黑羽番鴨AMH 基因多態性與產蛋及體重性狀的關聯分析
2021-06-19 02:34:04葛麗巖胡志剛張慧林劉小林
中國畜牧雜志 2021年6期
葛麗巖,胡志剛,張慧林,陳 強,劉小林*
(1.西北農林科技大學動物科技學院,陜西楊凌 712100;2.陜西省潼關安大番鴨育種場,陜西潼關 714300)
近年來,禽類行業發展迅速,肉蛋產品的消費比例逐漸提高[1]。黑羽番鴨的肉蛋產品營養價值高,受到廣大消費者認可[2]。黑羽番鴨原產于南美洲熱帶地區,在中國經過多年馴化和人工選擇,已成為我國優良肉用品種。雄性黑羽番鴨成年體重達3.5~4.5 kg,雌性黑羽番鴨成年體重為2.0~2.7 kg,適合集約化飼養。產肉量和產蛋量決定了黑羽番鴨的經濟價值[3]。目前影響產蛋性能的相關研究大多集中在營養水平、疾病控制以及生長環境[4-6]等方面,少有從遺傳育種角度研究基因多態性對產蛋性能的影響。
在育種工作中,為挖掘有效的遺傳標記常對某基因的單核苷酸多態性(Single Nucleotide Polymorphism,SNP)位點與生產性狀之間進行關聯分析。例如:周璇等[7]發現番鴨神經肽酪氨酸基因(NPY)的AG 基因型產蛋性能顯著高于GG 基因型和AA 基因型;張杉杉[8]發現褪黑素受體1B 基因(MTNR1B)多態性位點(g.9880654 C>T)和神經元特異基因家族成員1 基因(NSG1)多態性位點(g.63058214 C>T)均與番鴨300 日齡產蛋量顯著相關;牟芳[9]在海蘭褐蛋雞中發現α1B-腎上腺素受體基因(ADRA1B)和過氧化物酶體增值物激活受體γ輔激活因子1-β基因(PPARGC1B)多態性位點可作為產蛋性狀的潛在分子標記。
抗繆勒氏管激素(Anti-Mullerian Hormone,AMH)在哺乳動物繁殖方向研究較多,但在家禽繁殖方向研究甚少。AMH 是TGFβ(Transforming growth factor beta superfamily,TGFβ)超家族配體成員[10],是由雄性睪丸支持細胞和雌性卵巢顆粒細胞分泌的一種糖蛋白[11]。AMH基因位于20 號染色體,全長4 621 bp,包括4 個CDS 區,mRNA 長3 478 bp,AMH 蛋白包括670 個氨基酸,在性別分化以及卵泡發育中扮演重要角色。Emily 等[12]發現AMH基因在不孕婦女中存在與子宮內膜異位癥相關的SNP 位點。在人AMH基因啟動子區存在多態性位點,該位點的產生導致AMH 蛋白49 位異亮氨酸取代絲氨酸,影響AMH 的生物活性[13]。王莉娜[14]在豬AMH基因外顯子區發現了插入的突變位點,攜帶插入位點的母豬產仔數顯著高于未攜帶插入位點的母豬。呂愛霞等[15]在多囊卵巢綜合癥患者中發現AMH基因146 bp 處存在G/T 突變,該突變位點與輔助生殖技術以及輔助生殖技術激素水平有著重要關系。AMH通過AMHR2(Anti-Mullerian hormone receptor type 2,AMHR2)抑制卵泡對促卵泡激素(FSH)的敏感性,抑制卵泡的生長發育[16-18]。禽類排卵依賴于成熟卵泡的數量,卵泡選擇過程的正常進行決定了禽類正常排卵[19]。本研究旨在通過對AMH基因SNP 位點與黑羽番鴨產蛋及體重的關聯分析,篩選可靠的遺傳標記,為黑羽番鴨育種工作提供理論依據。
1 材料與方法
1.1 實驗動物與性狀測定 本實驗黑羽番鴨來自陜西安大農業發展有限公司潼關育種場,選用同一批次相同條件下飼養的籠養黑羽番鴨。記錄黑羽番鴨的產蛋情況,統計300 日齡產蛋數(EN300)、280 日齡體重(280W)。隨機選出186 只籠養黑羽番鴨,翅下靜脈采血。
1.2 DNA 提取和引物設計 采用傳統酚/氯仿抽提法提取基因組DNA,根據NCBI 中的鴨AMH基因的DNA序列(NC_040065)設計PCR 引物,擴增AMH基因序列,引物由北京擎科生物科技有限公司西安分公司合成(表1)。
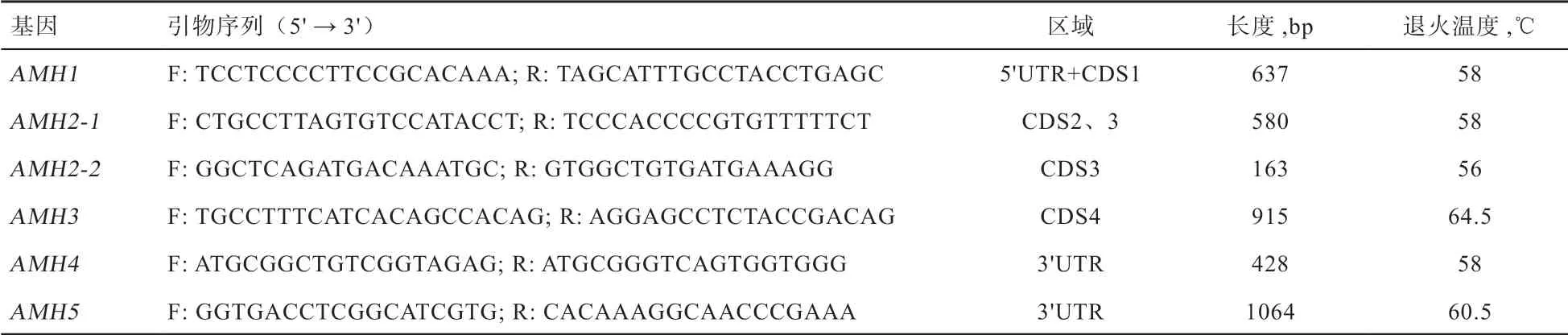
表1 AMH 基因引物信息
1.3 PCR 擴增及產物檢測 構建186 只個體DNA 混池,用引物AMH1~5 進行PCR 擴增,PCR 反應總體系20 μL:2×Taq PCR Mix 10 μL,上下游引物(10 μM)各1 μL,DNA 模板(500 ng/μL)2 μL,雙蒸水6 μL;反應程序:95 ℃ 5 min,95 ℃ 30 s,Tm 30 s,72 ℃ 1 min,35 個循環,72℃ 5 min。PCR 產物用1%的瓊脂糖凝膠電泳檢測后,送北京擎科生物科技有限公司測序。
1.4 基因分型 選取適宜的限制性內切酶(表2),判定個體的基因型。酶切總體系10 μL:6 μL PCR 產物、1 μL(10×buffer)、0.2 μL(lounits/uL)限制性內切酶、2.8 μL ddH2O。過夜消化,用3%瓊脂糖凝膠電泳分型。

表2 限制性內切酶
1.5 統計分析 計算基因頻率以及基因型頻率,進行遺傳多態性分析并利用SPSS 軟件的一般線性模型進行方差同質性檢驗以及單因素方差分析,單因素方差分析結果用“平均值±標準誤”表示。
2 結果與分析
2.1 SNPs篩選 通過測序篩選到3個SNP位點:g.1731G>A、g.2044T>C、g.3531C>T(圖1),對3 個位點進行基因分型。

圖1 AMH 基因的SNP 位點鑒定
2.2AMH基因PCR-RFLP 結果 將186 個黑羽番鴨血液DNA 樣本進行PCR 擴增后,用限制性內切酶消化目的片段,進行酶切分型。其中,g.1731G>A 可以分為GG、GA、AA 3 種基因型(圖2);g.2044T>C 可分為TT、TC、CC 3 種基因型(圖3);g.3531C>T 可分為CC、CT、TT 3 種基因型(圖4)。

圖2 g.1731G>A 電泳圖
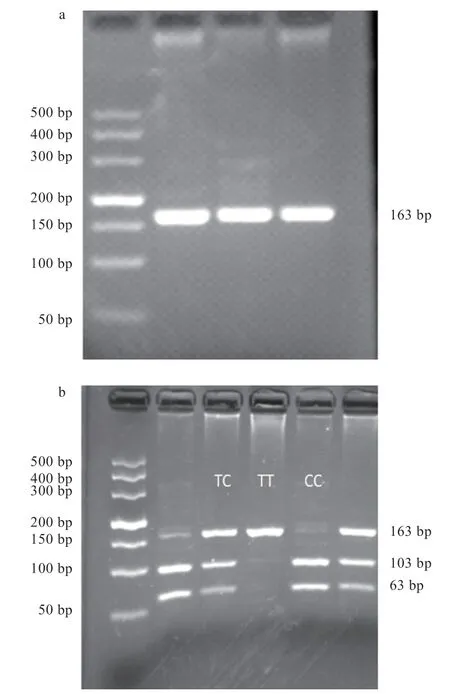
圖3 g.2044T>C 電泳圖
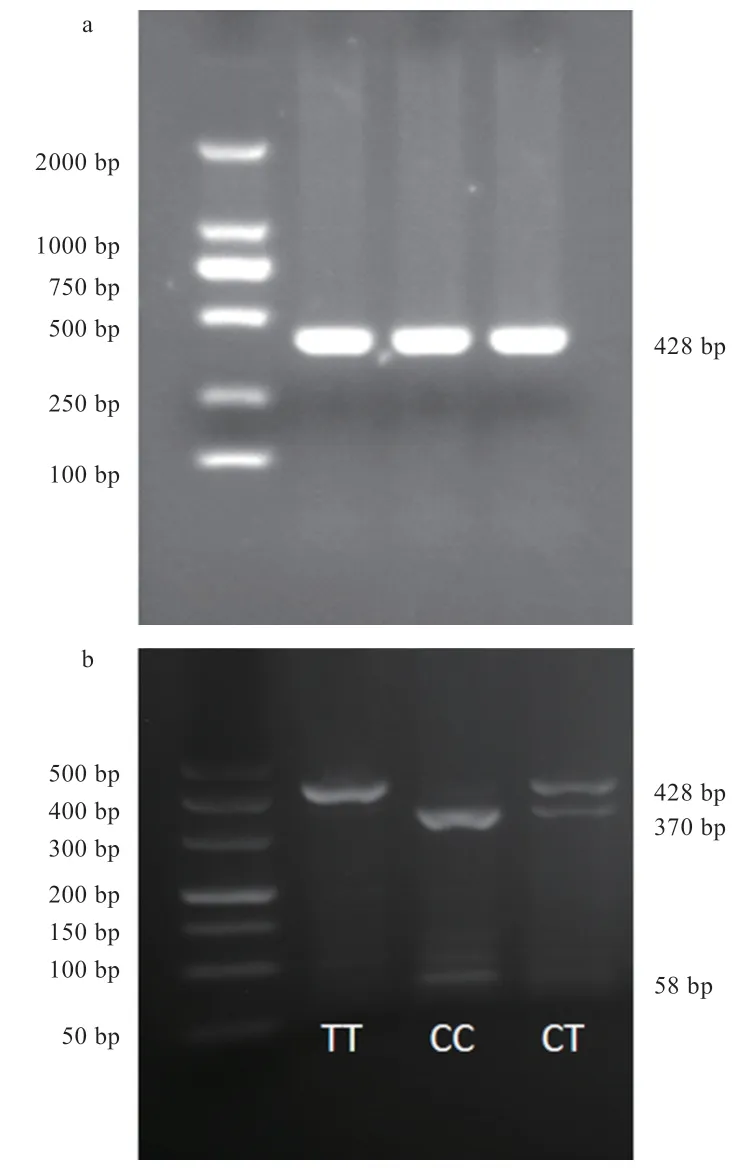
圖4 g.3531C>T 電泳圖
2.3 SNP 突變位點群體遺傳參數分析 對AMH基因的基因型頻率、基因頻率、多態信息含量(PIC)、遺傳純和度(Ho)、遺傳雜合度(He)、有效等位基因數(Ne)進行統計分析,結果見表3。經哈迪溫伯格平衡檢測3 個位點均處于平衡狀態,在g.1731G>A 位點中G 為優勢等位基因,AG 為優勢基因型,基因型頻率為AG>GG>AA;在g.2044T>C 位點中,T 為優勢等位基因,基因型頻率為CT>TT>CC,在產蛋和體重性狀中,CC 為優勢基因型;在g.3531C>T 位點中,C 為優勢等位基因,基因型頻率為CC>CT>TT。通過計算遺傳參數,g.1731G>A、g.2044T>C、g.3531C>T 位點的PIC在0.25~0.50,均為中度多態,g.1731G>A 純和度略高,雜合度略低,在群體中等位基因分布不均勻。

表3 AMH 基因群體遺傳參數
2.4AMH基因多態性與產蛋性狀和體重的關聯分析 將基因型與產蛋數據以及體重數據進行方差齊性檢驗后進行單因素方差分析,結果見表4。在分析結果中,在g.1731G>A 位點,番鴨300 日齡時,AG 基因型產蛋數顯著高于AA,AG 為優勢基因型,280 日齡體重均值約為3.1 kg,在不同基因型之間的體重均值無顯著差異,A/G 突變和體重不相關。在g.2044T>C 中,CC 基因型產蛋數顯著高于TC 和TT 型,CC 型體重顯著高于TT 型,CC 為優勢基因型。在g.3531C>T 中,3 種基因型的平均產蛋數約為44 枚,體重約為2.6 kg,不同基因型的產蛋數和體重均值無顯著差異。

表4 AMH 基因型與產蛋數和體重的相關分析
3 討 論
AMH 是TGFβ超家族成員,其主要受體為TGFβI 型受體和TGFβII 型受體[20],與受體結合后主要通過Smad、MAPK、PI3K/Akt 等多種途徑發揮作用[9]。AMH 由生長中卵泡的顆粒細胞分泌,在原始卵泡啟動中起抑制作用。若卵巢中AMH 不表達,會導致卵巢儲備過早耗盡[21],推測AMH 與黑羽番鴨繁殖性能有著密切關系。一些研究證明荷斯坦母牛中AMH 的基因組遺傳力與繁殖性狀有著重要聯系[22-24]。AMH 水平在許多哺乳動物[25-29]中已經成為預測卵巢儲備的代表。
研究基因的單核苷酸多態性引起的氨基酸變異對理解基因型和表型之間的關系具有重要意義[30]。本實驗對AMH基因在黑羽番鴨中進行SNP 鑒定,只檢測了5'UTR 以及CDS 區和3'UTR 的SNP 位點,發現了3 個SNP 位點。根據哈迪溫伯格平衡定律檢測,g.1731G>A(CDS 區)、g.2044T>C(CDS 區)、g.3531C>T(3'UTR)位點處于平衡狀態。本實驗中,對AMH基因在CDS區域的SNP(g.1731G>A、g.2044T>C)位點進行分析,發現g.1731G>A 位點G(GTA)突變至A(ATA)導致AMH編碼的第280 個氨基酸由纈氨酸變為異亮氨酸。薄秀梅等[31]研究發現MEKK3 中K391 的氨基酸突變引起蛋白結構和功能的改變。鎖培蘇等[32]在卵巢早衰患者中發現AMH 蛋白第372 位氨基酸由丙氨酸突變為酪氨酸,該位點的突變與卵巢早衰具有相關性。本實驗對AMH 的單核苷酸多態性位點與產蛋性狀和體重性狀進行關聯分析,同時關注SNP 位點的產生是否導致氨基酸的變化。本實驗結果表明,AA 基因型的平均產蛋數顯著低于AG 基因型,AA 型個體表型處于劣勢,AG 型個體表型處于優勢,為黑羽番鴨育種工作提供理論參考。通過expasy.org 網頁分析突變前后的氨基酸序列發現,G 變為A 導致纈氨酸含量由5.7%變為5.5%,異亮氨酸含量由11% 變為12%,這2 個氨基酸均為疏水性氨基酸,突變前后總平均親水性不變,AMH編碼的蛋白質不穩定系數均為54.01(將不穩定系數小于40的蛋白判定為能夠在體外穩定存在),可能間接影響了蛋白質的空間結構,AMH基因的單核苷酸突變可能影響了AMH 和AMHR2 互作[33]。本研究中,g.1731G>A 位點中AG 基因型的產蛋數均值顯著高于AA 型,但對體重的影響不顯著,在g.2044T>C 位點中,CC 基因型在產蛋和體重性狀上都有顯著優勢,番鴨中g.1731G>A和g.2044T>C 突變位點可作為遺傳標記參考基因。通過對AMH基因的5'UTR 及3'UTR 區域的SNP 檢測,發現g.3531C>T 位點位于3'UTR 區,CC 基因型頻率顯著高于TT 型,但該位點突變對產蛋和體重的影響不顯著。
本研究中g.1731G>A 和g.2044T>C 的突變位點均與產蛋性狀相關,g.2044T>C 的突變位點與體重相關,AG 基因型和CC 基因型可作為新一代黑羽番鴨育種的候選基因型,但由于產蛋和體重性狀受到多種因素影響,所以g.1731G>A 位點和g.2044T>C 位點對AMH 蛋白功能的影響有待深入研究。
4 結 論
本實驗結果表明,AMH基因存在g.1731G>A、g.2044T>C、g.3531C>T 3 個SNPs 位點。其中,g.1731G>A的AG 和g.2044T>C 的CC 為優勢基因型。g.1731G>A和g.2044T>C 與產蛋性狀相關,g.3531C>T 與產蛋性狀不相關,g.2044T>C 與體重性狀相關,g.1731G>A 和g.3531C>T 與體重性狀不相關。在哺乳動物中,AMH已經被證實可以作為預測卵巢儲備的標志物,但其是否可以作為黑羽番鴨繁殖性能的遺傳標記,還有待生產實際進一步驗證。
