特異性識別K1莢膜大腸桿菌的噬菌體PNJ1809-36生物學特性及全基因組分析
2021-06-30 13:57:18鞏倩雯李一昊于沛欣錢新杰王瑜欣戴建君
畜牧獸醫學報 2021年6期
鞏倩雯,李一昊,曾 頏,于沛欣,錢新杰,王瑜欣,戴建君,湯 芳
(南京農業大學動物醫學院 教育部動物健康與食品安全國際聯合實驗室 農業農村部細菌學重點實驗室, 南京 210095)
大腸桿菌(Escherichiacoli)為埃希菌屬(Escherichia)代表菌,它包括動物或人共生菌株和致病性菌株,致病性菌株又分為腸道致病菌株和腸道外致病菌株[1]。腸道致病性菌株有產腸毒素大腸桿菌、產志賀毒素大腸桿菌等,可以引起人和動物的腸炎和腹瀉[2],某些血清型甚至會導致死亡[3]。腸道外致病性菌株能引起人或動物的腸道外感染,如引起人或動物的尿路感染[4],新生兒腦膜炎及敗血癥以及禽的呼吸系統疾病從而導致禽的急性死亡。致病性大腸桿菌給人和動物健康造成危害,也給農業生產帶來了巨大的損失。
大腸桿菌有4種表面抗原:O、K、H和F抗原,分別為脂多糖抗原、莢膜多糖抗原、鞭毛抗原和菌毛抗原。4種表面抗原是對大腸桿菌進行分型的基礎,也是重要的毒力因子。K抗原是大腸桿菌重要的毒力因子之一[5],可能存在于莢膜、被膜或菌毛中。目前已發現的K抗原有80多種,其中K1莢膜抗原備受關注。引起新生兒細菌性腦膜炎的腸道外致病性大腸桿菌大多有K1莢膜,它能夠穿過血腦屏障致病,而K1莢膜則是主要致病因子[6]。具有K1莢膜的大腸桿菌能夠抵抗補體殺菌作用,增強其在腦微血管內皮細胞中的存活,從而減弱巨噬細胞對細菌的吞噬能力[7-8]。Mellata等[9]構建了K1基因缺失株,發現缺失株的增殖能力大大降低。目前,針對細菌性腦膜炎已經有治療方案,但經治療后也會伴有神經后遺癥及發育異常[10]。盡管使用了先進的抗生素,但與大腸桿菌K1引起的腦膜炎相關的疾病發病率和死亡率在過去幾十年中并沒有下降[11-12]。另外,由于近年來耐藥性大腸桿菌K1菌株的數量激增,死亡率可能會進一步增加[13]。
近年來,噬菌體治療被認為是能夠緩解細菌多重耐藥的有效方案之一。噬菌體具有分布廣泛、種類多樣、宿主特異性強等特點,可作為治療細菌感染的新型生物制劑。目前已經發現多種具有多糖專一性的噬菌體,可以特異性降解細菌胞外多糖,從而使細菌毒力下降。K1專一性噬菌體可以特異性降解K1莢膜的多聚唾液酸,從而使細菌毒力減弱[14]。同時,這些噬菌體可用于大腸桿菌的分型[15]。本研究分離得到可裂解K1莢膜大腸桿菌的噬菌體PNJ1809-36,并對其進行生物學特性和全基因組分析,期望為臨床疾病治療和細菌防控提供參考。
1 材料與方法
1.1 材料
1.1.1 細菌及樣品 大腸桿菌DE058及其他用于宿主譜檢測的大腸桿菌(表1)均為本實驗室保存。用于分離噬菌體的樣品為采集自青龍山養殖場的雞糞。
1.1.2 實驗儀器及試劑 參照《分子克隆實驗指南》第3版[16]配制LB液體培養基、LB固體培養基、半固體培養基以及SM緩沖液。LB培養基所用試劑購自北京索萊寶科技有限公司;NaCl、MgSO4·7H2O購自市科密歐化學試劑有限公司;酶標儀為TECAN NanoQuant PlateTM;電子透射顯微鏡為日立H7650。
1.2 方法
1.2.1 噬菌體分離和純化 參考Jamalludeen等[17]的方法對采集的雞糞樣品進行噬菌體分離與純化:用棉簽蘸取適量雞糞,放入含有SM液的EP管中4 ℃浸泡過夜。將該樣品5 000g離心10 min,隨后將上清用0.45 μm的濾器過濾,所得濾液在涂有大腸桿菌的平板上點樣,然后將平板置于37 ℃溫箱培養過夜,次日觀察平板上是否有空斑。如有空斑出現,則用雙層平板法對噬菌體進行純化:將宿主菌與該濾液各取100 μL混合,37 ℃靜置10 min后,將該混合液加入含0.5%瓊脂的LB半固體培養基中,混合均勻后鋪在LB固體平板上,37 ℃培養過夜,次日觀察噬菌斑。隨后用一次性無菌槍頭吸取單個空斑,放入含有SM液的EP管中4 ℃ 浸泡過夜,使其中的噬菌體充分釋放。將該SM液與宿主菌各取100 μL混合,再用雙層平板法培養噬菌體,重復該過程5次,直至平板上形成清晰、透明、大小一致的噬菌斑。取5 mL SM液加入該平板,4 ℃浸泡過夜,次日收集平板上的SM液,5 000g離心10 min,上清用0.22 μm濾器過濾,所得濾液即為純化的噬菌體,將其保存在4 ℃條件下備用。
1.2.2 電鏡觀察 將保存的噬菌體用雙層平板法富集并過濾,用2%的磷鎢酸(PTA)進行負染,使用透射電子顯微鏡觀察噬菌體的形態。
1.2.3 宿主譜分析 使用雙層平板法進行宿主譜分析。取100 μL培養至對數期的108株E.coli待檢菌培養液與100 μL噬菌體或PBS混合后鋪雙層瓊脂平板,PBS作為陰性對照。待平板晾干后,將其置于37 ℃ 溫箱培養5~6 h,記錄結果。
1.2.4 K1莢膜型大腸桿菌的鑒定 對用于宿主譜鑒定的細菌進行K1莢膜的鑒定。以F(上游引物):5′-CATCCAGACGATAAGCATGAGCA-3′,R(下游引物):5′-GCGCATTTGCTGATACTGTTG-3′為鑒定K1莢膜的特異性引物[18],使用PCR方法對宿主譜中的細菌進行K1型莢膜鑒定,擴增產物長度約為270 bp。PCR反應體系包括12.5 μL GreenTaqMix(Vazyme),上、下游引物各1 μL,超純水 8.5 μL,菌液模板2 μL。PCR擴增條件為95 ℃預變性5 min;95 ℃變性30 s,55 ℃退火30 s,72 ℃延伸20 s,30個循環;72 ℃終延伸10 min,4 ℃保存。取7 μL PCR產物進行瓊脂糖凝膠電泳,鑒定目的條帶。
1.2.5 最佳感染復數的檢測 參照Carey-Smith等[19]的方法進行噬菌體的感染復數(multiplicity of infection,MOI)的檢測。將細菌培養至濃度約為3×108CFU·mL-1,用LB液體培養基稀釋10倍。取不同稀釋濃度的噬菌體和濃度為3×107CFU·mL-1的宿主菌各200 μL,按照感染復數分別為0.001、0.01、0.1、1、10和100的比例混合,37 ℃,180 r·min-1震蕩培養5 h,10 000g離心8 min, 收集上清并用0.22 μm濾器過濾。將濾液進行梯度稀釋后,用雙層平板法測定噬菌體效價,效價最高的感染復數即為最佳感染復數(optimal multiplicity of infection)。
1.2.6 一步生長曲線測定 參照Ellis等[20]的方法測定噬菌體PNJ1809-36的一步生長曲線。取噬菌體和細菌各1 mL按照最佳MOI混合,37 ℃靜置10 min,然后10 000g離心10 min,并用2 mL LB液體培養基洗滌沉淀2次,以去除沒有吸附在細菌上的噬菌體,隨后用預熱好的5 mL LB液體培養基重懸沉淀,迅速置于37 ℃搖床震蕩培養。將置于37 ℃搖床震蕩開始的時間記為T0,每隔5 min或10 min取樣1次,分別在T0、T5、T10、T15、T20、T30、T40、T50、T60、T70、T80、T90、T100、T110、T120各取混合液100 μL,用雙層平板法測定噬菌體效價。試驗重復3次。試驗結果以時間為橫坐標,噬菌體的滴度為縱坐標,繪制一步生長曲線。
1.2.7 溫度及酸堿敏感性測定 參照文獻[21]描述的方法進行噬菌體溫度及酸堿敏感性測定。向每個1.5 mL EP管中加入1 mL噬菌體后,將其分別置于40、50、60、70、80 ℃的金屬浴中進行孵育。在30和60 min各取出100 μL噬菌體,稀釋后用雙層平板法檢測效價。每組重復3次。用HCl和NaOH調節SM液的pH至3~12,分別取900 μL不同pH的SM液加入1.5 mL EP管中,再向每管中加入100 μL噬菌體,37 ℃培養1 h,用雙層平板法檢測噬菌體的效價,每組重復3次。
1.2.8 體外裂解曲線測定 取500 μL濃度為3× 108CFU·mL-1的宿主菌用液體LB洗滌3次,用10倍體積的LB重懸。按照最佳MOI將細菌和噬菌體各取100 μL混勻,加入96孔板中,每孔200 μL, 重復5孔。取重懸后的菌液和液體LB各100 μL混勻加入96孔板中,每孔200 μL,重復5孔, 作為菌液對照。將200 μL LB加入96孔板中,重復5孔作為空白對照。將酶標儀參數設置為恒溫37 ℃,180 r·min-1震蕩,持續15 h,每30 min檢測1次OD600nm,用所得數值繪制體外裂解曲線。
1.2.9 全基因組測序及生物信息學分析 使用苯酚-氯仿法[22]提取噬菌體的基因組DNA,用雙蒸水溶解后,-20 ℃保存。將噬菌體DNA送至北京化工大學進行全基因組測序,采用Illumina Hiseq測序平臺,TruSeqTMDNA Sample Prep Kit構建DNA文庫。使用Newbler V3.0軟件(Roche 454)和CLC軟件(CLC Bio)組裝拼接噬菌體的完整基因組序列。基因組序列拼裝結果由北京化工大學提供。利用NCBI在線工具BLASTp(http://blast.ncbi.nlm.nih.gov/)對噬菌體基因組開放閱讀框(ORF)作功能注釋,并利用GC view Sever繪制噬菌體的全基因組圖譜。以噬菌體的RNA連接酶和全基因組為基準,使用MEGA-X繪制噬菌體的系統發育樹,利用Maximum Likelihood最大似然法進行同源進化分析。
2 結 果
2.1 形態學觀察
以K1莢膜大腸桿菌DE058為宿主菌,用雙層平板法進行噬菌體分離。經反復純化后,得到以DE058為宿主菌的噬菌體PNJ1809-36。在雙層瓊脂上產生的噬菌斑大小均一,邊緣整齊,斑清澈透亮(圖1A)。經過負染后,進行電鏡觀察,該噬菌體的頭部直徑為80 nm,為正多面體;具有可以收縮的尾部,長度大約100 nm,底板上有較多的短尾絲(圖1B),表明PNJ1809-36為肌尾科噬菌體。

A. PNJ1809-36形成的噬菌斑形態;B. PNJ1809-36負染后的電鏡圖(比例尺為20 nm)
2.2 宿主譜分析
對實驗室保存的108株大腸桿菌使用雙層平板法測定噬菌體PNJ1809-36的裂解譜。其中21株菌可以被裂解(表1),經PCR鑒定這些菌株均為K1莢膜菌株。圖2列舉了部分PCR鑒定結果,其中泳道1、5、9、19在270 bp處出現條帶,鑒定為K1莢膜型大腸桿菌。該試驗結果表明,該噬菌體對K1型大腸桿菌具有廣譜抗菌活性。
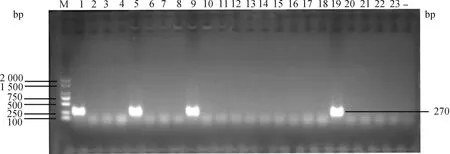
M. DL2000 DNA相對分子質量標準;1~23.檢測K1莢膜的基因條帶;-.陰性對照
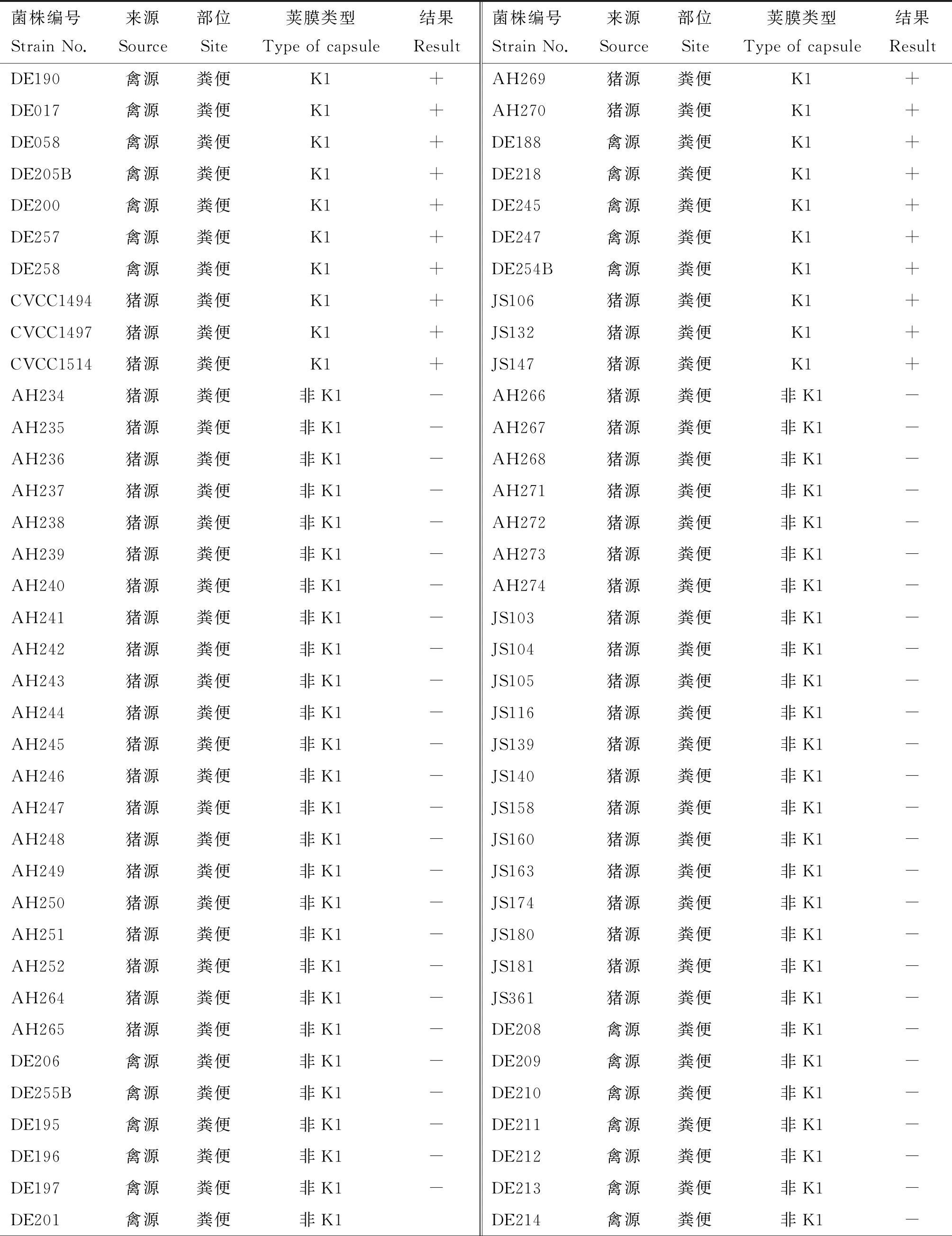
表1 噬菌體PNJ1809-36宿主譜
2.3 一步生長曲線
一步生長曲線測定結果顯示(圖3 A),前10 min噬菌體效價無明顯變化,10~50 min噬菌體的效價急劇升高,然后趨于穩定。即噬菌體PNJ1809-36的潛伏期為10 min,爆發期為40 min。爆發期時噬菌體效價最高可達1.84×108PFU·mL-1。由“爆發量=噬菌體末期數量/初期細菌數量”可知,噬菌體PNJ1809-36的爆發量為122。
2.4 最佳MOI
如圖3 B所示,將噬菌體和細菌按照不同的MOI混合后測定噬菌體的滴度,比較在不同的MOI情況下產生噬菌體的數量。在MOI為0.01時,噬菌體的滴度最高,因此PNJ1809-36與DE058宿主菌的最佳MOI為0.01。
2.5 熱穩定性及酸堿耐受性
通過對噬菌體PNJ1809-36的熱穩定性檢測可知(圖3 C),在40 ℃時,噬菌體的效價比較穩定;50 ℃ 孵育30 min時,噬菌體存活率為94.34%,孵育60 min時,存活率為67.65%;60 ℃時,噬菌體存活率明顯下降,孵育時間為30 min時,存活率僅為0.2%,孵育時間為60 min時,存活率為0;70和80 ℃ 時,孵育30和60 min噬菌體存活率均為0,表明60 ℃以上30 min可使噬菌體幾乎完全滅活。
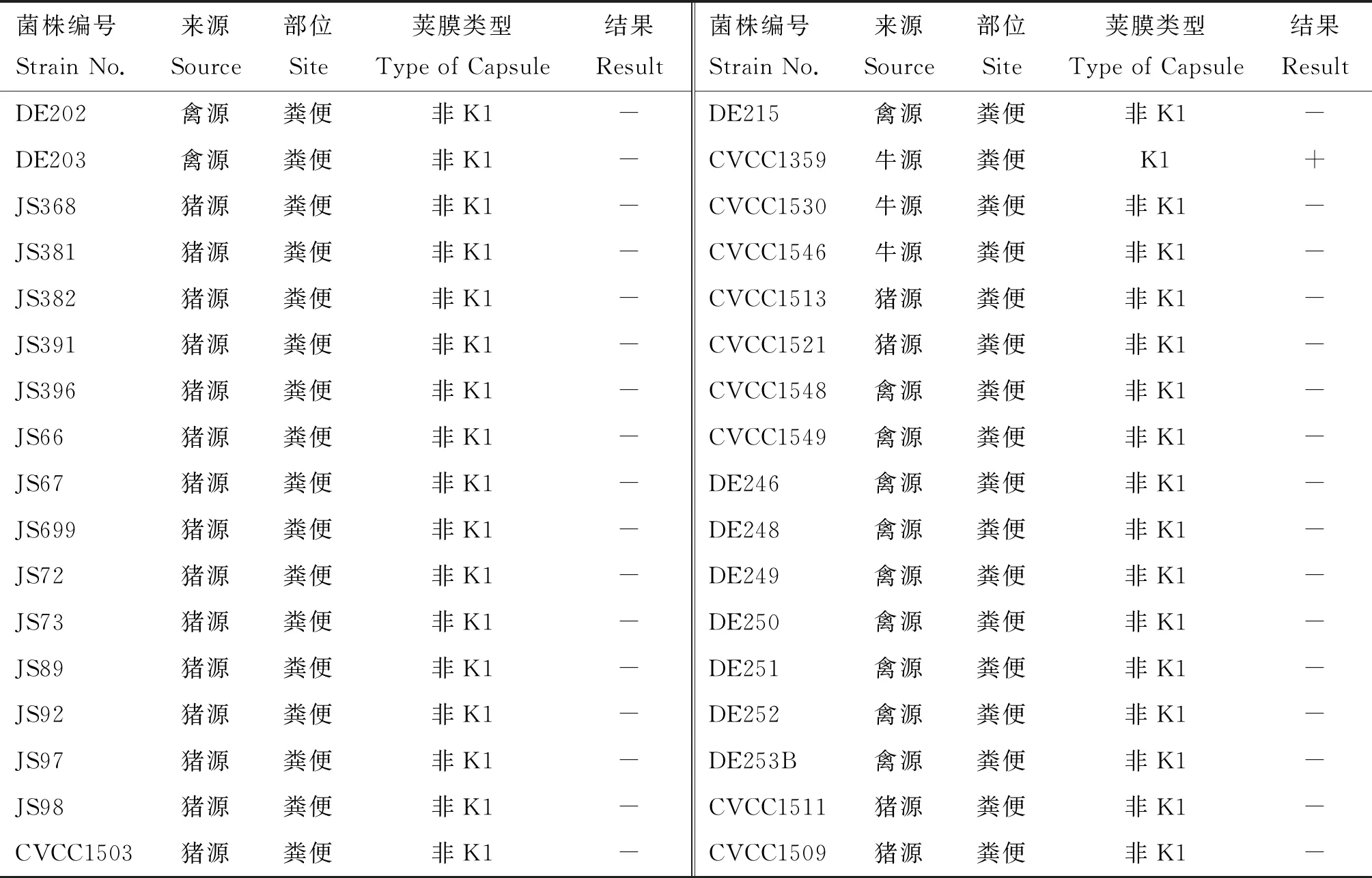
(續表1 Continued)
圖3D表示噬菌體PNJ1809-36對于不同酸堿度的耐受情況。在pH為3~11,PNJ1809-36均可存活;pH為6時,噬菌體的活性最高;pH為2或12時, PNJ1809-36完全失活。
2.6 體外裂解曲線
將噬菌體PNJ1809-36和宿主菌以最佳MOI混合培養,持續觀察15 h,結果如圖3 E所示,在前6 h內,噬菌體PNJ1809-36可完全抑制宿主菌DE058的生長;而未加噬菌體的宿主菌則保持持續增長,大約在12 h后進入平臺期;空白對照則未見細菌生長,表明該體系沒有被污染。
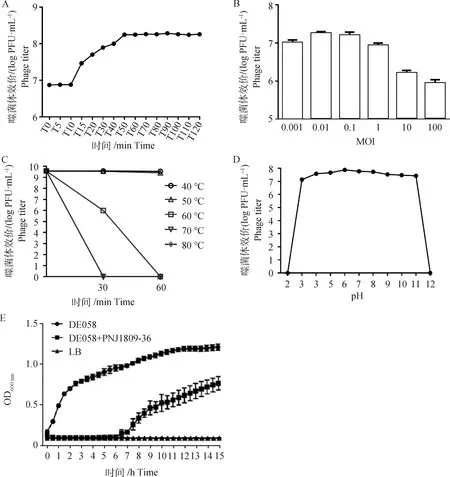
A.噬菌體PNJ1809-36一步生長曲線;B.最佳 MOI檢測;C.溫度敏感性檢測;D.酸堿度耐受性檢測;E.噬菌體體外裂解曲線
2.7 全基因組測序及分析
噬菌體PNJ1809-36基因組為線性的雙鏈DNA,已將該基因組上傳至GenBank(Accession number:MT944117)。經分析可知,其基因組長度為152 343 bp;GC含量為39.11%;A、T、G、C含量分別為30.76%、30.13%、19.27%、19.83%。根據基因功能預測,該基因組共有277個開放閱讀(ORF),11個tRNA基因。277個ORF中,僅有36個注釋了可能存在的功能(表2),包括與噬菌體的結構和包裝相關的基因、DNA復制過程中的相關酶、裂解宿主相關基因以及噬菌體的尾部纖維基因,無噬菌體整合酶基因。剩余的87% ORF均為未知功能的蛋白。
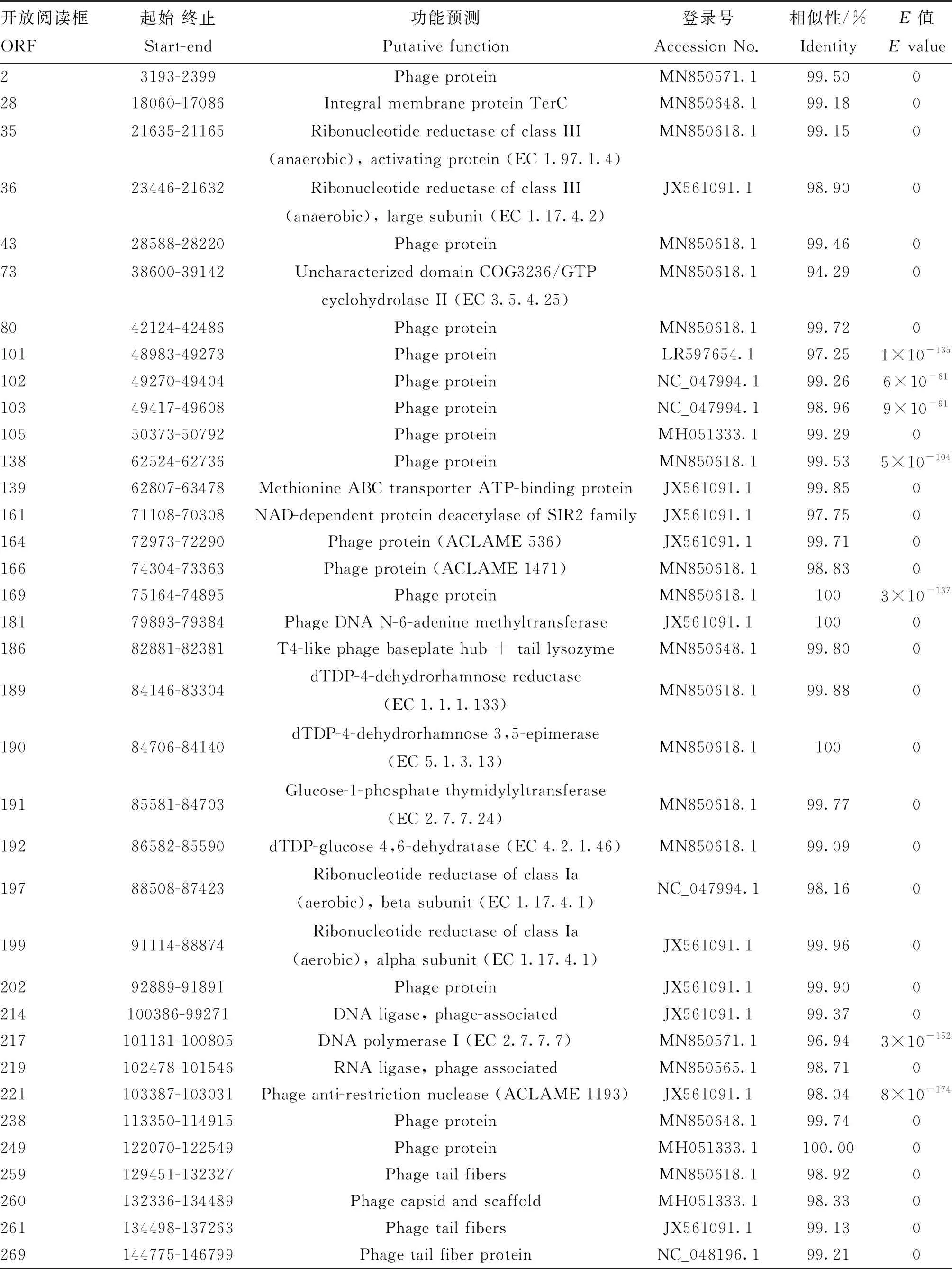
表2 噬菌體PNJ1809-36部分ORFs預測
使用GC view Server[23]繪制了全基因組圖譜(圖4),并用不同的顏色進行標注。粉色代表假定蛋白,藍色為噬菌體結構蛋白,黃色標注基因與噬菌體復制和包裝有關,紅色為尾部纖維蛋白。由于該噬菌體的大多數基因與其他噬菌體的同源性較低,因此存在大量的未知功能的假定蛋白,可見該噬菌體基因異質性較強,有待繼續研究和驗證。
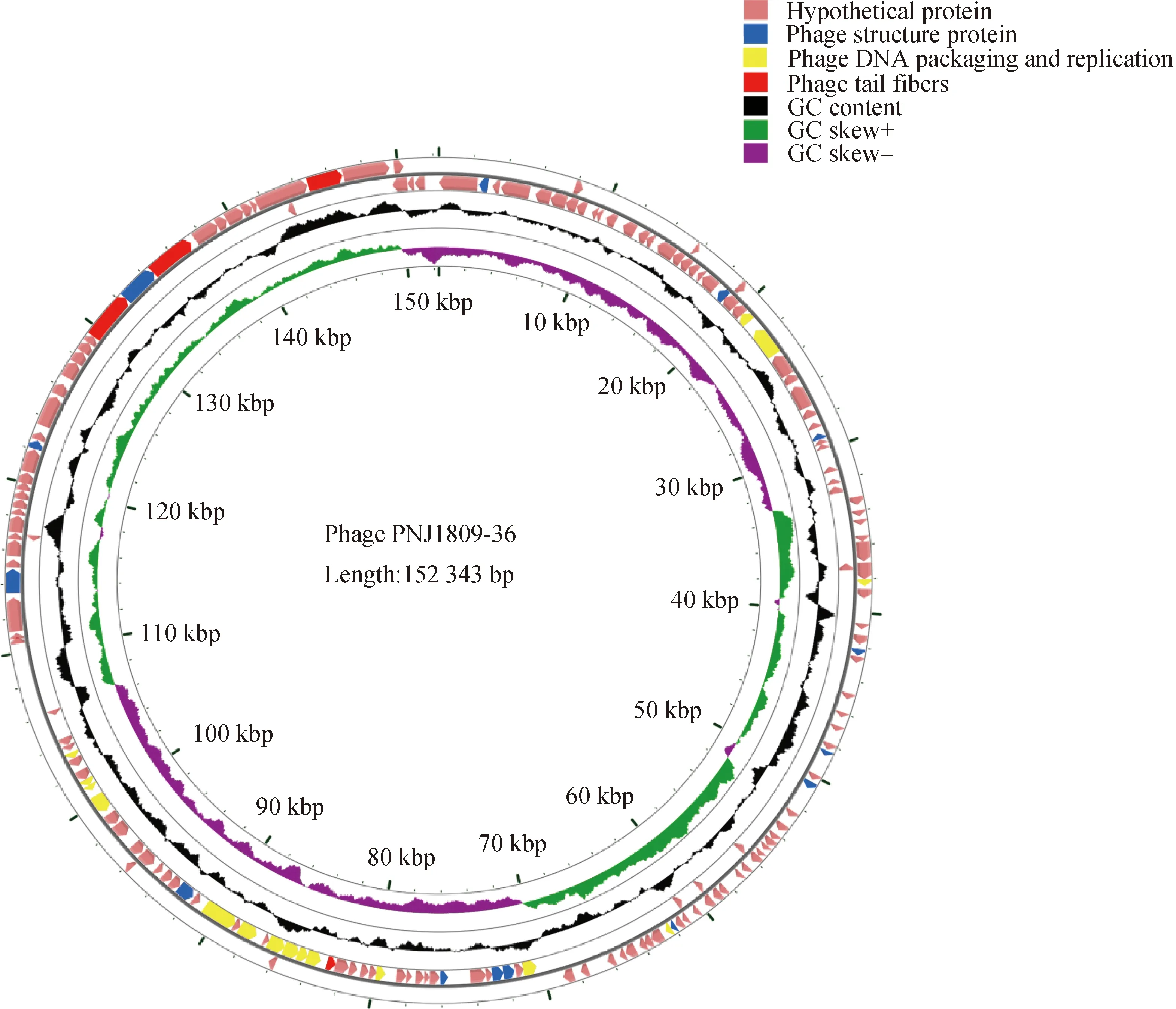
粉色為假定蛋白;藍色為噬菌體結構蛋白;黃色為DNA復制和外殼包裝有關蛋白;紅色為噬菌體尾部蛋白;黑色代表GC含量;綠色和紫色代表正、負平均GC偏差
2.8 系統發育樹分析
為了解噬菌體PNJ1809-36與其他噬菌體之間的進化關系,選擇相對保守的且具有進化意義的基因進行系統發育樹分析,例如頭部蛋白、末端酶大亞基[24]、RNA聚合酶[25]、RNA連接酶[26]以及全基因組等。本研究以RNA連接酶為基準,采用MEGA-X軟件中的Maximum Likelihood繪制同源進化樹(圖5)。分析結果顯示,噬菌體PNJ1809-36與噬菌體vB EcoM-Ro121c4YLVW(2019年分離)、噬菌體nepoznato(2019年分離)親緣性較近,這兩株噬菌體基因組中均含有與K1莢膜降解相關的蛋白,Korf等[27]建議將這類噬菌體歸于肌尾科噬菌體中的新屬,并將該新屬命名為“Phapecoctavirus”。

分支點上的數字. 可信度,數值越接近100,可信度越強;標尺.代表遺傳距離;黑色三角.PNJ1809-36
3 討 論
隨著細菌耐藥現象的持續惡化,尋找新的抗生素替代品乃當務之急。而噬菌體作為一種細菌病毒,具有特異性分解某些細菌的能力,在克服耐藥領域具備一定的潛力。本研究分離得到的大腸桿菌噬菌體PNJ1809-36是一株肌尾科噬菌體,透射電鏡下可以發現其頭部大小約為80 nm且呈正多面體,具有可以收縮的尾部,大約100 nm。它可以在雙層瓊脂上形成大小均一、透亮的噬菌斑。一步生長曲線顯示該噬菌體PNJ1809-36的潛伏期為10 min,爆發期持續40 min,裂解量為122,說明其吸附宿主菌所需時間較短,復制子代效率較高。最佳MOI為0.01,表明噬菌體在裂解細菌時所需數量較少,在實際應用中較低的MOI會降低純化及應用成本。其溫度和酸堿耐受性顯示,在40 ℃以下活性完全不會被抑制,60 ℃以上被滅活。pH為6的弱酸性條件下活性最好。這提示該噬菌體可能適用于體內細菌感染的治療。體外裂解曲線顯示了在最佳MOI的比例下,噬菌體可以在6 h內持續抑制細菌的生長,給使用該噬菌體進行治療時的給藥時間提供了參考。在噬菌體的基因組分析中沒有發現噬菌體整合酶基因,結合其培養特征,可以證明該噬菌體為烈性噬菌體。
PNJ1809-36對20株K1莢膜大腸桿菌均有裂解作用,這提示噬菌體PNJ1809-36可能是一個K1莢膜特異性的噬菌體。具有K1莢膜的大腸桿菌能夠引起新生兒腦膜炎,尤其是近幾年隨著耐藥性K1菌株的出現,情況變得嚴峻。針對K1莢膜菌株的噬菌體能夠降解K1莢膜,使細菌毒力下降,在治療K1菌株引起的疾病上具有潛力。
噬菌體PNJ3809-36與nepoznato親緣性非常高,提示PNJ1809-36可能和近期發現的噬菌體同屬于能夠分解K1莢膜且具有可收縮尾部的肌尾科噬菌體。且Lavigne等[28]提到,這些噬菌體應當屬于新的肌尾科系統發育分支。莢膜特異性噬菌體是一種特殊的能夠降解胞外多糖的噬菌體。根據莢膜類型的不同,對應的噬菌體也進行了分類[15]。研究最多的是針對K1莢膜的噬菌體。K1莢膜是一種單一的多聚唾液酸[29]。在早期發現的針對K1莢膜的噬菌體多為短尾噬菌體:如K1E[30]、K1F[31]等。后來發現了長尾噬菌體63D[32]也能夠特異性分解K1莢膜。近十年又發現了與早期發現的這些噬菌體同源性較低的新的降解K1莢膜的噬菌體,如沙門菌噬菌體 PVP-SE1[33](2011年分離)、大腸桿菌噬菌體phAPEC8[34](2010年分離)、ESCO5、ESCO13[35](2017年分離)、nepoznato(2019年分離)等。
噬菌體宿主特異性主要取決于其尾部蛋白與細菌表面受體的結合,宿主菌的外膜蛋白、莢膜、菌毛或鞭毛上都存在特異性的受體,這些受體以蛋白質、多糖或脂多糖形式存在。在本研究中,噬菌體PNJ1809-36的受體為K1莢膜,是一種胞外多糖,其成分為多聚唾液酸[36]。據報道裂解K1莢膜型大腸桿菌的噬菌體存在一種尾部蛋白,即內切唾液酸酶,能夠將多聚唾液酸分解成2,8唾液酸單體,從而降解K1莢膜[37]。通過基因比對發現,PNJ1809-36基因組中ORF261與K1莢膜特異性噬菌體的內切唾液酸酶的氨基酸相似性為74%[34],由此可推測ORF261可能具有內切唾液酸酶的功能,后續我們會進一步驗證該酶的功能。
4 結 論
噬菌體PNJ1809-36是1株特異性高,裂解能力強的噬菌體。它具有能夠針對性裂解K1莢膜大腸桿菌的特性,提示該噬菌體具有防控K1莢膜大腸桿菌感染的潛力。
