豬源組織蛋白酶S抑制O型口蹄疫病毒在PK-15細胞復制
2021-06-30 13:57:12史喜絹劉原子張大俊申超超袁興國任瑞瑞杜曉華張克山鄭海學劉湘濤
畜牧獸醫學報 2021年6期
關鍵詞:檢測
史喜絹,劉原子,張大俊,侯 景,申超超,楊 博,張 婷,袁興國,任瑞瑞,杜曉華*,張克山*,鄭海學,劉湘濤
(1.甘肅農業大學動物醫學院,蘭州 730070; 2.中國農業科學院蘭州獸醫研究所 家畜疫病病原生物學國家重點實驗室 農業部畜禽病毒學重點開放實驗室 國家口蹄疫參考實驗室,蘭州 730046)
口蹄疫(foot-and-mouth disease,FMD)是由口蹄疫病毒(foot-and-mouth disease virus,FMDV)感染偶蹄動物引起的一種烈性傳染病[1-3]。FMDV是微RNA病毒科(Picornaviridae)口蹄疫病毒屬(Aphthovirus)的成員,已知有O、A、C、Asia1和SAT1、SAT2、SAT3 7種血清型,成熟的FMDV粒子無囊膜,具有二十面體對稱性[4-6]。FMDV全基因組約為8 400 bp,含有一個大的開放閱讀框(ORF),ORF編碼的多聚蛋白被病毒自身編碼的蛋白酶(L、2A、3C)切割成4個結構蛋白和8個非結構蛋白[7-9]。FMDV可利用自身編碼的蛋白進化形成抑制或逃避宿主先天性免疫反應,從而促進其自身在宿主體內的存活和復制[10]。FMDV前導蛋白酶(LPro)通過降低IFN-α/β和干擾素刺激因子的早期分泌水平,從而阻止宿主蛋白的合成[11]。FMDV 3A蛋白能抑制病毒觸發的IFN-β信號通路,從而逃避宿主免疫反應[12]。同樣,宿主也可以識別病原體并激發炎癥反應來抑制病毒復制[13]。已經發現有一些宿主蛋白在FMDV復制過程中起重要作用,如DCTN3與FMDV 3A結合負調控FMDV復制[14]。
CTSS是半胱氨酸蛋白酶家族中具有內切肽酶活性的重要成員,由N端16 aa信號肽(SP)、前肽和成熟肽331個氨基酸組成的非活化酶原,其活化需要蛋白酶裂解其N端前肽或由各種因素誘導[15]。CTSS酶活性是其發揮功能的關鍵,如棕櫚酸酯抑制組織蛋白酶誘導內皮細胞侵襲從而抗血管生成,部分是通過抑制CTSL和CTSS活性而起作用的[16]。CTSS的N端有3個凹槽(S1、S2 和S3)與底物的特異性結合相關,這一特性也決定了半胱氨酸蛋白酶抑制劑的特異性;該基因C端有一個與底物結合的位點,即S1’,在酶與主要組織相容性復合體二類分子(MHC-Ⅱ)保守區的特異性結合中起關鍵作用[17]。CTSS主要在樹突狀細胞、B細胞和巨噬細胞等抗原呈遞細胞中表達[18],其參與細胞外基質、抗血管生成肽和黏附蛋白的降解,促進新生血管形成和腫瘤細胞侵襲轉移[19]。CTSS通過調節p38 MAPK和JNK1 途徑參與甲基原薯蕷皂苷(methyl protodioscin,MP)誘導的細胞凋亡和自噬[20];通過激活NF-κB和caspase-3 從而誘導肝癌細胞凋亡并增加其化學敏感性[21];也可通過激活CD74調控趨化因子CCL2的表達,進而對腫瘤微環境產生影響[22]。目前對CTSS的研究主要集中在自身免疫性疾病[23]、心血管疾病[24]及腫瘤相關疾病[25],目前尚無有關病原體方面的報道。
初乳在抵抗病原感染中具有重要作用[26]。本團隊前期應用iTRAQ技術研究發現母豬初乳中CTSS的含量顯著高于常乳,但目前對宿主CTSS在病原感染中的作用研究較少,宿主CTSS在FMDV感染中的作用及其調控機制至今尚不明確。為闡明宿主CTSS在FMDV-O感染過程中發揮的作用,本研究探究了FMDV-O感染和宿主CTSS的相互調控作用,發現宿主CTSS能夠抑制FMDV-O在PK-15細胞中復制,而FMDV-O感染增加了宿主CTSS酶活性,進一步研究發現CTSS促進FMDV-O誘導的抗病毒細胞因子產生,明確了豬源CTSS抑制FMDV-O復制的初步原因。本研究結果為更深層次探究豬源CTSS在FMDV-O觸發的免疫應答中的作用機制奠定了基礎。
1 材料與方法
1.1 材料
口蹄疫病毒毒株FMDV O/MYA98/BY/2010、PK-15細胞和FMDV-O抗體由蘭州獸醫研究所口蹄疫與新發病流行病學團隊保存;兔多克隆抗體CTSS購于Abcam公司;鼠抗Flag單抗、鼠抗Myc單抗、鼠抗β-actin單抗、HRP標記山羊抗鼠IgG二抗和HRP標記山羊抗兔IgG二抗均購于Thermo Scientific公司。
大腸桿菌DH5α感受態、LATaqDNA聚合酶、限制性核酸內切酶BamHⅠ和XhoⅠ、T4 DNA連接酶、RNA抽提試劑Trizol、5×Prime script RT Master Mix、SYBR Permix ExTaqII和蛋白預染Marker均購于寶生物工程大連有限公司; LipofectamineTM2000轉染試劑購于Invitrogen公司;Opti-MEM、0.25% EDTA胰酶和胎牛血清(FBS)均購于Gibco公司;MEM細胞培養液和PBS溶液購于建順公司;ECL顯色劑購于Thermo Scientific公司;NP-40裂解液和PMSF購于碧云天公司;織蛋白酶S試劑盒(貨號ab65306)購于艾博抗(上海)貿易有限公司;CTSS干擾序列由上海吉瑪制藥有限公司合成。
1.2 方法
1.2.1 CTSS真核表達質粒的構建 根據GenBank公布的CTSS基因序列(XM_021089893.1)設計合成CTSS引物,引入酶切位點BamHⅠ和XhoⅠ,以pcDNA3.1為載體,構建pcDNA3.1-CTSS-Myc真核表達質粒,進行PCR擴增、酶切和序列測定。
1.2.2CTSS基因 RNAi序列設計與合成 根據GenBank公布的CTSS基因序列(XM_021089893.1)設計并合成CTSS RNAi序列。分別設計了3對針對CTSS基因的RNAi序列。
1.2.3 細胞瞬時轉染和病毒感染 將細胞消化后接種于細胞板中,待細胞長至70%~90%時,將質粒與Lip2000試劑(DNA∶Lip2000=1 μg∶2 μL) 分別加至Opti-MEM中,混合后靜置15 min,將Opti-MEM混合物直接加至細胞中,將細胞放于培養箱中培養數小時。用無血清的MEM清洗細胞,用無血清的MEM將FMDV-O稀釋至MOI為1.0時感染PK-15細胞,置于37 ℃、5%CO2培養箱孵育1 h之后,棄去病毒液,用含2% FBS的MEM維持液繼續培養。在感染后12 h收取2份細胞樣品,一份用于RT-qPCR,分別檢測CTSS和FMDV-O轉錄水平的變化,并以豬源GAPDH作為內參;一份用于Western blot,分別檢測CTSS和FMDV-O蛋白水平的變化,并以β-actin作為內參。
1.2.4 RT-qPCR 收集細胞樣品,采用Triozl法提取細胞總RNA,利用合成好的引物進行絕對定量和相對定量檢測[1]。相關定量擴增引物信息見表2。

表2 引物序列信息
1.2.5 CTSS酶活性測定 利用組織蛋白酶S活性檢測試劑盒(貨號ab65306;Abcam)提供的裂解液裂解細胞,離心取50 μL上清于96孔板,加等量反應緩沖液和10 mmol·L-1Ac-VVR-AFC(CTSS底物),根據試劑盒說明書進行處理。使用SpectraMax M5熒光計在400 nm激發波長和505 nm發射波長下測量熒光。
1.2.6 Western blot 收樣并處理細胞樣品,加入適量的NP-40裂解液(PMSF 1 mol·L-1);充分裂解后,12 000 r·min-1離心10 min,取上清加入含β-巰基乙醇的5×SDS Loading Buffer,100 ℃變性10~15 min,按20 μL的上樣量進行SDS-PAGE凝膠電泳[1],最后用高分辨圖像采集系統進行ECL顯影并保存結果。
1.2.7 數據分析 所有試驗至少重復3次,應用 GraphPad Prism 7軟件進行分析并作圖,運用獨立樣品T檢驗進行統計學分析,*.P<0.05表示數據具有統計學意義,**.P<0.01表示數據具有顯著性差異,***.P<0.001表示數據間具有極顯著性差異。
2 結 果
2.1 FMDV-O感染PK-15細胞促進內源性CTSS表達
將PK-15細胞鋪于35 mm小皿中,待細胞長至80%~90%,用MOI為1的FMDV-O感染PK-15細胞,在0、4、8、12 h后收取細胞樣品,利用RT-qPCR和Western blot方法檢測內源性CTSS的變化。結果表明FMDV-O感染PK-15細胞后內源性的CTSS蛋白水平(圖1A)和轉錄水平(圖1B)均高于對照組,提示FMDV-O感染可促進宿主細胞內源性CTSS的表達。
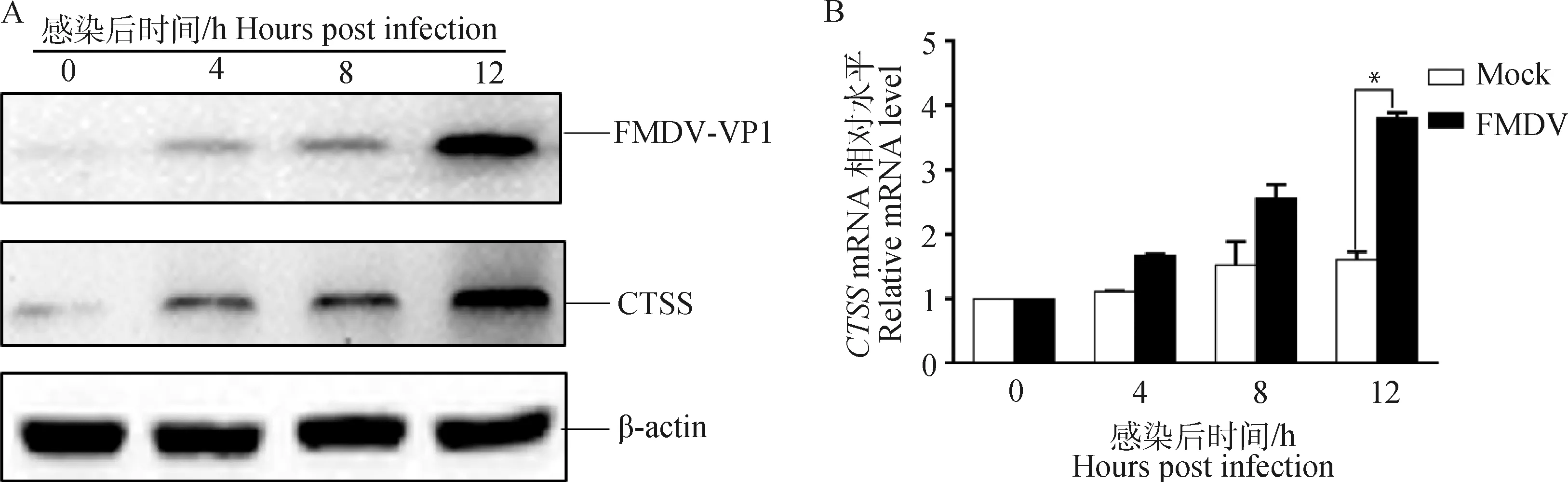
A. Western blot檢測CTSS蛋白水平的變化;B. RT-qPCR檢測CTSS轉錄水平的變化;*.P<0.05
2.2 CTSS重組質粒構建及表達驗證
構建pcDNA3.1-CTSS-Myc重組質粒,進行PCR擴增,用BamH Ⅰ和XhoⅠ雙酶切鑒定,10 g·L-1瓊脂糖凝膠電泳檢測可在1 000 bp處見目的條帶,在5 000 bp 見載體條帶(圖2A)。經測序后進一步確定該真核質粒構建成功。將構建的pcDNA3.1-CTSS-Myc以不同的劑量分別瞬時轉染PK-15細胞,24 h后收取細胞樣品,處理樣品并進行Western blot驗證其表達情況,結果表明pcDNA3.1-CTSS-Myc重組質粒在PK-15細胞中呈劑量依賴性表達(圖2B)。
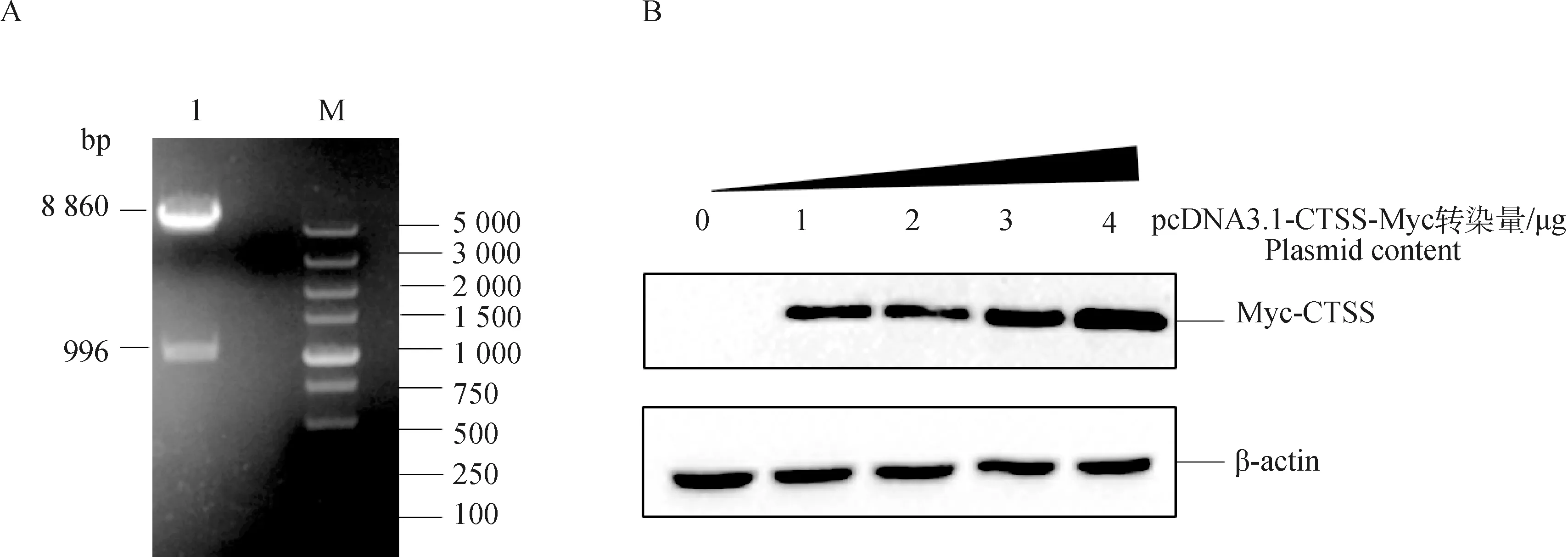
A. 雙酶切鑒定結果(M. DL5000 DNA相對分子質量標準;1. BamHⅠ和XhoⅠ酶切產物);B. Western blot驗證CTSS蛋白在PK-15細胞表達
2.3 FMDV-O感染能上調CTSS酶活性
轉染0.25 μg CTSS重組質粒至PK-15細胞,20 h后用FMDV-O(MOI=1)感染細胞,同時設不用病毒刺激的Mock組,收取0、2、4、6、8、10、12、14 h 細胞樣品,裂解細胞用Fluorometric Method檢測CTSS酶活性。結果表明,FMDV-O感染能上調CTSS酶活性,且隨著FMDV-O感染時間的增加,CTSS的活性也隨之增加(圖3)。
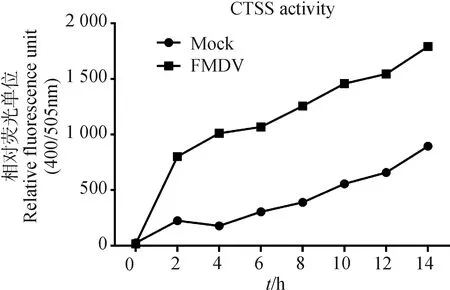
圖3 FMDV-O感染上調CTSS酶活性
2.4 過表達CTSS能抑制FMDV-O復制
轉染1、2、4 μg pcDNA3.1-CTSS-Myc至PK-15細胞,轉染24 h后用MOI為1的FMDV-O感染細胞,12 h后收取細胞樣品,檢測其對FMDV-O復制的調控作用,發現隨著CTSS表達量增加(圖4A、C),FMDV-O復制水平呈現劑量依賴性降低(圖4B、C)。結果表明,過表達CTSS抑制FMDV-O在PK-15細胞中復制。
A. RT-qPCR檢測CTSS轉錄水平的變化;B. RT-qPCR檢測FMDV-O拷貝數的變化;C. Western blot檢測CTSS和FMDV-O蛋白水平的變化; *.P<0.05; **.P<0.01; ***.P<0.001
2.5 CTSS siRNA干擾序列的篩選及其對FMDV-O復制的促進作用
為進一步確定宿主CTSS對FMDV-O復制的影響,針對CTSS設計合成3對特異性siRNA(表1),將siRNA-2947、siRNA-3629、siRNA-3458分別轉染至PK-15細胞, 在24和36 h分別收取樣品,用RT-qPCR方法選擇干擾效果最好的序列,結果顯示編號siRNA-2947的干擾效果最好(圖5A)。在此試驗結果基礎上,PK-15細胞轉染siRNA-2947,以NC siRNA為對照,36 h后用等量FMDV-O(MOI=1)感染細胞,12 h后收取細胞樣品,用RT-qPCR和Western blot檢測siRNA-2947對FMDV復制的影響,結果表明siRNA-2947能下調宿主CTSS的表達進而促進FMDV-O在PK-15細胞中復制(圖5B、C)。

表1 干擾序列信息
A. RT-qPCR檢測CTSS轉錄水平的變化;B. RT-qPCR檢測FMDV-O拷貝數的變化;C. Western blot檢測CTSS和FMDV-O蛋白水平的變化; *.P<0.05; **.P<0.01; ***.P<0.001
2.6 過表達CTSS促進FMDV-O誘導的宿主抗病毒細胞因子產生
為明確豬源CTSS抑制FMDV-O復制的原因,檢測宿主CTSS是否影響由FMDV-O感染誘導的抗病毒細胞因子產生。在PK-15細胞中分別轉染CTSS和pcDNA3.1,24 h后感染FMDV-O(MOI=1),并設無FMDV-O感染對照組,12 h后收集細胞處理樣品。RT-qPCR結果顯示,CTSS可促進FMDV誘導的IFN-α、IFN-β、IFN-γ、IL-6、IL-10、IL-4的mRNA水平,說明宿主CTSS能激活FMDV-O誘導的宿主抗病毒細胞因子產生。
*.P<0.05; **.P<0.01; ***.P<0.001
3 討 論
CTSS是一種溶酶體蛋白酶,主要在抗原呈遞細胞中表達,其活性調節對于MHC-Ⅱ信號傳導及CD4+T細胞介導的免疫反應激活非常重要[27]。有研究報道CTSS活性可以由腸道菌群調節,共生體觸發生理性CTSS活性;病原體引起病理性CTSS活性增加,導致T細胞活化和增殖[28]。而本研究發現FMDV-O感染PK-15細胞可上調內源性CTSS的表達并增強CTSS活性。
FMDV以其自身優勢拮抗宿主免疫應答進而達到成功感染宿主的目的,當然其生命周期也受不同宿主因素影響[29-32]。有文獻報道熱休克蛋白DNAJA3與VP1互作并通過自噬/溶酶體途徑降解VP1,從而減弱VP1對IFN-β信號通路的拮抗作用,最終抑制FMDV復制[30]。本研究發現FMDV-O感染PK-15細胞可上調內源性CTSS的表達并增強CTSS活性,過表達CTSS抑制FMDV-O在PK-15細胞中復制,而下調內源性CTSS能促進FMDV-O復制。有報道稱CTSS能使CX3CL1與CX3CR1相互作用,將免疫細胞募集到炎癥部位增加CX3CL1脫落進入間質,從而改變自身免疫性淚腺炎和淚腺分泌[33];干燥綜合征患者淚液中CTSS活性升高可誘導促炎細胞因子產生[34];缺乏CTSS會增加高血壓小鼠線粒體的受損并提高ROS水平和NF-κB活性,從而調節心臟炎癥和纖維化[35]。筆者隨后檢測了CTSS對抗病毒細胞因子mRNA水平的影響,RT-qPCR結果顯示CTSS可促進FMDV-O誘導的IL-6、IL-10、IL-4、IFN-α、IFN-β、IFN-γ基因的轉錄。這一結果與文獻報道一致,提示CTSS可能參與炎癥反應。天然殺傷細胞(NKT)可募集并激活其他先天免疫細胞,從而調節多種免疫反應,以加劇肝的炎癥反應,但CTSB和CTSS抑制劑可降低LPS誘導的炎癥過程中NKT細胞的活化[36],進一步說明CTSS可能與炎癥有關。FMDV在進化過程中獲得了許多逃避宿主免疫系統的策略[37], 例如FMDV 3A通過破壞RIG-Ⅰ、MDA5和VISA蛋白的表達而抑制病毒觸發的IFN-β信號通路[12];FMDV VP3降解JAK1以抑制IFN-γ信號轉導途徑[38]; LPro可以抑制天然免疫下游抗病毒細胞因子的產生從而促進病毒的復制[39-41],本文雖然檢測了抗病毒細胞因子的變化,但機體的免疫系統是一個錯綜復雜的網絡,宿主除了通過調控干擾素信號通路發揮抗病毒功能;還可以通過自噬和凋亡途徑影響病毒復制,比如PCBP2和FMDV VP0互作可以通過凋亡途徑促進FMDV復制[42]。這說明CTSS促進FMDV-O誘導的抗病毒細胞因子的產生,可能是宿主CTSS抑制FMDV-O復制的原因之一,具體詳細機制還需要進一步研究。
本研究首次證實了CTSS在FMDV-O感染過程中發揮抗病毒作用的新功能,為宿主 CTSS拮抗FMDV-O感染方面的研究提供了理論依據,也為下一步探究豬源CTSS在FMDV-O觸發的免疫應答中的作用積累了素材;此外,本研究結果也提示CTSS可能作為抑制FMDV-O復制的潛在靶點發揮作用。
4 結 論
FMDV-O感染與宿主CTSS之間具有相互調控作用,FMDV-O感染PK-15細胞顯著上調內源CTSS表達并增強CTSS酶活性;過表達CTSS能抑制FMDV-O在PK-15細胞中復制,利用特異性siRNA干擾CTSS表達可以促進FMDV-O復制,并且 CTSS能促進FMDV-O誘導的IFN-α、IFN-β、IFN-γ、IL-6、IL-10和IL-4細胞因子上調表達,明確了CTSS抑制FMDV-O復制的初步原因,具體機制將是下一步研究的方向和重點。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
