橡膠樹葉片衰老相關基因HbLEA3 的克隆與表達分析
2021-07-08 08:32:20郭運玲
西南林業大學學報 2021年4期
鄒 智 郭運玲 孔 華
(中國熱帶農業科學院熱帶生物技術研究所/海南省南繁生物安全與分子育種重點實驗室,海南 海口 571101)
胚胎發育晚期豐富(LEA)蛋白是一類高度親水的小分子蛋白,因其最早發現于棉花胚胎發育后期的子葉而得名,他們廣泛參與生物體的滲透調節和脅迫應答[1]。在植物中,該類蛋白成員眾多,其根據特有的Pfam 結構域可分為8 大家族,即LEA_1、LEA_2、LEA_3、LEA_4、LEA_5、LEA_6、DHN(dehydrin)和SMP(seed maturation protein)[2-3]。與其他家族相比,有關LEA_3 的研究相對較少。該家族早期命名為LEA5 或D?73,在模式植物擬南芥(Arabidopsis thaliana)中報道的51 個LEA 中,有4 個可歸為該家族,即AtDI21(AT4G15910)、AtSAG21/AtLEA5(AT4G-02380)、AtLEA3(AT1G02820)和AT3G53770[2]。亞細胞定位分析表明,除AtLEA3 胞質定位外,其他3 個蛋白都定位在線粒體;組織特異性蛋白組學分析顯示,AtDI21 和AtLEA3 主要分布于花中,而AtLEA5 分布于花和根中[4]。這些基因在正常生長條件下的表達豐度都比較低,但逆境脅迫可誘導AtDI21和AtLEA5的表達,其中,AtLEA5還與葉片衰老相關[5-7]。深入研究發現,過表達AtLEA5可提高酵母對氧化脅迫的耐受能力,而在擬南芥中的過表達則可促進開花和植株衰老,初生根的長度和地上部分的生物量也隨之減少[8-9]。雖然如此,與擬南芥相比,其他植物中有關LEA_3家族基因的研究還鮮見報道,尤如橡膠樹(Hevea brasiliensis)等非模式木本植物。
橡膠樹隸屬于大戟科,最早起源于南美的亞馬遜河流域,由其生產的天然橡膠是一種重要的工業原料和戰略物資[10-12]。與原產地相比,我國植膠區地處熱帶北緣,除病蟲害外,寒害、旱害和風害等嚴重影響我國橡膠樹的分布、生長以及橡膠產量[13-15]。因此,開展橡膠樹逆境生物學研究、挖掘抗逆相關基因具有重要的理論意義和應用價值。在前期的研究中,研究組通過利用HbCAB1、HbSAG12H1等標記基因成功構建了成熟和衰老葉片的轉錄組文庫[14,16-18],從中篩選到一個在衰老葉片中顯著上調的LEA3類基因,本文重點報道該基因的序列特征及其在葉片發育過程中和逆境脅迫下的轉錄調控模式。
1 材料與方法
1.1 實驗材料
供試材料為1 年齡的熱研7?33?97 組培苗,古銅期、變色期、淡綠期、成熟期、衰老早期和衰老中期等6 個典型發育時期葉片的采集詳見文獻[14,17]。與深綠的成熟期葉片相比,衰老期葉片在外觀上呈現出不同程度的黃化。為保持材料的一致性和可重復性,研究將衰老早期和衰老中期葉片中的葉綠素含量分別定義為成熟期葉片的15%~25% 和45%~55%[17]。脅迫因子和逆境相關植物生長調節劑的處理均采用成熟期的葉片,分別為4 ℃低溫、30% PEG 6000、250 mmol/L 氯化 鈉(NaCl)、50 μmol/L 過氧化氫(H2O2)、50 μmol/L 脫落酸(ABA)、50 μmol/L 乙烯利(乙烯釋放劑)、50 μmol/L 茉莉酸甲酯(MeJA)、100 μmol/L 水楊酸(SA),采集處理0、3、6、12、24 h 后的樣品,液氮速凍后置于?80 ℃超低溫冰箱保存備用。橡膠樹的基因組和轉錄組數據下載于NCBI 數據庫。酶、試劑盒等各種生化試劑和實驗耗材詳見文獻[19]。
1.2 分析方法
1.2.1 基因克隆與序列分析
HbLEA3的轉錄本序列來源于研究組從頭組裝的轉錄組數據庫,序列全長1096 bp,其中包含一個303 bp 的開放讀碼框(ORF)、235 bp 的5′UTR 以及558 bp 的3′ UTR。BLASTn 分析表明,該序列對應于熱研7?33?97 基因組Scaffold0677[20]反向鏈的491468 到492657,除編碼區存在一個94 bp 的內含子外,未發現堿基差異。為實驗克隆該基因,研究在基因編碼區的兩側設計引物對HbLEA3F(5′?AGC TAC TAT ATC TGA GAC TTA GTG G?3′)和HbLEA3R(5′?TGA TAA CGA TGT AGC CAT GTA AGC?3′)。樣品總RNA 的提取、反轉錄、基因的克隆、常規測序以及序列分析參照文獻[16,19]。
1.2.2 基因的表達分析
熒光定量分析以18 SrRNA 作為內參,具體參照文獻[12,15],引物序列分別為18SF(5′?GCT CGA AGA CGA TCA GAT ACC?3′)/18SR(5′?TTC AGC CTT GCG ACC ATAC?3′)和HbLEA3Fq(5′?TTA GCG GAG AAG AAG GTG ACG?3′)/HbLEA3Rq(5′?CTC AGG AAT CCA GTT GCC AGT?3′)。實驗采用大連寶生物SYBR?Green Mix 和熱電Type 5100 實時定量PCR 儀 進行,反應程序為:95 ℃預變性30 s,接著是95 ℃變 性5 s 和60 ℃延 伸30 s(40 個循環),最后60 ℃延伸30 s。每個樣品至少3 次生物學重復,使用2ΔΔt法計算基因的相對表達值,并用SPSS 進行單因素方差分析。
2 結果與分析
2.1 基因克隆
根據前期的轉錄組測序結果,相比成熟期葉片,HbLEA3在衰老葉片中的表達豐度較高,因此,研究以衰老中期葉片反轉錄的cDNA 為模板進行PCR 擴增。成功克隆到HbLEA3基因652 bp 的cDNA 序列(GenBank 登陸號KP244471),其中包含173 bp 的5′ UTR、303 bp 的ORF 和176 bp 的3′ UTR。該序列與轉錄組及熱研7?33?97基因組的對應區域完全一致,進一步證實該基因確實存在一個內含子。值得注意的是,將序列與RRIM600 基因組[21-22]進行比對發現存在7 個堿基變異,其中有5 個位于編碼區(圖1),而這導致了4 個氨基酸的變異。BLASTn 分析顯示,該基因在公共數據中至今還沒有表達序列標簽(EST)。
2.2 基因的生物信息學分析
2.2.1HbLEA3編碼蛋白 的理化特 性、亞細胞定位及保守結構域
HbLEA3編碼區的GC 含量為41.91%,編碼100 個氨基酸,預測的理論分子量(Mw)為11.61 kDa,等電點(pI)為9.07,不穩定系數(II)為32.37,脂肪族指數(AI)為78.10,GRAVY 為?0.498。在構成蛋白的20 種氨基酸中,亮氨酸(Leu)和賴氨酸(Lys)的比例最高(12%);其次是丙氨酸(Ala)和蘇氨酸(Thr)(8%);而半胱氨酸(Cys)和組氨酸(His)的比例最低,僅為1%。與其他LEA 蛋白類似,HbLEA3 的親水氨基酸高達75%,其N 端表現為疏水性,而C 端表現為親水性(圖2a)。事實上,TargetP?2.0(http://www.cbs.dtu.dk/services/TargetP/)分析顯示,蛋白的前28 個殘基為線粒體信號肽。MotifScan(https://myhits.isb-sib.ch/cgibin/motif_scan)分析表明,蛋白的22~89 位為保守的LEA_3 結構域(圖1 和圖2b);53~56 位為潛在的環磷酸腺苷磷酸化位點;90~93 位為酪氨酸蛋白激酶II 磷酸化位點;24~26、44~46 和90~92 位為潛在的蛋白激酶C 磷酸化位點。這表明,HbLEA3 是一個隸屬于LEA_3 家族、受磷酸化調控的高度親水的線粒體定位蛋白。

圖1 HbLEA3 的cDNA 序列及其與RRIM600 基因組中相應基因序列的比較Fig.1 Comparison of the cDNA sequence of HbLEA3 with its gene sequence in the RRIM600 genome
2.2.2 多序列比對與進化分析
同源分析表明,HbLEA3 與木薯(Manihot esculenta)、蓖麻(Manihot esculenta)、麻瘋樹(Jatropha curcas)和可可樹(Theobroma cacao)中同源蛋白的相似性分別為75.0%、67.6%、66.0%和61.8%;在模式植物擬南芥和水稻中的多個成員中,HbLEA3 與AtLEA3 和OsLEA10 的相似性最高,分別為36.0%和41.1%。與上述蛋白相比,HbLEA3 擁有較長的N 端(11 個殘基,圖2b)。進化分析表明,HbLEA3 與白云杉EMB12[23]聚在同一支(圖2c),兩者相似性為35.0%,暗示HbLEA3 的直系同源基因至少在被子與裸子植物分化之前就已經產生,并且在擬南芥中發生了丟失。
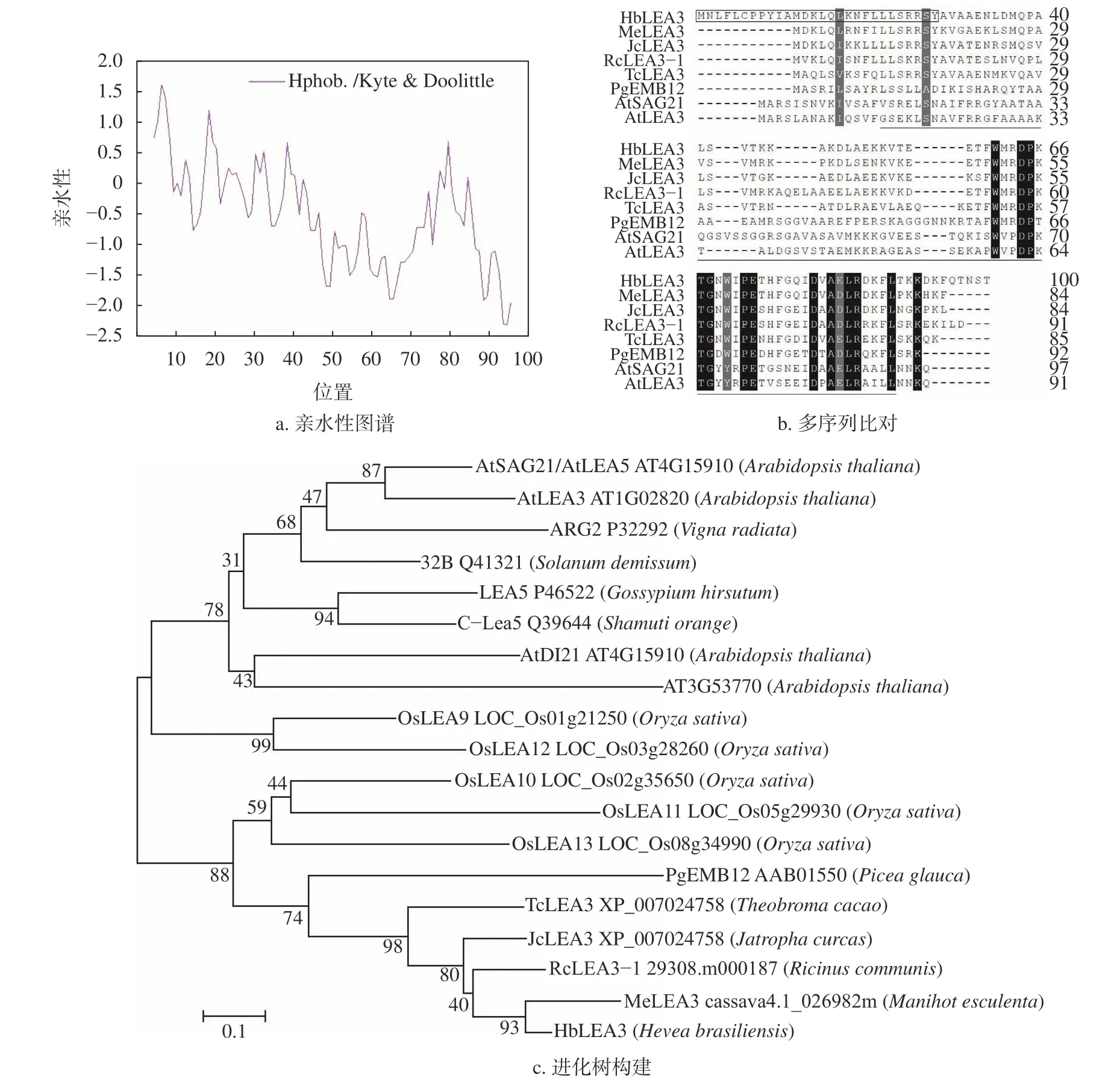
圖2 HbLEA3 蛋白的親水性圖譜、多序列比對與進化分析Fig.2 Hydropathy plot,multiple sequence alignment,and phylogenetic analysis of HbLEA3
2.3 HbLEA3 基因的表達模式分析
為了解HbLEA3的表達特性,研究進一步搜索了公共數據中乳管、樹皮、葉片、雌花、雄花、根和種子等不同組織的轉錄組數據,基于FKPM 的定量分析表明該基因主要在葉片中表達。為此,研究首先分析了基因在不同發育時期葉片中的表達模式,發現HbLEA3的表達水平隨葉片的成熟而穩步上升,但在衰老早期短暫下調,隨后在衰老中期顯著上調(圖3)。
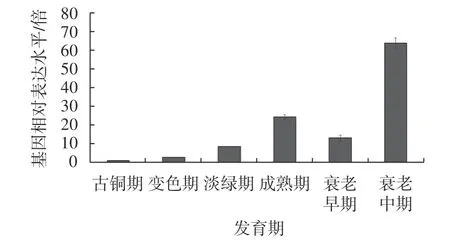
圖3 HbLEA3 在不同發育時期葉片中的表達模式(將對照設為1,以差異倍數作圖)Fig.3 Expression profile of HbLEA3 during leaf development
為揭示HbLEA3的轉錄調控模式,研究進一步分析了低溫、PEG、NaCl、H2O2、ABA、乙烯、MeJA 和SA 對基因表達的影響,結果表明,這些因子均可顯著誘導HbLEA3的表達,但響應模式存在一定的差異:在低溫、PEG 和H2O2脅迫下,主要表現為先升后降再升的趨勢;在NaCl和MeJA 脅迫下,呈單峰型;在乙烯、ABA 和SA 脅迫下,主要表現為先降后升趨勢(圖4)。
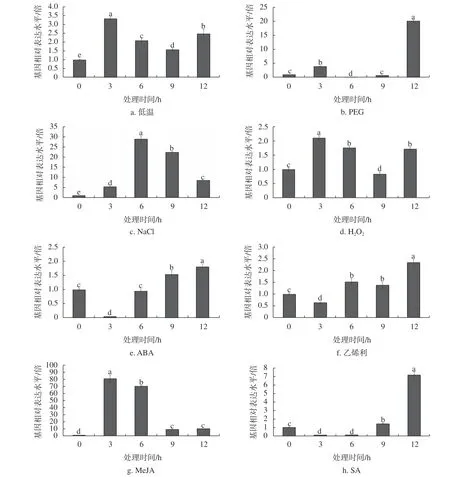
圖4 HbLEA3 對主要脅迫因子和逆境激素的響應模式Fig.4 Expression patterns of HbLEA3 upon several hormone and abiotic stress treatments
3 結論與討論
LEA 蛋白作為一類與植物逆境適應密切相關的明星分子而被廣泛關注[2-4]。在橡膠樹中,先前報道了5 個LEA類基因,其中3 個屬于LEA_2 家族[25-27],另外2 個屬于DHN 家族[28-29]。本研究報道了一個LEA_3 家族成員,命名為HbLEA3。該基因含有1 個內含子,預測編碼100 個氨基酸,蛋白含有1 個保守的LEA_3 結構域,堿性、高度親水。與其他同源蛋白相比,HbLEA3 擁有一延伸的N 端。由于橡膠樹與麻瘋樹、蓖麻、木薯同屬大戟科,且橡膠樹和木薯在與蓖麻和麻瘋樹分化之后共享了一次叫做ρ 的全基因重復事件(3900~4700 萬年前)[30-32],這表明HbLEA3極可能通過近期的堿基突變額外獲得了N 端的這11 個氨基酸。與多數擬南芥LEA3 蛋白類似[4],HbLEA3 預測定位在線粒體。
包含白云杉(Picea glauca)PgEMB12、4 個擬南芥LEA3 蛋白和5 個水稻(Oryza sativa)LEA3 蛋白的進化分析表明,LEA_3 家族至少含有3 個亞類,而HbLEA3 和PgEMB12[24]所處的亞類在裸子植物與被子植物分化之前就已形成,后期在不同物種中出現了不同的進化模式,如在擬南芥中發生了丟失,而在水稻中出現了較大水平的擴張(存在3 個成員)。目前有關LEA_3 家族的研究主要集中在4 個擬南芥LEA3 蛋白所處的分支,如綠豆(Vigna radiata)的Arg2[33]、棉花(Gossypiumspp.)的LEA5/D?73[34]、沙姆提橙(Citrusspp.)的C?Lea5[35]和馬鈴薯(Solanum tuberosum)的32B[36],他們與下胚軸的伸長、塊莖的發育以及非生物脅迫響應相關。與AtLEA5類似,HbLEA3在衰老葉片中高水平表達,而其穩步上升的表達模式暗示該基因可能參與了葉片的發育。考慮到低溫、PEG、NaCl、H2O2、ABA、乙烯、茉莉酸和水楊酸等脅迫因子或逆境相關植物生長調節劑均可誘導葉片衰老,研究深入分析了他們對HbLEA3的轉錄調控模式,事實證明這些因子均可誘導HbLEA3的表達,雖然在某些時間點也存在抑制作用。有趣的是,HbLEA3對這8 種因子表現3 種主要的響應模式,先升后降再升型包括低溫、PEG 和H2O2,單峰型包括NaCl和MeJA,先降后升型包括乙烯、ABA 和SA。事實上,除AtDI21和AtLEA5外,PgEMB12受ABA和PEG 誘導[24];Arg2受IAA、機械損傷和熱擊誘導[33];C?Lea5受NaC1、干旱和熱擊誘導[35];Os-LEA10和OsLEA12受ABA、PEG 和NaCl 誘導[37]。
綜上,研究表明HbLEA3是一個與橡膠樹葉片發育和脅迫響應相關的胚胎發育晚期豐富蛋白基因。雖然如此,更多的實驗如在原核生物和模式植物中過表達或進行內源敲除將有助于進一步闡明HbLEA3的生物學功能及調控通路。
