DNA加雙氧酶TET1在高壓誘導心肌纖維化中的作用研究
2021-07-09 03:36:04李路安吳清蕊鄺素娟張黔桓
中國藥理學通報 2021年7期
關鍵詞:水平
李路安,吳清蕊,李 倩,饒 芳,鄺素娟,楊 慧,張黔桓,張 斌
[1.華南理工大學醫學院,廣東 廣州 510006;2.廣東省心血管病研究所心血管內科,廣東 廣州 510080;3.廣東省人民醫院(廣東省醫學科學院)醫學研究部臨床藥理重點實驗室,廣東 廣州 510080]
心臟壓力負荷增高是多種心臟疾病共同存在的現象,這類心臟疾病常見包括原發/繼發性高血壓、心臟瓣膜疾病、原發/繼發性心肌病以及肺動脈高壓等,長時間的心臟壓力負荷增高會引發心肌纖維化,造成心臟不可逆的損害,最終危害生命健康[1]。心肌纖維化是一種心臟受到異常刺激后不斷發生損傷與修復的病理改變,主要是由于心肌成纖維細胞(CFs)增殖轉化成肌成纖維細胞并大量分泌細胞外基質(ECM),最終ECM過度沉積所導致的[2]。ECM的主要成分為膠原蛋白,其中I型與Ⅲ型膠原蛋白(COL-1、COL-3)在心肌組織中占總膠原蛋白含量的90%以上,COL-1和COL-3比例異常升高會使得心肌變硬,最終出現收縮與舒張功能障礙[3]。
目前研究表明,轉化生長因子β(transforming growth factor beta,TGF-β)參與纖維化的發生發展,是治療纖維化的重要靶點。此外,TGF-β可以促進CFs分泌膠原蛋白,引起心肌纖維化[4],且在壓力負荷誘導的心肌纖維化中也發揮著重要的作用[5]。
DNA加雙氧酶TET(Ten-eleven translocation)家族是一種去甲基化相關的DNA加雙氧酶,它可以通過將5-甲基胞嘧啶(5-mC)氧化成5-羥甲基胞嘧啶(5-hmC)、5-胞嘧啶甲酰(5-fc)和5-胞嘧啶羧基(5-cac),5-fc與5-cac最終被胸腺嘧啶DNA糖基化酶(TDG)識別并消化,實現DNA去甲基化[6]。目前認為,DNA去甲基化與基因激活有關,研究發現TET2可以通過調控TGF-β調節區域的去甲基化實現TGF-β的激活,進而影響糖尿病腎病的發生發展[7]。另外也有研究報道DNA去甲基化參與主動脈縮窄小鼠心肌纖維化的發生發展[8];但在高壓負荷下TET家族參與心肌纖維化的病理機制尚未明確。因此,本研究將Wistar及自發性高血壓大鼠(spontaneous hypertension rat,SHR)作為動物模型、以高壓誘導的原代SD乳大鼠心肌成纖維細胞(NRCFs)作為細胞模型,探討高壓負荷下心肌纖維化的發生機制是否與TET家族對TGF-β的甲基化修飾相關,旨在為高壓負荷下防控心肌纖維化的發生提供新的治療靶點,并為改善心肌纖維化患者的心功能提供新的治療方案。
1 材料
1.1 實驗動物出生7周的♂ SHR和Wistar大鼠,購自北京維通利華實驗動物技術有限公司,生產許可證為SCXK(京)2016-0006。出生0-3 d的SD乳大鼠,不限雌雄,購自南方醫科大學實驗動物中心,生產許可證號為SCXK(粵)2016-0041。動物質量檢測單位為廣東省實驗動物檢測所。動物實驗獲廣東省人民醫院(廣東省醫學科學院)倫理委員會批準,批準號No.GDREC2014095A。
1.2 主要試劑TET1(ab191698)、TET2(ab94580)、TET3(ab139311),COL-1(ab34710)、COL-3(ab7778)和5-hmC(ab214728)抗體,均購自Abcam公司;TGF-β(3711)抗體購自CST公司;GAPDH(60004-1)抗體購自Proteintech公司;Loding Buffer(9173)和TB Green? Premix Ex TaqTMⅡ(RR820A)購自TaKaRa公司;蛋白酶抑制劑(539131)和RIPA裂解液(20-188)購自Millipore公司;Lipo3000轉染試劑盒(L3000-015)購自Thermo公司;DMEM/F12培養基(C11330500BT)購自Gibco公司;BCA試劑盒(P0009)和DNA提取試劑盒(D0063)購自碧云天公司;TBS(AR0031)和PBS(AR0030)購自BOSTER公司;EpiMark?5-hmC 和5-mC分析試劑盒(E3317)購自NEB公司;Triton X-100(X100-100ML)購自Sigma公司;sh-RNA-TET1購自吉凱基因。
2 方法
2.1 動物實驗
2.1.1動物造模及心臟標本采集 SHR是自發性高血壓大鼠,其血壓隨鼠齡增高而不斷增高,并在12周左右確立高血壓,本研究將出生7周的雄性Wistar大鼠(n=6)作為正常血壓對照組,性別周齡匹配的SHR(n=6)作為高血壓組,飼養于恒定的標準室溫、濕度和光照周期下,自由進食標準鼠食及自來水,飼養8周后,根據SHR與Wistar大鼠的血壓明確SHR造模是否成功。血壓的測量采用無創血壓計測定大鼠尾部動脈血壓,測量3次并取其均值為1次血壓。
將造模成功的SHR與Wistar大鼠稱重,腹腔注射戊巴比妥鈉(30 mg·kg-1)麻醉,待麻醉后頸椎脫臼法處死大鼠,快速取出整顆心臟標本,冰上快速分離出大鼠左心室組織,并剪取綠豆大小的左心室組織置入預冷的多聚甲醛(40 g·L-1)中4 ℃保存,用于后續的HE及Masson染色,剩余一部分心肌組織置于-80 ℃ 保存備用。
2.1.2Western blot 心臟組織總蛋白提取:剪取適量組織塊,PBS清洗后加入RIPA裂解液(含10 g·L-1的蛋白酶抑制劑),冰上破碎裂解20-30 min,12 000 r·min-14 ℃ 離心15 min,取上清為組織總蛋白。采用BCA法測蛋白濃度,按照20 μg蛋白上樣量配置上樣緩沖液,經變性、電泳、電轉,封閉后用相應的一抗(1 ∶1 000稀釋)4 ℃ 孵育過夜,用與一抗相同來源的二抗(1 ∶5 000稀釋)室溫孵育1 h,最后使用ECL法顯影目的條帶,Image J軟件測定條帶灰度值,并計算目的條帶與內參GAPDH灰度值的比值。
2.1.3HE和Masson染色 將Wistar大鼠和SHR的左心室心肌組織在多聚甲醛(40 g·L-1)中固定48 h后,進行石蠟包埋,切片,HE和Masson染色,光學顯微鏡下觀察心肌形態及心肌纖維膠原的改變。
2.2 細胞實驗
2.2.1原代NRCFs分離及造模 快速取出0-3 d的SD乳大鼠心臟,用酶解法將心肌組織消化成單個細胞,再利用心肌細胞與NRCFs貼壁時間的差異分離出NRCFs,使用的酶液為含有胰酶(0.8 g·L-1,溶劑為PBS溶液)和膠原酶(0.8 g·L-1,溶劑為PBS溶液)的混合酶。用含10% 胎牛血清的DMEM/F12培養基培養NRCFs,并將細胞置于37 ℃、5% CO2培養箱中培養,待細胞密度達到80%-90% 時進行細胞傳代。選取P2-P5代NRCFs進行干預。使用我們自行研制的高壓裝置,可提供并維持一定的壓力(專利號201420109263.1,中國),根據大鼠正常左心室的壓力負荷為90 mmHg-120 mmHg,設置0 mmHg為對照組、120 mmHg為正常壓力組、180 mmHg為高壓力組,處理24 h,并以NRCFs的纖維化相關蛋白(COL-1和COL-3)表達的增加來判斷心肌纖維化模型是否建立成功。
2.2.2原代NRCFs質粒轉染 采用Lipofectamine 3000試劑對NRCFs進行質粒轉染,分別設置Control+Neg組、180 mmHg+Neg組和180 mmHg+sh-TET1組。在細胞成長至80%-90% 的密度時,按125 μL ∶7.5 μL比例加入Opti-MEMTM培養基與 Lipofectamine 3000試劑,混勻;按125 μL ∶2.5 μg比例加入Opti-MEMTM培養基稀釋DNA,然后添加5 μL的P3000 試劑;按1 ∶1的比例在每管已稀釋的LipofectamineTM3000 試劑中加入稀釋的DNA,室溫孵育15 min,將DNA-脂質復合物加入到細胞中,轉染24 h換液,繼續轉染24 h,顯微鏡觀察細胞轉染效率。
2.2.3Western blot 細胞總蛋白提取:將處理好的細胞用PBS清洗殘留培養液,加入RIPA裂解液(含10 g·L-1的蛋白酶抑制劑),冰上靜置裂解20-30 min,收集裂解液,12 000 r·min-14 ℃離心15 min,取上清為細胞總蛋白。其余步驟同“2.1.2”。
2.2.4細胞免疫熒光 在貼壁的NRCFs中加入多聚甲醛(40 g·L-1)固定細胞15 min,PBS洗凈;加入 Triton X-100(2 g·L-1)通透細胞20 min,PBS洗凈;加入HCl(2 mol·L-1)變性細胞30 min,PBS洗凈;加入BSA(50 g·L-1)封閉細胞30 min,去除BSA,加入一抗(5-hmC,1 ∶500),置于避光濕盒中4 ℃ 孵育過夜,加入適量的熒光二抗(1:200)室溫孵育1 h;最后加入DAPI染細胞核,激光共聚焦顯微鏡觀察染色結果,Image J軟件測定目的細胞5-hmC的平均熒光強度。
2.2.55-mC與5-hmC的水平檢測 使用碧云天公司的DNA提取試劑盒提取NRCFs的DNA,將提取好的DNA使用NEB公司的EpiMark 試劑盒來分析5-mC與5-hmC的水平。這種方法是利用了同分異構體Mspl和Hpall對不同修飾的DNA甲基化敏感性來區分5-mC和5-hmC。將目的基因組DNA用T4-BGT處理,將所有的5-hmC葡萄糖化(5-ghmC),但是未修飾或含有5-mC的DNA不受影響。糖基化后使用Mspl和Hpall限制性內切酶消化相同的目的序列(CCGG),Hpall只裂解一個完全未修飾的位點,任何修飾(5-mC、5-hmC或5-ghmC)在任何一個胞嘧啶都將阻斷裂解;而Mspl識別和切割5-mC和5-hmC,但不識別和切割5-ghmC。
將不同限制性內切酶裂解的DNA樣品用熒光定量PCR來分析DNA片段的5-mC與5-hmC的變化。將目的DNA配置成10 μL的PCR反應體系,內含:5 μL TB Green?Premix Ex Taq,3.2 μL超純水,0.4 μL PCR正義引物,0.4 μL PCR反義引物,1 μL DNA模板。反應條件如下:95 ℃ 2 min,PCR反應40個循環(95 ℃ 25 s,58 ℃ 25 s,72 ℃ 25 s),72 ℃ 5 min,60 ℃ 25 s,95 ℃ 15 s。通過2-ΔΔCT法計算基因的相對表達水平,β-actin作為內參對照。TGF-β引物序列:Forward 5′-GCCGCGGATCCTCCAGAC A-3′, Reverse 5′-CAGACTCTGGGGCCTCGGA-3′,β-actin引物序列:Forward 5′-TGTCCCTGTATGCCTC TGGT-3′, Reverse 5′-GATGTCACGCACGATTTCC-3′。

3 結果
3.1 SHR心肌纖維化改變及TET1和TGF-β蛋白表達水平變化為了探索在體TET與TGF-β以及心肌纖維化的關系,選取Wistar與SHR大鼠心室肌組織來檢測心肌纖維化以及TET、TGF-β蛋白表達水平的改變。結果顯示:與Wistar大鼠相比,SHR大鼠血壓明顯升高(P<0.01,Tab 1),心室組織中的TET1、TGF-β、COL-1和COL-3的蛋白表達均增加(P<0.05,Fig 1A)。HE染色顯示心肌細胞間質增加、排列紊亂,心肌細胞呈現肥大狀態。Masson染色可見心肌組織間膠原纖維增多(Fig 1C)。
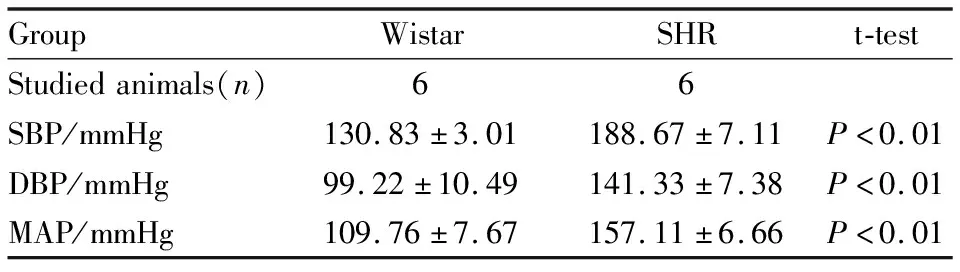
Tab 1 Comparison of blood pressure between Wistar rats and SHR
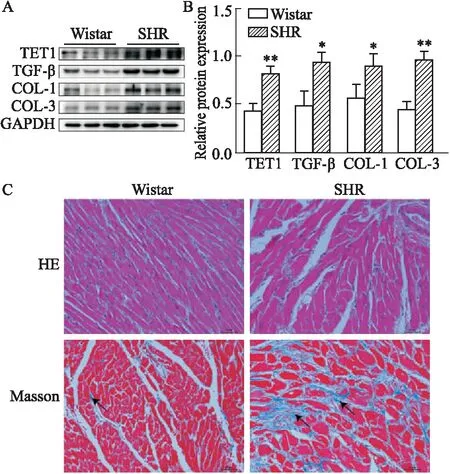
Fig 1 Expressions of TET1, TGF-β and fibrosis related protein in myocardium of Wistar rats and SHR
3.2 高壓對NRCFs中纖維化相關蛋白及TGF-β的表達水平的影響將P2-P5代心肌成纖維細胞置于不同的壓力(0、120、180 mmHg)中培養24 h,使用Western blot 檢測不同壓力梯度下COL-1和COL-3的表達,以及檢測促纖維化因子TGF-β的表達。結果顯示:與對照組相比,隨著120 mmHg與180 mmHg壓力梯度的增高,TGF-β蛋白表達水平以及COL-1和COL-3蛋白均逐漸升高,在180 mmHg壓力下達最高水平(P<0.01,Fig 2A);而與120 mmHg壓力組相比,180 mmHg壓力組的上述蛋白表達水平均有增加,但差異無統計學意義(P>0.05,Fig 2A)。
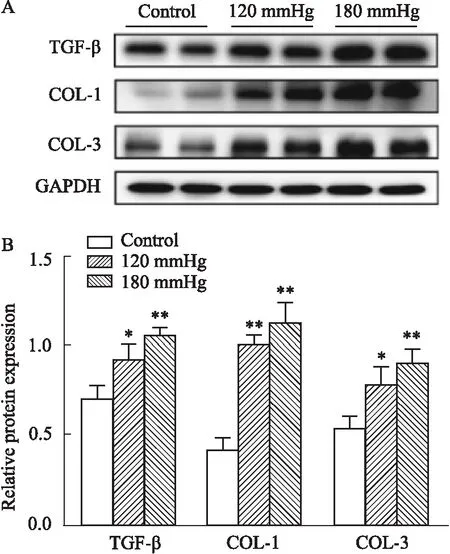
Fig 2 High pressure affected expression of fibrosis-related proteins and TGF-β in NRCFs
3.3 高壓處理NRCFs對TET家族蛋白表達及5-hmc水平的影響在高壓誘導NRCFs中纖維化相關蛋白高表達的基礎上,使用Western blot 檢測TET1、TET2和TET3的蛋白表達,以及使用免疫熒光檢測5-hmC的熒光強度。結果顯示:與對照組相比,壓力負荷組5-hmC的熒光強度明顯增加,并呈壓力依賴性(P<0.01,Fig 3C);去甲基化相關酶TET1蛋白表達隨著壓力梯度的升高而升高,但僅在180 mmHg組差異有統計學意義(P<0.05,Fig 3A),而TET2和TET3無明顯的變化(P>0.05,Fig 3A)。以上結果顯示,在高壓誘導心肌纖維化模型中,細胞呈現了去甲基化的趨勢,且這種改變與TET1表達升高相關,提示TET1在高壓誘導的心肌纖維化中可能發揮著重要的作用。
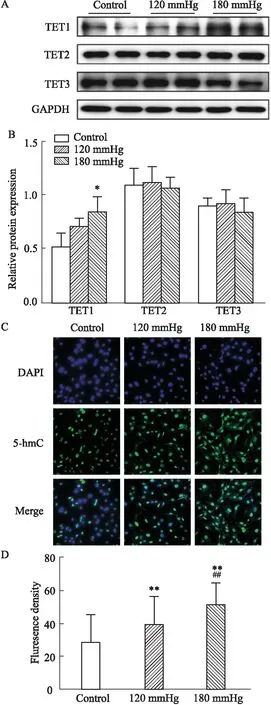
Fig 3 High pressure affected TET1 expression and 5-hmC level in NRCFs
3.4 敲低TET1對高壓誘導的5-hmC水平的影響為了進一步明確高壓狀態下5-hmC的水平增加與TET1的高表達相關,我們對180 mmHg壓力下的NRCFs予以TET1 shRNA質粒轉染敲低TET1(轉染效率見補充Fig 1),觀察5-hmC水平的變化。結果發現,TET1 shRNA質粒轉染后TET1表達水平降低(P<0.01,Fig 4A)并伴隨著5-hmC水平的降低(P<0.01,Fig 4C),說明TET1參與調控高壓狀態下5-hmC水平的變化。
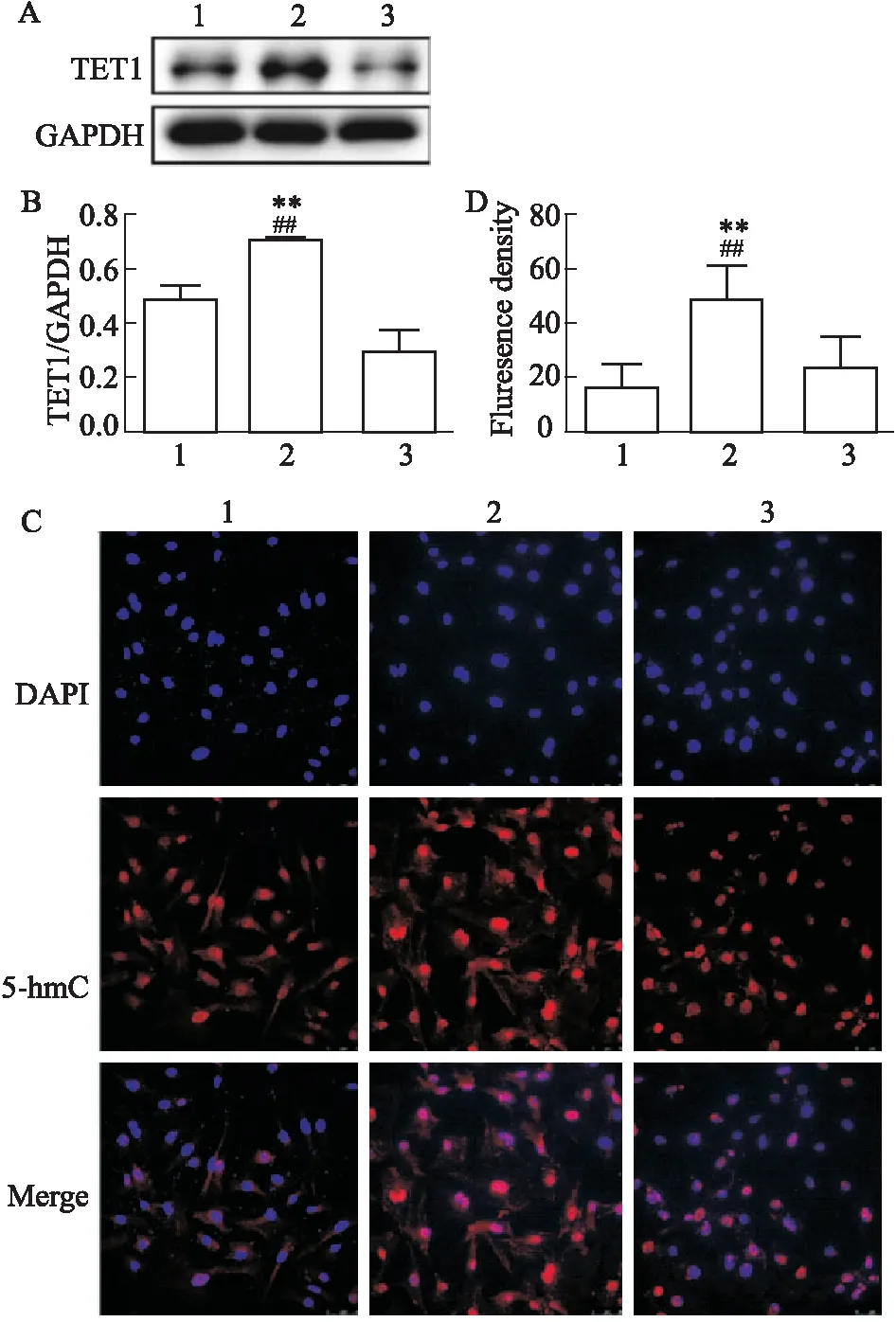
Fig 4 Knockdown of TET1 affected 5-hmC level under high pressure
3.5 敲低TET1對高壓誘導TGF-β啟動子區域5-hmC水平、TGF-β蛋白表達以及心肌纖維化相關蛋白表達的影響為了確定TET1在高壓相關的心肌纖維化中發揮關鍵作用,在高壓下敲低TET1的NRCFs中進一步檢測纖維化相關蛋白和TGF-β的表達,以及TGF-β啟動子區域5-hmC的水平。結果顯示,敲低TET1之后,TGF-β表達水平下降(P<0.01,Fig 5C),并伴隨著纖維化相關蛋白COL-1和COL-3表達降低(P<0.05,Fig 5C);此外,TGF-β啟動子區域5-hmC在敲低TET1后水平明顯降低(P<0.01,Fig 5B),而5-mC和5-hmC的總水平升高(P<0.05,Fig 5A),提示敲低TET1促使TGF-β啟動子區域甲基化。此部分研究顯示,高壓狀態下敲低TET1可以通過促進TGF-β啟動子區域甲基化從而降低TGF-β表達以及降低纖維化相關蛋白的表達。
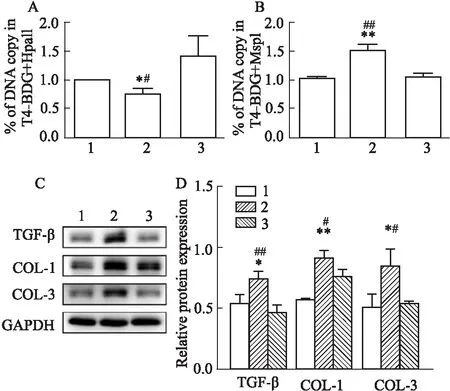
Fig 5 Knockdown of TET1 affected 5-hmC level in TGF-β promoter region, TGF-β expression and fibrosis related proteins induced by high pressure
4 討論
本研究結果表明,壓力負荷增高,DNA加雙氧酶TET1、5-hmC、TGF-β以及纖維化相關蛋白增加;高壓下敲低TET1可以降低TGF-β啟動子區5-hmC水平、增加5-mC和5-hmC總水平,以及降低TGF-β表達,抑制纖維化相關蛋白表達。提示TET1可能通過調控TGF-β啟動子區域去甲基化進而激活TGF-β表達,參與高壓負荷誘導的心肌纖維化。
心臟高壓負荷會通過持續的機械應力誘發心臟的一系列自我保護作用,包括心肌肥大以及心肌纖維化等,其中心肌纖維化以膠原沉積為主要特征,最終會導致心臟順應性下降,收縮與舒張功能障礙,心力衰竭。已有研究表明,機體內心血管系統承受的三種不同的力學刺激,剪切應力、牽張力和靜水壓[9];剪切應力是指血流對細胞表面產生的摩擦力[10]。牽張力是指血流導致血管擴張和回縮產生的力[11];靜水壓是指血流對單位面積血管壁的壓力(即血壓)[9]。我們認為高壓負荷所導致心臟的一系列病理改變,其主要的力學改變是高靜水壓。靜水壓升高進而導致心臟擴大,發生牽拉,導致牽張力的增加。因此,本研究使用自行研制的高靜水壓裝置,在體外給予一定的壓力負荷,以觀察高壓負荷導致心肌纖維化的病理機制。在本課題組已報道研究中,通過體外高壓裝置給與心房肌成纖維細胞20 mmHg及40 mmHg壓力梯度來模仿心房高壓狀態,并發現心肌纖維化相關因子水平隨壓力梯度升高,成功構建心房肌成纖維細胞心肌纖維化模型[12]。在本研究中,通過體外給與NRCFs 120 mmHg及180 mmHg的壓力來模仿心室的正常壓力和高壓狀態,發現NRCFs在120 mmHg及180 mmHg壓力處理24 h后,促纖維化因子TGF-β以及心肌纖維化相關蛋白COL-1和COL-3呈現穩定的梯度增加,以高壓180 mmHg更為明顯。
TGF-β作為已知的心肌纖維化發病重要靶點,主流研究聚焦于TGF-β與Smad家族蛋白之間的關系,認為TGF-β可以通過結合于細胞膜TGF-β的I型和Ⅱ型受體形成復合體,然后作用于胞質中的Smad家族蛋白并使其磷酸化活化,進而啟動纖維化的發生[13-14]。但是在心肌纖維化中何種機制能夠調控TGF-β還未得到很好闡明。本研究中,不論是細胞高壓模型還是SHR動物模型中,TGF-β表達均明顯升高,與目前主流研究一致,因此需進一步探索調控TGF-β的作用機制。
TET家族蛋白是DNA去甲基化最主要的因子,通過將5-mC氧化成5-hmC啟動去甲基化過程進而調控基因的表達[15]。研究表明,DNA去甲基化與心肌纖維化存在密切關系。如在升主動脈縮窄模型小鼠中,骨形成蛋白7通過促進 RASAL1啟動子去甲基化,促進RASAL1表達增加,進而減少了心肌纖維化[8];在SHR心臟中,也發現抑制DNA甲基化可以顯著減少心肌膠原蛋白沉積以及心肌細胞的大小[16]。雖然直接關注TET家族蛋白在器官纖維化中作用的研究不多,但在硬皮病患者的成纖維細胞中,TET1的mRNA水平明顯高于正常成纖維細胞,5-hmC水平明顯升高,總體甲基化水平降低[17],提示TET1與硬皮病患者的纖維化有著密切關系。本研究中,在120 mmHg與180 mmHg壓力造模成功的基礎上,發現5-hmC的水平梯度上升,TET1蛋白表達梯度升高,而TET2和TET3并無明顯改變,并在SHR中也發現了TET1表達升高,提示TET1可能通過調控DNA去甲基化參與高壓誘導的心肌纖維化。進一步在180 mmHg壓力狀態下敲低NRCFs的TET1,發現5-hmC水平和TGF-β蛋白表達均下降,且伴隨著COL-1和COL-3蛋白表達下降,說明TET1在高壓誘導的TGF-β參與的心肌纖維化中發揮著重要作用。有研究發現,在糖尿病腎病中,TET2可以通過調節TGF-β調節區域的去甲基化進而激活TGF-β的表達來調控糖尿病腎病的發病。本研究中,在敲低TET1的基礎上,發現TGF-β啟動子區域5-hmC水平降低,而5-mC及5-hmC總水平升高,說明敲低TET1后TGF-β啟動子區域中甲基化水平增高,抑制了高壓狀態下被激活的TGF-β,進一步說明在高壓誘導的心肌纖維化模型中,TET1可以調控TGF-β啟動子區域的去甲基化進而調控TGF-β的表達。
綜上所述,本研究初步證實TET1可能通過調控TGF-β啟動子區域的去甲基化來激活TGF-β參與高壓誘導的心肌纖維化。本研究的創新點在于將DNA去甲基化相關酶TET1與TGF-β參與的高壓誘導的心肌纖維化關聯起來,旨在探索高壓誘導的心肌纖維化的發生機制,尋找高壓誘導心肌纖維化的潛在藥物治療靶點。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
