山楂酸調(diào)控STAT3磷酸化保護(hù)LPS致RAW264.7細(xì)胞炎癥反應(yīng)的研究
2021-07-06 02:12:38岳彩霞王文魁李桂蘭胡永婷
食品與機(jī)械 2021年6期
岳彩霞 陳 磊 王文魁 李桂蘭 胡永婷
(山西農(nóng)業(yè)大學(xué)動(dòng)物醫(yī)學(xué)學(xué)院,山西 太谷 030801)
山楂酸(maslinic acid,MA)屬于三萜酸類物質(zhì),存在于多種可食用的天然植物中,如山楂、紅棗、油橄欖和枇杷葉等[1-3],具有抗炎、抗氧化和抗2型糖尿病等多種生物學(xué)活性[2,4-6]。已有研究[2]表明,山楂酸在體內(nèi)具有抑制急性炎癥的作用,對(duì)角叉菜膠誘導(dǎo)的大鼠足腫脹有保護(hù)作用。體外研究[3,7]也發(fā)現(xiàn),MA的抗炎機(jī)制與抑制白介素6(Interleukin-6,IL-6)、白介素1β(Interleukin-1β,IL-1β)和腫瘤壞死因子α(Tumor necrosis factorα,TNF-α)等細(xì)胞因子的分泌有關(guān)。誘導(dǎo)型一氧化氮合酶(Inducible nitric oxide synthase,iNOS)是NO合成的調(diào)控因子,受細(xì)胞因子的誘導(dǎo)表達(dá)和調(diào)節(jié)[8-9]。Janus激酶/信號(hào)轉(zhuǎn)導(dǎo)與轉(zhuǎn)錄激活子(Janus kinase-signal transducer and activator of transcription,JAK-STAT)是重要的炎癥信號(hào)轉(zhuǎn)導(dǎo)途徑之一,能夠介導(dǎo)免疫應(yīng)答[10]。信號(hào)轉(zhuǎn)導(dǎo)和轉(zhuǎn)錄活化因子3(signal transducer and activator of transcription 3,STAT3)是脂多糖(lipopolysaccharide,LPS)誘導(dǎo)炎癥反應(yīng)的關(guān)鍵性調(diào)控因子[11]。
目前,MA調(diào)控炎癥的研究主要集中于體外動(dòng)物模型中抗炎效果的觀察及細(xì)胞模型中抑制炎癥因子的表達(dá)[2-3,12]。王樂(lè)旬等[7]研究發(fā)現(xiàn)MA可抑制LPS誘導(dǎo)的RAW264.7細(xì)胞炎癥反應(yīng)機(jī)制可能與調(diào)控NF-κB、STAT3和AKT的活性相關(guān),然而,MA在LPS誘導(dǎo)的RAW264.7細(xì)胞中是否通過(guò)調(diào)控STAT3的磷酸化發(fā)揮抗炎作用的具體分子機(jī)制尚不明確,研究擬利用課題組[13]前期建立的LPS誘導(dǎo)的RAW264.7細(xì)胞炎癥模型,在體外細(xì)胞水平觀察MA對(duì)LPS誘導(dǎo)的RAW264.7細(xì)胞炎癥反應(yīng)的影響,進(jìn)一步探討MA能否通過(guò)調(diào)控STAT3的磷酸化來(lái)減輕LPS誘導(dǎo)的RAW264.7細(xì)胞的炎性反應(yīng),為揭示MA的抗炎機(jī)制、綜合開(kāi)發(fā)利用MA作為天然抗炎藥物提供依據(jù)。
1 材料與方法
1.1 材料
1.1.1 細(xì)胞
RAW264.7細(xì)胞:中國(guó)科學(xué)院上海生命科學(xué)研究院細(xì)胞庫(kù)。
1.1.2 藥品與試劑
MA(CAS:4373-41-5):純度≥98%,美國(guó)Sigma公司;
脂多糖(LPS)和DCFH-DA:美國(guó)Sigma公司;
DMEM培養(yǎng)基:美國(guó)gibco公司;
標(biāo)準(zhǔn)胎牛血清:杭州四季青生物技術(shù)有限公司;
核蛋白提取試劑盒:北京索萊寶科技有限公司;
NO測(cè)試試劑盒:碧云天生物技術(shù)研究所;
IL-1β、IL-6和IL-10 ELISA試劑盒:上海江萊生物技術(shù)有限公司;
β-actin抗體:北京中杉金橋生物公司;
STAT3和iNOS抗體:武漢三鷹技術(shù)有限公司;
p-STAT3(Y705)、p-STAT3(S727)、JAK2、p-JAK2和SOCS3抗體:北京博奧森生物技術(shù)有限公司;
辣根過(guò)氧化物標(biāo)記山羊抗鼠和山羊抗兔IgG:北京康為世紀(jì)生物科技技術(shù)有限公司。
1.2 方法
1.2.1 細(xì)胞培養(yǎng) RAW264.7細(xì)胞使用含10%胎牛血清的DMEM培養(yǎng)基于37 ℃、5.0% CO2的細(xì)胞培養(yǎng)箱中。
1.2.2 給藥及分組 取對(duì)數(shù)生長(zhǎng)期的RAW264.7細(xì)胞,參照課題組[13]前期建立的LPS誘導(dǎo)的RAW264.7細(xì)胞炎癥模型進(jìn)行分組,分為對(duì)照組(以含10%胎牛血清的完全培養(yǎng)基培養(yǎng))、LPS模型組(以含1 μg/mL LPS的完全培養(yǎng)基培養(yǎng)24 h)和山楂酸組(不同濃度的山楂酸預(yù)處理2 h后,再用含1 μg/mL LPS和相應(yīng)濃度山楂酸的完全培養(yǎng)基繼續(xù)培養(yǎng)24 h),每組至少設(shè)3個(gè)重復(fù)。
1.2.3 NO含量的測(cè)定 取對(duì)數(shù)生長(zhǎng)期的細(xì)胞接種至96孔板,以不同濃度的LPS單獨(dú)刺激RAW264.7細(xì)胞24 h,并以不同濃度的山楂酸預(yù)作用細(xì)胞2 h后,再用含1 μg/mL LPS繼續(xù)培養(yǎng)進(jìn)行試驗(yàn),每組設(shè)6個(gè)重復(fù)孔。培養(yǎng)結(jié)束后,按照試劑盒說(shuō)明書(shū)進(jìn)行NO含量的測(cè)定。
1.2.4 ELISA檢測(cè)相關(guān)的細(xì)胞因子 取對(duì)數(shù)生長(zhǎng)期的RAW264.7細(xì)胞,藥物作用培養(yǎng)結(jié)束后,吸取每組細(xì)胞的上清液,加入96孔板,按照ELISA試劑盒的操作說(shuō)明,分別測(cè)定細(xì)胞因子IL-6、IL-1β和IL-10的含量,并進(jìn)行統(tǒng)計(jì)分析。
1.2.5 蛋白提取及Western blot試驗(yàn) 按照試驗(yàn)分組培養(yǎng)結(jié)束后,用現(xiàn)配的RIPA細(xì)胞裂解液裂解細(xì)胞30 min,提取各組RAW264.7細(xì)胞總蛋白,并按照核蛋白提取試劑盒說(shuō)明書(shū)提取相應(yīng)的核蛋白。取等量蛋白樣品進(jìn)行SDS-PAGE電泳,轉(zhuǎn)膜,5%的脫脂牛奶封閉,一抗4 ℃孵育過(guò)夜,二抗(HRP標(biāo)記)室溫孵育1 h后,暗室內(nèi)ECL顯色法曝光,進(jìn)行蛋白條帶定量分析。
1.2.6 ROS的測(cè)定 取對(duì)數(shù)生長(zhǎng)期的RAW264.7細(xì)胞接種于12孔板,按試驗(yàn)分組進(jìn)行細(xì)胞爬片,作用結(jié)束后,吸出培養(yǎng)基,用PBS清洗,每組加入含10 μmol/L DCFH-DA的無(wú)血清培養(yǎng)基,37 ℃避光孵育細(xì)胞30 min,棄掉培養(yǎng)基,PBS清洗。取出細(xì)胞爬片,在熒光顯微鏡(200×)下觀察熒光強(qiáng)度并拍照。
1.3 數(shù)據(jù)分析
采用Graphpad Prism 5軟件進(jìn)行統(tǒng)計(jì)分析,組間比較采用單因素方差,所有數(shù)值使用mean±SD表示,并設(shè)置多個(gè)重復(fù),P<0.05、P<0.01表示差異具有統(tǒng)計(jì)學(xué)意義。
2 結(jié)果與分析
2.1 MA對(duì)RAW264.7細(xì)胞NO生成量的影響
由圖1(a)可知,與正常對(duì)照組相比,NO生成量隨LPS刺激濃度的升高顯著增加(P<0.01);由圖1(b)可知,與LPS模型組相比,5,10,15,20,25 μmol/L MA干預(yù)后NO含量依次顯著降低(P<0.01),且呈一定的劑量依賴性,而30 μmol/L MA作用后NO含量有所增加,可能與MA的劑量較大有關(guān)。綜上,MA可以明顯抑制由LPS引起的RAW264.7細(xì)胞的NO生成量增加。
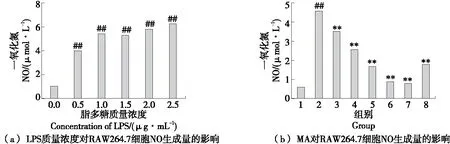
1.空白組 2.LPS組 3.5 μmol/L MA+LPS 4.10 μmol/L MA+LPS 5.15 μmol/L MA+LPS 6.20 μmol/L MA+LPS 7.25 μmol/L MA+LPS 8.30 μmol/L MA+LPS ##.與對(duì)照組相比P<0.01 **.與LPS組相比P<0.01
2.2 MA對(duì)RAW264.7細(xì)胞因子釋放和iNOS蛋白表達(dá)的影響
由圖2(a)~圖2(c)可知,LPS刺激RAW264.7細(xì)胞后IL-6、IL-1β和IL-10釋放顯著增加,與對(duì)照組相比差異具有統(tǒng)計(jì)學(xué)意義(P<0.05),MA可不同程度地拮抗RAW264.7細(xì)胞中IL-6和IL-1β的釋放,而與LPS模型組相比,MA作用可顯著增加RAW264.7細(xì)胞中IL-10的含量(P<0.01),且呈一定的劑量依賴性。研究表明,受LPS刺激的巨噬細(xì)胞會(huì)產(chǎn)生大量的炎癥因子如IL-1β、IL-6和TNF-α等,導(dǎo)致炎癥細(xì)胞浸潤(rùn),與炎癥的發(fā)生發(fā)展密切相關(guān)[14],而IL-10是巨噬細(xì)胞產(chǎn)生的一種重要的抗炎細(xì)胞因子,其表達(dá)對(duì)于炎癥在進(jìn)行免疫干預(yù)治療方面十分關(guān)鍵[15]。研究結(jié)果顯示,MA能明顯抑制LPS誘導(dǎo)的RAW264.7細(xì)胞IL-1β、IL-6的釋放,增加IL-10的含量,從而可以較好地抑制LPS誘導(dǎo)的巨噬細(xì)胞炎癥反應(yīng)。
iNOS是重要的炎癥因子,在LPS的刺激下iNOS表達(dá)增加進(jìn)而誘導(dǎo)NO的大量產(chǎn)生[16]。采用Western blot法檢測(cè)了LPS誘導(dǎo)后RAW264.7細(xì)胞中iNOS的表達(dá),如圖2(d)和圖2(e)所示,10,15,20 μmol/L MA可劑量依賴性地抑制LPS誘導(dǎo)的iNOS蛋白的表達(dá)(P<0.05),而20 μmol/L MA單獨(dú)刺激細(xì)胞,iNOS蛋白的表達(dá)與對(duì)照組無(wú)顯著差異(P>0.05)。NO是iNOS的產(chǎn)物,炎癥疾病發(fā)生時(shí)iNOS通常表達(dá)增加,因此抑制iNOS的表達(dá)對(duì)減輕炎癥反應(yīng)很重要[8-9,16]。在試驗(yàn)中,可以得出MA能通過(guò)抑制LPS誘導(dǎo)的iNOS表達(dá),進(jìn)而抑制NO的釋放。

1.空白組 2.LPS組 3.10 μmol/L MA+LPS 4.15 μmol/L MA+LPS 5.20 μmol/L MA+LPS 6.20 μmol/L MA ##.與對(duì)照組相比P<0.01 #.與對(duì)照組相比P<0.05 **.與LPS組相比P<0.01 *.與LPS組相比P<0.05
2.3 MA濃度對(duì)RAW264.7細(xì)胞STAT3的影響
STAT3在調(diào)控iNOS過(guò)程中發(fā)揮了重要的作用[17],利用Western blot檢測(cè)MA對(duì)LPS誘導(dǎo)的RAW264.7細(xì)胞STAT3的Y705和S727位點(diǎn)磷酸化產(chǎn)生影響,結(jié)果如圖3(a)所示,LPS刺激能快速誘導(dǎo)胞內(nèi)STAT3的酪氨酸和絲氨酸磷酸化,而經(jīng)不同濃度(5,10,15,20 μmol/L)的MA處理細(xì)胞后,胞內(nèi)p-STAT3(Y705)和p-STAT3(S727)蛋白的表達(dá)明顯被抑制(P<0.01),且呈劑量依賴性。已有研究[18]表明,STAT3的磷酸化有助于STAT3的二聚化并促進(jìn)其核轉(zhuǎn)位并發(fā)揮轉(zhuǎn)錄活性,由此猜測(cè)MA是否也抑制STAT3的入核,通過(guò)提取細(xì)胞核蛋白進(jìn)行檢測(cè),結(jié)果如圖3(b)所示,LPS刺激后STAT3快速入核,細(xì)胞經(jīng)MA處理后核內(nèi)p-STAT3(Y705)和p-STAT3(S727)蛋白表達(dá)顯著下降(P<0.05),并且呈劑量依賴性。由此得出,MA能下調(diào)LPS誘導(dǎo)的RAW264.7細(xì)胞p-STAT3的表達(dá)并抑制STAT3的核轉(zhuǎn)位,p-STAT3的表達(dá)量與MA濃度具有明顯的量效關(guān)系。

1.空白組 2.LPS組 3.10 μmol/L MA+LPS 4.15 μmol/L MA+LPS 5.20 μmol/L MA+LPS 6.20 μmol/L MA+LPS ##.與對(duì)照組相比P<0.01 **.與LPS組相比P<0.01 *.與LPS組相比P<0.05
2.4 MA刺激時(shí)間對(duì)RAW264.7細(xì)胞STAT3的影響
選用20 μmol/L的MA預(yù)孵細(xì)胞2 h,1 μg/mL LPS分別作用細(xì)胞0,1,2,4,8 h后,檢測(cè)細(xì)胞和核內(nèi)p-STAT3的蛋白表達(dá)變化。如圖4所示,LPS刺激4~8 h時(shí),MA能顯著降低胞內(nèi)STAT3的酪氨酸和絲氨酸磷酸化水平(P<0.05),且呈一定的時(shí)間依賴性,在LPS刺激8 h時(shí),MA能顯著降低核內(nèi)p-STAT3(Y705)和p-STAT3(S727)的表達(dá)(P<0.01),由此表明,LPS誘導(dǎo)的RAW264.7胞內(nèi)p-STAT3的表達(dá)與MA濃度之間具有一定的時(shí)間依賴性。
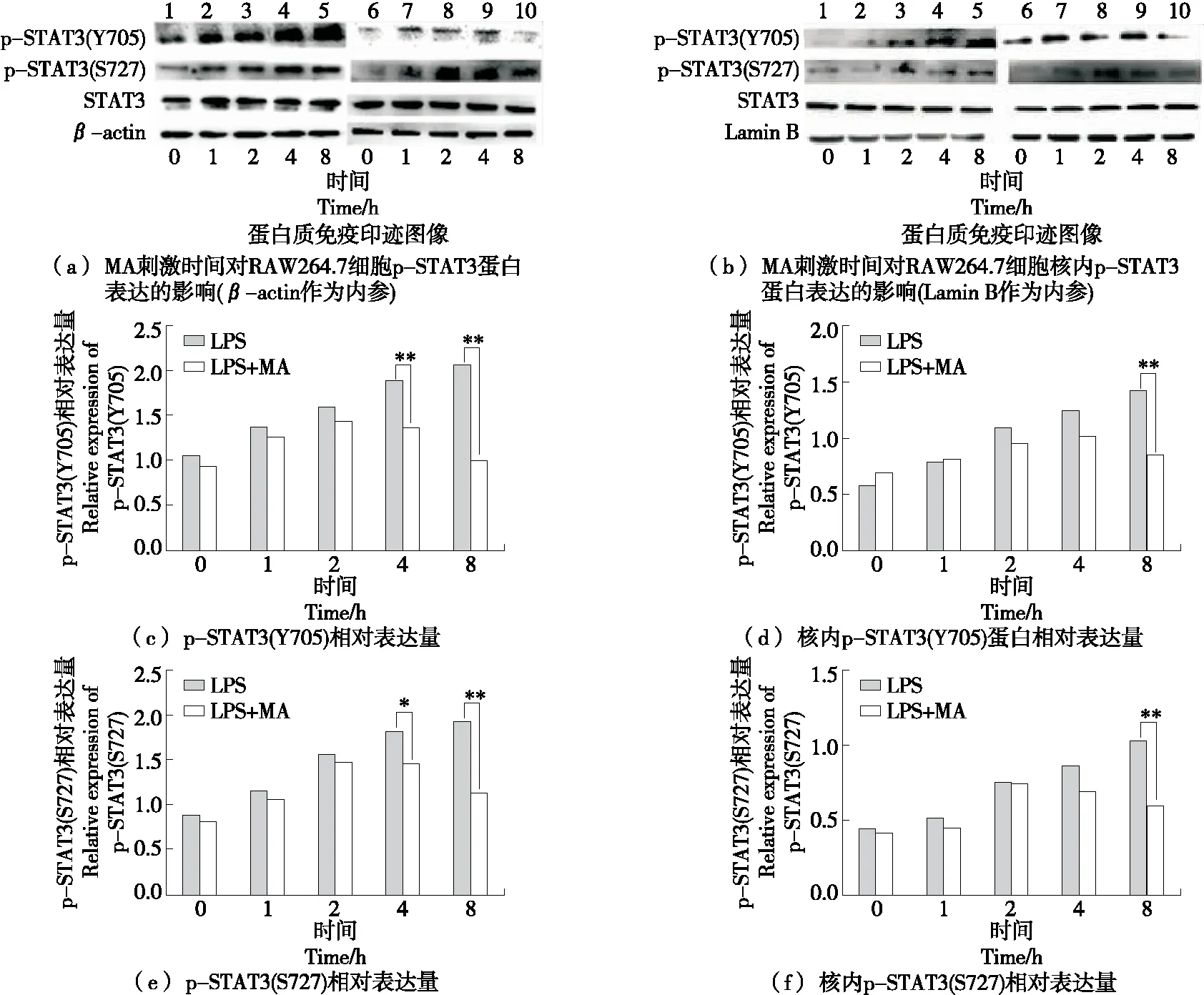
1.空白組 2.LPS 1 h 3.LPS 2 h 4.LPS 4 h 5.LPS 8 h 6.空白組 7.20 μmol/L MA+LPS 1 h 8.20 μmol/L MA+LPS 2 h 9.20 μmol/L MA+LPS 4 h 10.20 μmol/L MA+LPS 8 h **.與LPS組相比P<0.01 *.與LPS組相比P<0.05
研究[19]表明,STAT3在增殖、生長(zhǎng)、凋亡、免疫等重要的生理過(guò)程中發(fā)揮調(diào)控作用,而STAT3的異常活化可導(dǎo)致炎癥反應(yīng)與多種疾病相關(guān)。已有研究[20-21]證實(shí),STAT3作為典型的炎癥信號(hào)通路可以調(diào)節(jié)iNOS及相關(guān)促炎因子包括IL-1β、IL-6和TNF-α等的表達(dá)。由此可以推測(cè),MA可以通過(guò)對(duì)STAT3磷酸化的影響抑制其合成iNOS及對(duì)炎癥因子IL-1β和IL-6的釋放。
2.5 MA對(duì)RAW264.7細(xì)胞p-JAK2和SOCS3表達(dá)的影響
如圖5所示,與對(duì)照組相比,LPS模型組JAK2的磷酸化水平顯著升高(P<0.05),SOCS3蛋白表達(dá)水平顯著降低(P<0.01);與LPS模型組相比,不同濃度的MA可抑制p-JAK2的蛋白表達(dá)(P<0.05),而激活SOCS3蛋白表達(dá)(P<0.05),并均呈一定的劑量依賴性。結(jié)果表明,MA能抑制JAK2的磷酸化水平,激活SOCS3的蛋白表達(dá),與其抗炎機(jī)制相關(guān)。

1.空白組 2.LPS組 3.5 μmol/L MA+LPS 4.10 μmol/L MA+LPS 5.15 μmol/L MA+LPS 6.20 μmol/L MA+LPS ##.與對(duì)照組相比P<0.01 **.與LPS組相比P<0.01 *.與LPS組相比P<0.05
STAT3的激活依賴于JAK的活化,且有研究[22]表明,SOCS蛋白可以抑制JAK的活性。故檢測(cè)了MA對(duì)RAW264.7細(xì)胞中p-JAK2和SOCS3表達(dá)的影響,進(jìn)一步確定MA對(duì)JAK/STAT通路的作用,試驗(yàn)結(jié)果表明,MA可以通過(guò)調(diào)控JAK2/STAT3通路抑制LPS誘導(dǎo)的RAW264.7細(xì)胞炎癥反應(yīng)。
2.6 MA對(duì)RAW264.7細(xì)胞ROS水平的影響
細(xì)胞內(nèi)ROS的產(chǎn)生對(duì)于炎癥信號(hào)傳導(dǎo)發(fā)揮著重要的作用。采用DCFH-DA檢測(cè)山楂酸對(duì)RAW264.7細(xì)胞中ROS水平的變化。熒光顯微鏡(200×)下觀察如圖6所示,與對(duì)照組相比,LPS刺激后熒光強(qiáng)度顯著增強(qiáng)(P<0.01),不同劑量的山楂酸干預(yù)后,熒光強(qiáng)度依次減弱,表明ROS水平顯著被抑制(P<0.01),提示山楂酸可能通過(guò)降低RAW264.7細(xì)胞胞內(nèi)ROS的釋放發(fā)揮抗炎作用。

圖6 MA對(duì)LPS誘導(dǎo)RAW264.7細(xì)胞ROS水平的影響
在巨噬細(xì)胞中,ROS可以作為第二信使參與細(xì)胞的增殖、分化和成熟,但在致病因子作用下,ROS在機(jī)體中迅速大量表達(dá),通過(guò)激活胞內(nèi)多條信號(hào)通路包括JAK-STAT來(lái)誘導(dǎo)促炎因子的釋放和大量炎癥介質(zhì)的產(chǎn)生[23]。故測(cè)定了MA對(duì)RAW264.7細(xì)胞中ROS水平的影響,試驗(yàn)結(jié)果顯示,MA能抑制LPS誘導(dǎo)的RAW264.7細(xì)胞ROS的釋放,與李桂蘭等[5]發(fā)現(xiàn)的MA具有一定的抗氧化功能相一致。
由此,根據(jù)研究結(jié)果和之前的相關(guān)研究,提出山楂酸發(fā)揮抗炎保護(hù)作用可能的分子機(jī)制如圖7所示。
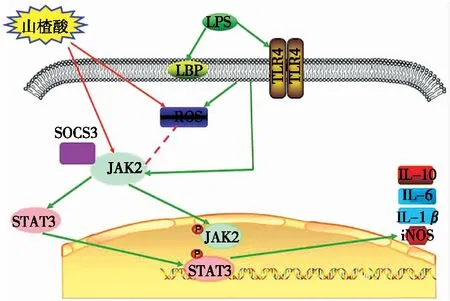
圖7 山楂酸發(fā)揮抗炎保護(hù)作用的分子機(jī)制
3 結(jié)論
在脂多糖誘導(dǎo)的RAW264.7細(xì)胞炎癥反應(yīng)模型中,山楂酸能通過(guò)抑制一氧化氮的生成、抑制白介素6和白介素1β釋放,增加白介素10的含量發(fā)揮抗炎保護(hù)作用,研究結(jié)果表明山楂酸可能通過(guò)抑制Janus激酶2/信號(hào)轉(zhuǎn)導(dǎo)與轉(zhuǎn)錄激活子3信號(hào)通路發(fā)揮抗炎作用,其機(jī)制還與抑制胞內(nèi)活性氧的釋放相關(guān)。但山楂酸如何通過(guò)活性氧調(diào)控Janus激酶/信號(hào)轉(zhuǎn)導(dǎo)與轉(zhuǎn)錄激活子信號(hào)通路,山楂酸是否還可以通過(guò)其他一些方式來(lái)抑制Janus激酶/信號(hào)轉(zhuǎn)導(dǎo)與轉(zhuǎn)錄激活子信號(hào)通路,相關(guān)的機(jī)制研究還有待進(jìn)一步的深入。
