暴馬桑黃gpd啟動子的克隆與序列分析
2021-06-21 08:18:56王淑婷唐玉倩劉增才鄒莉
中國林副特產 2021年3期
關鍵詞:分析
王淑婷,唐玉倩,劉增才,鄒莉
(東北林業大學林學院,哈爾濱 150040)
暴馬桑黃(Sanghuangporusbaumii)是寄生在暴馬丁香樹上的一種藥用價值較高的大型真菌,具有抗菌消炎[1]、降血糖[2]、保護肝臟[3-4]、抗癌[5]等功效。其良好的藥用效果主要源于其含有的多糖、黃酮、酚類、三萜等活性成分[6-7]。但是這些成分在暴馬桑黃中含量很低[8],難以滿足生產需求。近年來,隨著分子生物技術的發展,利用基因工程定向培育優質的暴馬桑黃菌株成為一種可靠的技術手段,目前轉基因技術已在多種真菌中成功實現,1986年MUNOZ-RIVAS等人在色氨酸營養缺陷型裂褶菌突變株中轉入TRPC基因,成功獲得正常菌株[9];李剛等人在通過PEG轉換緩沖液將大腸桿菌潮霉素B轉磷酸酶基因轉化到靈芝中,成功轉化出能穩定表達出HmB抗性的轉基因靈芝品種[10]。但目前食藥用菌遺傳轉化依舊存在轉化表達效率低的問題[11],啟動子是控制基因表達的重要工具,因此找到合適的啟動子是提高食藥用菌轉化表達效率的關鍵。
食藥用菌遺傳轉化所采用的啟動子主要有兩種:一種是ras啟動子,另一種則是gpd啟動子[12]。Hirano等在1999年從香菇中分離出gpd啟動子,將其用于香菇轉化體系的構建,結果顯示該體系比ras啟動子轉化體系的表達效率高[13],這說明gpd啟動子比ras啟動子具有更強的調控外源基因表達的能力。此外,gpd啟動子已被用于靈芝[14]、雙孢蘑菇[15]等多種食用菌轉化表達載體的構建,表明運用gpd啟動子進行食用菌轉化有很高的可行性。本試驗通過對暴馬桑黃中分離出的gpd基因啟動子序列進行克隆及序列分析,為建立高效穩定的暴馬桑黃外源基因表達體系奠定基礎,從而獲得優質的目標菌株。
1 材料與方法
1.1 材料
1.1.1 試驗菌株。本研究采用的菌種經ITS鑒定為暴馬桑黃(GenBank登錄號為KP974834),保藏于東北林業大學森林保護學科實驗室。
1.1.2 試驗試劑。DL Marker 2000、DL Marker 10000、6×Loding Buffer、Premix Taq酶、pMD18-T vector均購自大連Takara公司。Tryptone和Yeast Extract購自Oxoid公司。氨芐青霉素購自納川生物技術公司。基因組DNA提取試劑盒和瓊脂糖凝膠DNA回收試劑盒均購自北京天根生化公司。其余藥品均為國產分析純。
1.2 方法
1.2.1 暴馬桑黃菌絲培養與收集。將4 ℃保存的暴馬桑黃菌種取出,在超凈工作臺中用滅過菌的接種鉤挑取黃豆粒大小的菌塊接種在PDA培養基中央,在25 ℃恒溫培養箱中避光培養10 d左右,待菌絲長滿平板后,將表面菌絲體收集到1.5 mL離心管中,-80 ℃保存備用。
1.2.2gpd啟動子引物設計與合成。根據暴馬桑黃基因組數據,通過BioEdit軟件篩選得到暴馬桑黃gpd啟動子的電子序列,利用Primer Premier 5.0軟件設計特異性引物,將其分別命名為gpd-F(5’-AGGTATGACAGTGGAACCAAGCA-3’)和gpd-R(5’-GCATTTCGGAGGACGATACGGC-3’),在哈爾濱擎科生物公司進行引物合成。
1.2.3gpd啟動子序列擴增及測序。利用植物基因組DNA提取試劑盒提取暴馬桑黃總DNA,以得到的DNA為模板,gpd-F和gpd-R為引物,對暴馬桑黃gpd啟動子序列進行PCR擴增。PCR擴增產物用瓊脂凝膠電泳檢測,純化回收后檢測回收產物濃度。將純化回收的gpd啟動子序列與PMD18-T載體相連接并轉化進入DH5α感受態細胞,克隆、篩選和檢驗后送至哈爾濱擎科生物公司測序,并將序列提交到GenBank。
1.2.4gpd啟動子序列分析。利用Neural Network Promoter Prediction(http://fruitfly.org/seq_tools/promoter.html)在線網頁對暴馬桑黃gpd啟動子的核心啟動子區進行分析,利用Plantcare(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)以及Gene Promoter Miner(http://gpminer.mbc.nctu.edu.tw/index.php)對暴馬桑黃gpd啟動子序列的作用元件、CpG島及轉錄因子結合位點分析[16]。
1.2.5gpd啟動子系統發育分析。將暴馬桑黃gpd啟動子序列測序結果提交至NCBI進行BLAST同源性分析,采用MEGA4軟件構建暴馬桑黃與近緣真菌gpd啟動子序列的系統發育樹。
2 結果與分析
2.1 gpd啟動子的PCR擴增結果及膠回收質量檢測

M.Marker 2000;1. PCR擴增產物;2.膠回收產物
以暴馬桑黃總DNA為模板,gpd-F和gpd-R為引物進行PCR擴增,擴增產物采用1%瓊脂凝膠電泳進行檢測,結果在大約1400 bp處顯示出特異性條帶;膠回收DNA凝膠電泳檢測得到質量較好的單一條帶(圖1)。
2.2 gpd啟動子的連接、轉化
將回收的DNA溶液與pMD18-T vector過夜連接,然后轉入大腸桿菌DH5α感受態細胞進行培養,37 ℃培養13 h后,隨機挑選6個單菌落進行搖菌擴繁,然后進行菌液PCR擴增檢測。檢測結果顯示擴增出的條帶均呈陽性,大約1500 bp(包含約100 bp的載體序列),表明目的片段轉化成功(圖2)。陽性的克隆產物在哈爾濱擎科生物公司測序后提交至GenBank (登錄號為MT779798)。

M.Marker 10000;1-6.菌液PCR擴增產物
2.3 gpd啟動子的序列分析
2.3.1 gpd啟動子核心啟動子區分析
核心啟動子區是轉錄因子與RNA聚合酶在啟動子上的結合區域,對啟動子的活性有較大影響[17]。利用Neural Network Promoter Prediction對暴馬桑黃gpd啟動子的核心啟動子區進行分析,結果預測到3個核心啟動子區(表1),其中在995-1045 bp之間為核心啟動子區的概率最高,為0.99,其余兩處均低于0.90。

表1 暴馬桑黃gpd啟動子核心啟動子區預測
2.3.2gpd啟動子序列作用元件分析。利用plantcare對gpd啟動子作用元件分析。結果發現暴馬桑黃gpd啟動子除了含有CAAT-box、TATA-box等典型的啟動子順式作用元件之外,還含有CCAAT-box、G-box、A-box、LTR、ABRE、STRE、MYB等多種重要的作用元件,部分重要元件在gpd序列中的位置及有關功能見表2及圖3,多種暴馬桑黃gpd啟動子作用元件的存在表明克隆得到的目的基因序列具有調控基因表達的功能。

表2 暴馬桑黃gpd啟動子的作用元件種類與功能
2.3.3 CpG島預測及分析。CPG島常位于真核生物編碼基因的調控區,其中啟動子區中處于非甲基化狀態的CpG島是基因轉錄所必需的;轉錄因子結合位點是啟動子上與轉錄因子相結合的DNA片段,是一些起始轉錄的關鍵結合部位。通過Gene Promoter Miner對暴馬桑黃gpd啟動子序列的CpG島及轉錄因子結合位點進行預測分析,結果顯示序列在7-1380 bp之間有一個CpG島(圖3),并且發現啟動子序列中還具有AP1、MAF、OCT1、STAT6、SREBP、IPF1、OSF2等多種轉錄因子的結合位點。

暗色部分代表CpG島
2.3.4 暴馬桑黃與其它真菌gpd啟動子序列系統發育分析。在NCBI網站上對暴馬桑黃gpd啟動子序列進行BLAST比對,獲得近緣真菌gpd序列,采用MEGA4軟件構建暴馬桑黃與其近源真菌gpd啟動子序列系統發育樹(圖4)。發現暴馬桑黃gpd啟動子與香菇(GQ457137.1)及糙皮側耳(HQ286597.1)的gpd啟動子序列同源性相對高,與韋伯靈芝(MN010574.1)和云芝(AY081189.1)等之間的相似性較低,表明gpd啟動子序列在食用真菌種類范圍內保守性不強,也可能因為在暴馬桑黃中還存在其他gpd啟動子[18]。
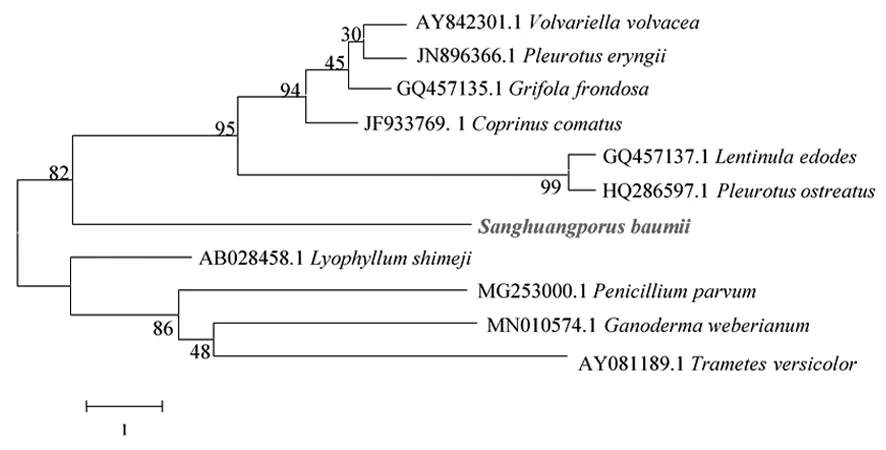
圖4 暴馬桑黃與其它真菌gpd啟動子的系統發育分析
3 討論
gpd啟動子具有的強大調控外源基因表達的能力,使其成為食用菌遺傳轉化體系中重要的調控工具。本研究從暴馬桑黃中克隆得到一段1380 bp的gpd啟動子序列,通過分析發現gpd啟動子在995-1045 bp之間是核心啟動子區概率最高(0.99),說明該部位最有可能是轉錄因子和RNA聚合酶結合的區域,該結果有待于進一步驗證。其含有典型的啟動子順式作用元件CAAT-box、TATA-box等,以及其他重要作用元件如CCAAT-box、G-box、A-box。TATA-box是決定基因轉錄的關鍵作用元件,RNA聚合酶與TATA框牢固結合后才能開始轉錄。在該序列中,共有8個TATA-box,其中4個在正鏈上,這些TATA-box集中分布在gpd基因啟動子的后半段。CAAT-box是啟動子行使正常功能的重要元件,控制轉錄的起始頻率,在暴馬桑黃gpd啟動子中共發現13個CAAT-box,且CAAT-box在轉錄起始點前37-109 bp的區域內分布較集中,與前人研究一致[19],同時也可推測本研究克隆的gpd啟動子具有正常的轉錄功能。除此之外,在該啟動子中還發現了參與光反應的G-box、參與低溫響應的LTR、參與脫落酸反應的ABRE等重要順式元件。
此外,經過預測發現,克隆得到的暴馬桑黃gpd啟動子序列具有多個轉錄因子結合位點,說明克隆得到的啟動子參與多種基因表達的調控。根據是否含有CpG島可將啟動子分為CpG啟動子和非CpG啟動子,啟動子中的CpG島在轉錄過程中有重要作用[20],經預測暴馬桑黃gpd啟動子序列在7-1380 bp之間存在一個CpG島。但是根據已有研究,目前發現的CpG島長度一般在500-1000 bp,此研究預測得到的CpG島卻長達1373 bp,出現該結果的原因尚待進一步研究。
以上序列的分析結果可以初步斷定暴馬桑黃gpd啟動子序列具有啟動子功能,能起到起始轉錄的作用。本研究結果能夠應用于建立高效、穩定的暴馬桑黃遺傳轉化體系,為利用基因工程培育更多目標菌株奠定基礎。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
