 崔仲楠, 魯興華. 油菜秸稈飼用價值分析及其開發利用. 內蒙古草業, 2007, 19(1): 41–42.
Wu L, Ma W J, Yi R G L T, Cui Z N, Lu X H. Feeding value analysis and development and utilization of rapeseed straw., 2007, 19(1): 41–42 (in Chinese with English abstract).
[24] 左旭. 我國農業廢棄物新型能源化開發利用研究. 中國農業科學院博士學位論文, 北京, 2015.
Zuo X. A Research on the Development and Utilization of the Agricultural Residues as New Sources Energy in China. PhD Dissertation of Chinese Academy of Agricultural Sciences, Beijing, China, 2015 (in Chinese with English abstract).
[25] Li N, Song O J, Peng W, Zhan J P, Shi J Q, Wang X F, Liu G H, Wang H Z. Maternal control of seed weight in rapeseed (L.): the causal link between the size of pod (mother, source) and seed (offspring, sink).2019, 17: 736–749.
[26] Guo M, Rupe M A, Dieter J A, Zou J J, Spielbauer D, Duncan K E, Howard R J, Hou Z L, Simmons C R. Cell number regulator1 affects plant and organ size in maize: implications for crop yield enhancement and heterosis., 2010, 22: 1057–1073.
[27] Ma J, Zhang D F, Cao Y Y, Wang L F, Li J J, Lubberstedt T, Wang T Y, Li Y, Li H Y. Heterosis-related genes under different planting densities in maize., 2018, 69: 5077–5087.
[28] Qian C R, Yu Y, Gong X J, Jiang Y B, Zhao Y, Yang Z L, Hao Y B, Li L, Song Z W, Zhang W J. Response of grain yield to plant density and nitrogen rate in spring maize hybrids released from 1970 to 2010 in Northeast China., 2016, 4: 459–467.
[29] 沈金雄, 傅廷棟, 楊光圣, 馬朝芝, 涂金星. 甘藍型油菜雜種優勢及產量性狀的遺傳改良. 中國油料作物學報, 2005, 27(1): 5–9.
Shen J X, Fu T D, Yang G S, Ma C Z, Tu J X. Analysis of heterosis reveals genetic improvement for yield traits in rapeseed (L.)., 2005, 27(1): 5–9 (in Chinese with English abstract).
[30] 劉絢霞, 董振生, 劉創社, 董軍剛, 李紅兵. 甘藍型優質雜交油菜主要農藝性狀配合力與遺傳力研究. 中國油料作物學報, 2001, 23(3): 2–5.
Liu X X, Dong Z S, Liu C S, Dong J G, Li H B. Combining ability and genetic ability of main agronomy characters in double-low rapeseed (L.)., 2001, 23(3): 2–5 (in Chinese with English abstract).
[31] 王天宇, 祝云芳, 陳華璋, 陳澤輝. 玉米正反交雜交種F1主要性狀的差異性分析. 玉米科學, 2007, 15(4): 52–55.
Wang T Y, Zhu Y F, Chen H Z, Chen Z H. Study on Differences of main characters between maize hybrids and reciprocal crosses., 2007, 15(4): 52–55 (in Chinese with English abstract).
[32] 張耀文, 趙小光, 田建華, 王輝, 王學芳, 李殿榮, 侯君利, 關周博, 韋世豪. 甘藍型油菜正反交組合葉片凈光合速率和葉綠素含量的比較. 華北農學報, 2015, 30(5): 135–140.
Zhang Y W, Zhao X G, Tian J H, Wang H, Wang X F, Li D R, Hou J L, Guan Z B, Wei S H.Comparison of net photosynthetic rate and chlorophyll content ofbetween orthogonal and reciprocal combinations., 2015, 30(5): 135–140 (in Chinese with English abstract).
[33] Magrath R, Mithen R. Maternal effects on the expression of individual aliphatic glucosinolates in seeds and seedlings of., 1993, 111: 249–252.
[34] Hua W, Li R J, Zhan G M, Liu J, Li J, Wang X F, Liu G H, Wang H Z. Maternal control of seed oil content in: the role of silique wall photosynthesis., 2012, 69: 432–444.
[35] Li N, Peng W, Shi J Q, Wang X F, Liu G H, Wang H Z. The natural variation of seed weight is mainly controlled by maternal genotype in rapeseed (L.)., 2015, 10: e0125360.
[36] Liu J, Hao W J, Liu J, Fan S H, Zhao W, Deng L B, Wang X F, Hu Z Y, Hua W, Wang H Z. A novel chimeric mitochondrial gene confers cytoplasmic effects on seed oil content in polyploid rapeseed ()., 2019, 12: 582–596.
[37] 王學芳, 田建華, 董育紅, 關周博, 楊麗. 不同密度緊湊型油菜的源庫特征及與收獲指數的相關研究. 中國農學通報, 2020, 36(18): 33–38.
Wang X F, Tian J H, Dong Y H, Guan Z B, Yang L. Compact rapeseed under different densities: sink-source characteristics and their correlation with harvest index., 2020, 36(18): 33–38 (in Chinese with English abstract).
[38] 孫超, 汪騫, 唐偉杰, 李麗萍, 夏張婷, 趙海洋, 韓月鑫, 林良斌. 受母性影響的油菜種子性狀研究. 種子, 2018, 37(5): 9–12.
Sun C, Wang Q, Tang W J, Li L P, Xia Z T, Zhao H Y, Han Y X, Lin L B. Study on the characters of seed inaffected by maternale effects., 2018, 37(5): 9–12 (in Chinese with English abstract).
[39] 王瑞, 李加納, 唐章林, 諶利, 張學昆. 甘藍型黃籽油菜產量性狀的遺傳分析. 中國農學通報. 2004, 20(5): 37?38.
Wang R, Li J N, Tang Z L, Chen L, Zhang X K. Genetic analysis for yield characters in yellow-seeded rapeseed lines (L.).. 2004, 20(5): 37–38 (in Chinese with English abstract).
[40] 顧慧, 戚存扣. 甘藍型油菜(L.)抗倒性狀的主基因+多基因遺傳分析. 作物學報, 2008, 34: 376?381.
Gu H, Qi C K. Genetic analysis of lodging resistance with mixed model of major gene plus polygene inL., 2008, 34: 376?381 (in Chinese with English abstract).
附表1 種植密度對油菜產量與產量構成因子的影響
Table S1 Effects of planting densities on yield and yield components in rapeseed
(續附表1)
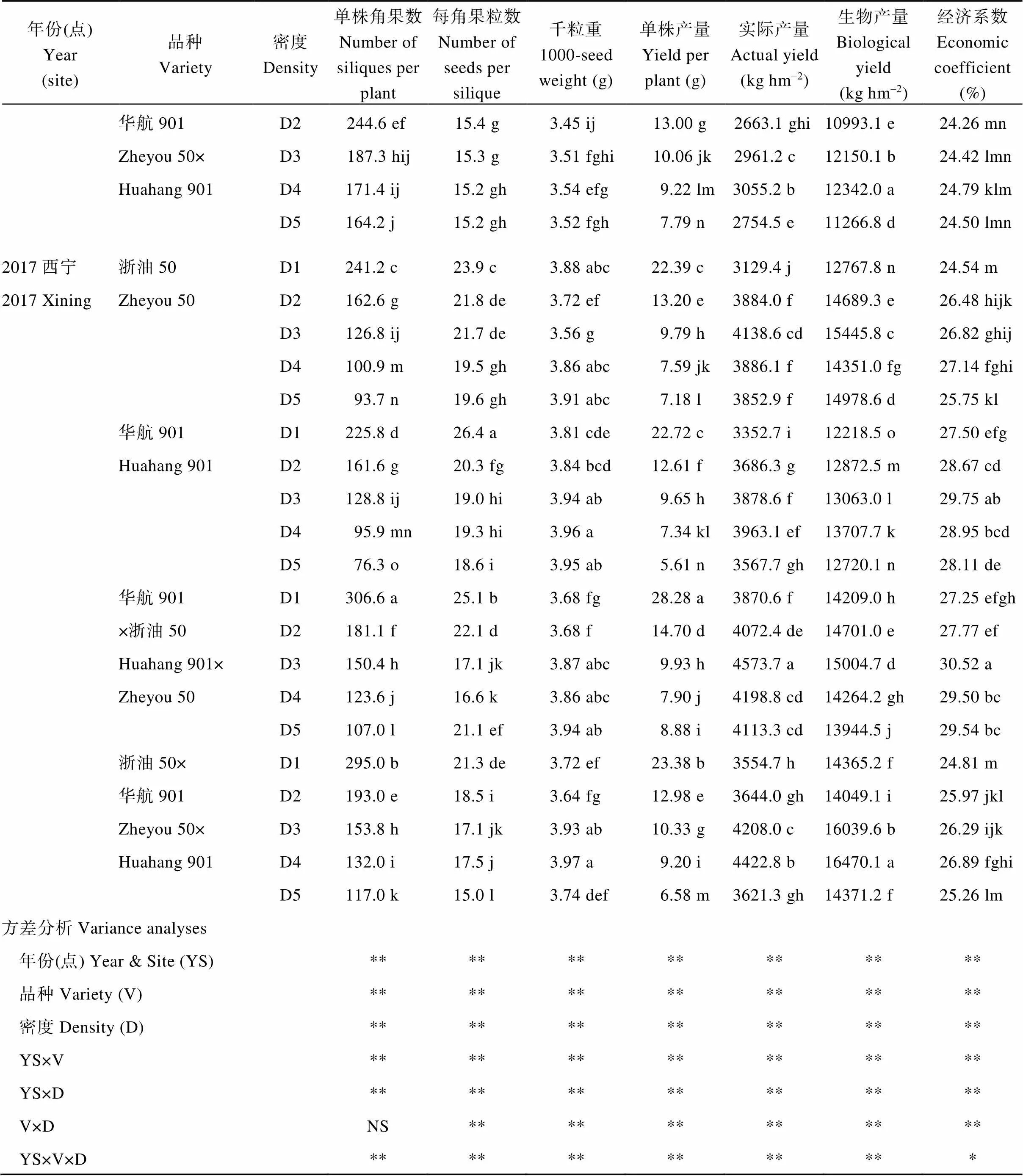
年份(點)Year(site)品種Variety密度Density單株角果數Number of siliques per plant每角果粒數Number of seeds per silique千粒重1000-seed weight (g)單株產量Yield per plant (g)實際產量Actual yield(kg hm–2)生物產量Biological yield(kg hm–2)經濟系數Economic coefficient(%) 華航901D2244.6 ef15.4 g3.45 ij13.00 g2663.1 ghi10993.1 e24.26 mn Zheyou 50×D3187.3 hij15.3 g3.51 fghi10.06 jk2961.2 c12150.1 b24.42 lmn Huahang 901D4171.4 ij15.2 gh3.54 efg9.22 lm3055.2 b12342.0 a24.79 klm D5164.2 j15.2 gh3.52 fgh7.79 n2754.5 e11266.8 d24.50 lmn 2017西寧浙油50D1241.2 c23.9 c3.88 abc22.39 c3129.4 j12767.8 n24.54 m 2017 XiningZheyou 50D2162.6 g21.8 de3.72 ef13.20 e3884.0 f14689.3 e26.48 hijk D3126.8 ij21.7 de3.56 g9.79 h4138.6 cd15445.8 c26.82 ghij D4100.9 m19.5 gh3.86 abc7.59 jk3886.1 f14351.0 fg27.14 fghi D593.7 n19.6 gh3.91 abc7.18 l3852.9 f14978.6 d25.75 kl 華航901D1225.8 d26.4 a3.81 cde22.72 c3352.7 i12218.5 o27.50 efg Huahang 901D2161.6 g20.3 fg3.84 bcd12.61 f3686.3 g12872.5 m28.67 cd D3128.8 ij19.0 hi3.94 ab9.65 h3878.6 f13063.0 l29.75 ab D495.9 mn19.3 hi3.96 a7.34 kl3963.1 ef13707.7 k28.95 bcd D576.3 o18.6 i3.95 ab5.61 n3567.7 gh12720.1 n28.11 de 華航901D1306.6 a25.1 b3.68 fg28.28 a3870.6 f14209.0 h27.25 efgh ×浙油50D2181.1 f22.1 d3.68 f14.70 d4072.4 de14701.0 e27.77 ef Huahang 901×D3150.4 h17.1 jk3.87 abc9.93 h4573.7 a15004.7 d30.52 a Zheyou 50D4123.6 j16.6 k3.86 abc7.90 j4198.8 cd14264.2 gh29.50 bc D5107.0 l21.1 ef3.94 ab8.88 i4113.3 cd13944.5 j29.54 bc 浙油50×D1295.0 b21.3 de3.72 ef23.38 b3554.7 h14365.2 f24.81 m 華航901D2193.0 e18.5 i3.64 fg12.98 e3644.0 gh14049.1 i25.97 jkl Zheyou 50×D3153.8 h17.1 jk3.93 ab10.33 g4208.0 c16039.6 b26.29 ijk Huahang 901D4132.0 i17.5 j3.97 a9.20 i4422.8 b16470.1 a26.89 fghi D5117.0 k15.0 l3.74 def6.58 m3621.3 gh14371.2 f25.26 lm 方差分析Variance analyses 年份(點) Year & Site (YS)************** 品種Variety (V)************** 密度Density (D)************** YS×V************** YS×D************** V×DNS************ YS×V×D*************
D1、D2、D3、D4和D5分別表示種植密度為1.5×105、3.0×105、4.5×105、6.0×105和7.5×105株hm–2。不同小寫字母表示同一年份同一品種不同處理間差異達顯著水平(< 0.05);*、**分別表示在0.05和0.01水平顯著差異,NS表示差異不顯著。
D1, D2, D3, D4, and D5 indicate the planting densities of 1.5×105, 3.0×105, 4.5×105, 6.0×105, and 7.5×105plants hm–2, respectively. Values followed by different lowercase letters are significantly different in the same year and the same variety among different treatments at the 0.05 probability level; * and ** indicate significant difference at the 0.05 and 0.01 probability levels; NS: not significant difference.
附表2 種植密度對油菜成熟期主要農藝性狀的影響(2017年, 武漢)
Table S2 Effects of planting densities on main agronomic traits of rapeseed at maturity (2017, Wuhan)
D1、D2、D3、D4和D5分別表示種植密度為1.5×105、3.0×105、4.5×105、6.0×105和7.5×105株hm–2。同列不同小寫字母表示同一品種不同處理間差異達顯著水平(<0.05);*、**分別表示在0.05和0.01水平顯著差異,NS表示差異不顯著。
D1, D2, D3, D4, and D5 indicate the planting densities of 1.5×105, 3.0×105, 4.5×105, 6.0×105, and 7.5×105plants hm–2, respectively. Values within a column followed by different lowercase letters are significantly different in the same variety among different treatments at the 0.05 probability level; * and ** indicate significant difference at the 0.05 and 0.01 probability levels; NS: not significant difference.
附表3 種植密度對油菜成熟期主要農藝性狀的影響(2017年, 西寧)
Table S3 Effects of planting densities on main agronomic traits of rapeseed at maturity (2017, Xining)
(續附表3)
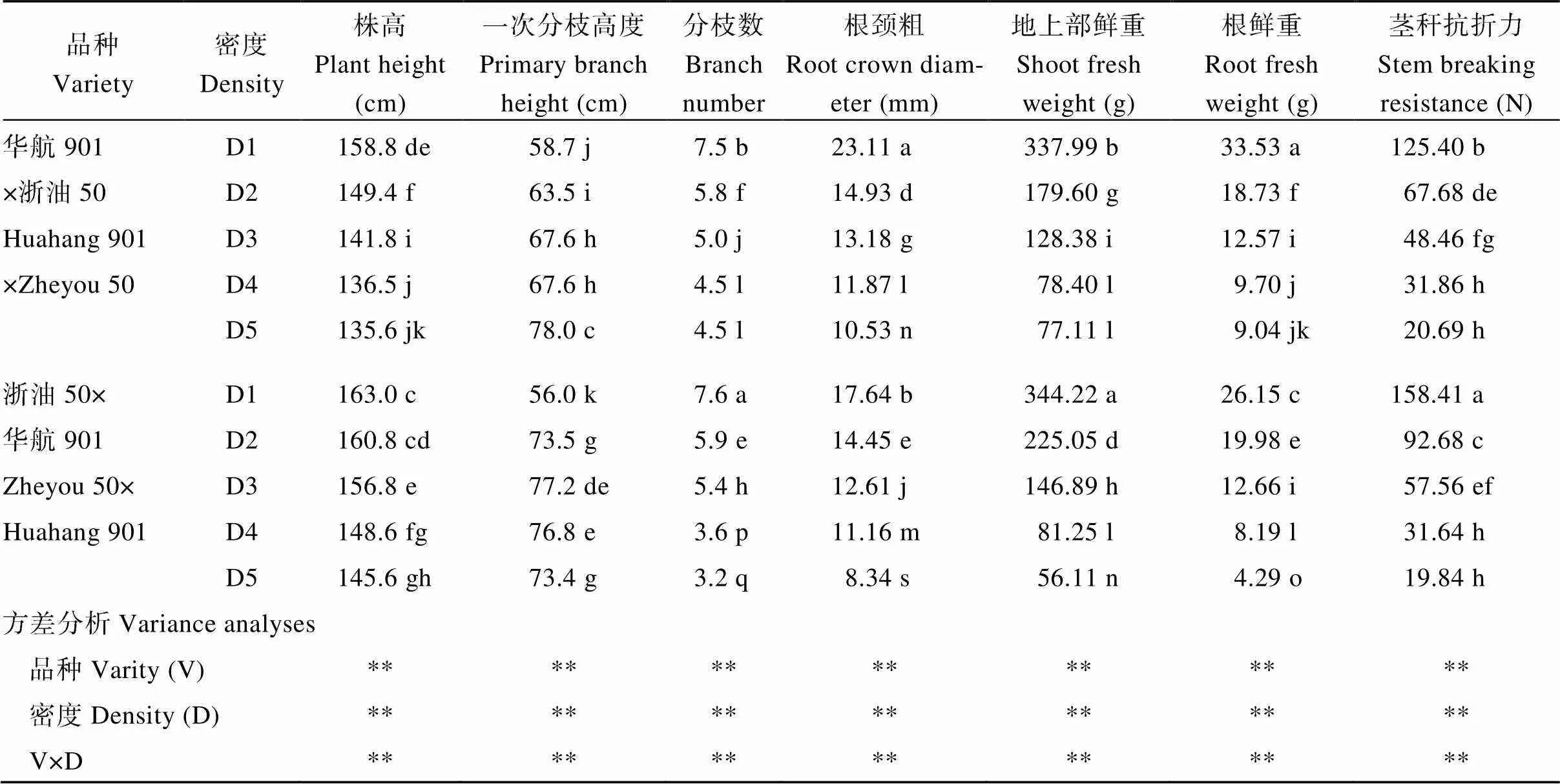
品種Variety密度Density株高Plant height (cm)一次分枝高度Primary branch height (cm)分枝數Branchnumber根頸粗Root crown diameter (mm)地上部鮮重Shoot fresh weight (g)根鮮重Root fresh weight (g)莖稈抗折力Stem breaking resistance (N) 華航901D1158.8 de58.7 j7.5 b23.11 a337.99 b33.53 a125.40 b ×浙油50D2149.4 f63.5 i5.8 f14.93 d179.60 g18.73 f67.68 de Huahang 901D3141.8 i67.6 h5.0 j13.18 g128.38 i12.57 i48.46 fg ×Zheyou 50D4136.5 j67.6 h4.5 l11.87 l78.40 l9.70 j31.86 h D5135.6 jk78.0 c4.5 l10.53 n77.11 l9.04 jk20.69 h 浙油50×D1163.0 c56.0 k7.6 a17.64 b344.22 a26.15 c158.41 a 華航901D2160.8 cd73.5 g5.9 e14.45 e225.05 d19.98 e92.68 c Zheyou 50×D3156.8 e77.2 de5.4 h12.61 j146.89 h12.66 i57.56 ef Huahang 901D4148.6 fg76.8 e3.6 p11.16 m81.25 l8.19 l31.64 h D5145.6 gh73.4 g3.2 q8.34 s56.11 n4.29 o19.84 h 方差分析Variance analyses 品種Varity (V)************** 密度Density (D)************** V×D**************
D1、D2、D3、D4和D5分別表示種植密度為1.5×105、3.0×105、4.5×105、6.0×105和7.5×105株hm–2。同列不同小寫字母表示同一品種不同處理間差異達顯著水平(< 0.05);*、**分別表示在0.05和0.01水平顯著差異,NS表示差異不顯著。
D1, D2, D3, D4 and D5 indicate the planting densities of 1.5×105, 3.0×105, 4.5×105, 6.0×105,and 7.5×105plants hm–2, respectively. Values within a column followed by different lowercase letters are significantly different in the same variety among different treatments at the 0.05 probability level; * and ** indicate significant difference at the 0.05 and 0.01 probability levels; NS: not significant difference.
附表4 種植密度對油菜莖稈不同部位倒伏指數的影響
Table S4 Effects of planting densities on lodging index at different parts of rapeseed stem
(續附表4)
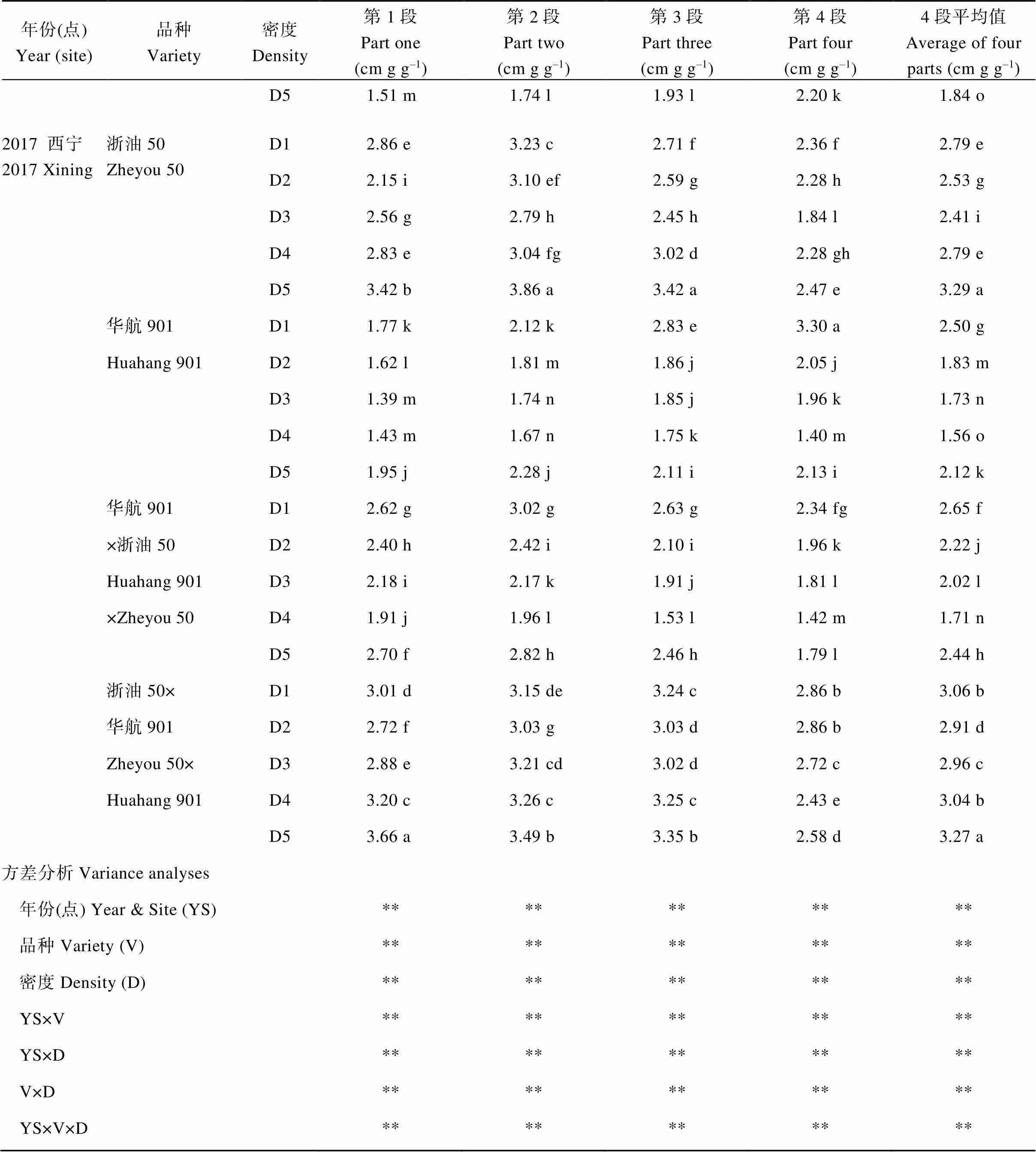
年份(點)Year (site)品種Variety密度Density第1段Part one(cm g g–1)第2段Part two(cm g g–1)第3段Part three(cm g g–1)第4段Part four(cm g g–1)4段平均值Average of four parts (cm g g–1) D51.51 m1.74 l1.93 l2.20 k1.84 o 2017 西寧2017 Xining浙油50Zheyou 50D12.86 e3.23 c2.71 f2.36 f2.79 e D22.15 i3.10 ef2.59 g2.28 h2.53 g D32.56 g2.79 h2.45 h1.84 l2.41 i D42.83 e3.04 fg3.02 d2.28 gh2.79 e D53.42 b3.86 a3.42 a2.47 e3.29 a 華航901D11.77 k2.12 k2.83 e3.30 a2.50 g Huahang 901D21.62 l1.81 m1.86 j2.05 j1.83 m D31.39 m1.74 n1.85 j1.96 k1.73 n D41.43 m1.67 n1.75 k1.40 m1.56 o D51.95 j2.28 j2.11 i2.13 i2.12 k 華航901D12.62 g3.02 g2.63 g2.34 fg2.65 f ×浙油50D22.40 h2.42 i2.10 i1.96 k2.22 j Huahang 901D32.18 i2.17 k1.91 j1.81 l2.02 l ×Zheyou 50D41.91 j1.96 l1.53 l1.42 m1.71 n D52.70 f2.82 h2.46 h1.79 l2.44 h 浙油50×D13.01 d3.15 de3.24 c2.86 b3.06 b 華航901D22.72 f3.03 g3.03 d2.86 b2.91 d Zheyou 50×D32.88 e3.21 cd3.02 d2.72 c2.96 c Huahang 901D43.20 c3.26 c3.25 c2.43 e3.04 b D53.66 a3.49 b3.35 b2.58 d3.27 a 方差分析Variance analyses 年份(點) Year & Site (YS)********** 品種Variety (V)********** 密度Density (D)********** YS×V********** YS×D********** V×D********** YS×V×D**********
D1、D2、D3、D4和D5分別表示種植密度為1.5×105、3.0×105、4.5×105、6.0×105和7.5×105株hm–2;第1段、第2段、第3段和第4段分別表示除去縮頸段后將主莖平均分為4段,從地面往上1、2、3、4段。不同小寫字母表示同一年份同一品種不同處理間差異達顯著水平(< 0.05);*、**分別表示在0.05和0.01水平顯著差異,NS表示差異不顯著。
D1, D2, D3, D4 and D5 indicate the planting densities of 1.5×105, 3.0×105, 4.5×105, 6.0×105,and 7.5×105plants hm–2, respectively; Part One, Part Two, Part Three, and Part Four indicate the first, second, third and fourth part from bottom to the top when averaged the main stem. Values followed by different lowercase letters are significantly different in the same year and the same variety among different treatments at the 0.05 probability level; * and ** indicate significant difference at the 0.05 and 0.01 probability levels; NS: not significant difference.
附表5 種植密度對正反交組合倒伏雜種優勢指數的影響
Table S5 Effects of planting densities on lodging heterosis index of reciprocal combinations
D1、D2、D3、D4和D5分別表示種植密度為1.5×105、3.0×105、4.5×105、6.0×105和7.5×105株hm–2;第1段、第2段、第3段和第4段分別表示除去縮頸段后將主莖平均分為4段,從地面往上1、2、3、4段。不同小寫字母表示同一年份同一品種不同處理間差異達顯著水平(< 0.05)。
D1, D2, D3, D4, and D5 indicate the planting densities of 1.5×105, 3.0×105, 4.5×105, 6.0×105,and 7.5×105plants hm–2, respectively; Part one, Part two, Part three and Part four indicate the first, second, third, and fourth part from bottom to the top when averaged the main stem. Values followed by different lowercase letters are significantly different in the same year and the same variety among different treatments at the 0.05 probability level.
Effects of planting density on yield and lodging related characters of reciprocal hybrids inL.
LOU Hong-Xiang1, JI Jian-Li1, KUAI Jie1, WANG Bo1, XU Liang2, LI Zhen1, LIU Fang3, HUANG Wei4, LIU Shu-Yan5, YIN Yu-Feng6, WANG Jing1,*, and ZHOU Guang-Sheng1
1College of Plant Science and Technology, Huazhong Agricultural University / Key Laboratory of Crop Ecophysiology and Farming System for the Middle Reaches of the Yangtze River, Ministry of Agriculture and Rural Affairs, Wuhan 430070, Hubei, China;2Qinghai Academy of Agricultural and Forestry Sciences, Xining 810016, Qinghai, China;3National Agricultural Technology Extension Service Center, Beijing 100125, China;4Huanggang Academy of Agriculture Science, Huanggang 438000, Hubei, China;5Jingzhou Academy of Agriculture Science, Jingzhou 434000, Hubei, China;6Xiangyang Academy of Agriculture Science, Xiangyang 441057, Hubei, China
High yield and lodging resistance are two goals in rapeseed production that are hard to reconcile, which restrict severely the further improvement of economic performance. This contradiction is particularly prominent in the high-density and mechanize farming mode. Due to maternal inheritance, there are obvious differences in yield and lodging resistance related traits among reciprocal hybrids, but the effects of planting density on these key traits are not clear. Here, Huahang 901 and Zheyou 50, which are conventional rapeseed varieties and their reciprocal hybrids, were planted with gradient density. The yield and lodging related traits were measured and the heterosis of each trait was calculated. In order to provide the basis for the selection of parents under the high-density and direct seeding farming mode, the effects of planting density on yield and lodging related traits of parents and reciprocal hybrids were investigated. The results showed that the actual yield and economic coefficient of two parents and their reciprocal hybrids were first increased and then decreased with the increase of the planting density. Interesting, the dynamic change pattern and the values of both traits of reciprocal hybrids under gradient density were well consistent with those of paternal parent. The variation of lodging index with increased density at two sites was different, but the values and the dynamic change patterns of reciprocal hybrids were consistent with that of the maternal parent. In 2017 in Xining, the actual yield of reciprocal hybrids had obvious heterosis and Huahang 901×Zheyou 50 showed higher heterosis than that of Zheyou 50×Huahang 901. Meanwhile, the Huahang 901×Zheyou 50 had greater heterosis of lodging resistances, but there was no such heterosis in Zheyou 50×Huahang 901. In conclusion, to obtain hybrids rapeseed varieties with high yield lodging resistance and dense planting under the high-density and direct seeding farming mode, it was better to select those varieties with high-yield as paternal parents and those varieties with strong lodging resistance as maternal parents.
rapeseed; reciprocal hybrids; planting density; yield; lodging
10.3724/SP.J.1006.2021.04253
本研究由國家重點研發計劃項目“大田經濟作物優質豐產的生理基礎與調控”(2018YFD1000900)資助。
This study was supported bythe National Key Research and Development Program of China “Physiological Basis and Agronomic Management for High-quality and High-yield of Field Cash Crops” (2018YFD1000900).
王晶, E-mail: wangjing@mail.hzau.edu.cn
E-mail: davidlou@webmail.hzau.edu.cn
2020-11-23;
2021-01-21;
2021-02-19.
URL: https://kns.cnki.net/kcms/detail/11.1809.S.20210219.1616.013.html
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
