地膜覆蓋對黃土高原地區兩種種植密度下玉米葉片代謝組的影響
2021-06-09 13:01:16白文波段鳳瑩趙如浪王永宏李少昆宋吉青周文彬
作物學報 2021年8期
牛 麗 白文波 李 霞 段鳳瑩 侯 鵬 趙如浪 王永宏 趙 明 李少昆 宋吉青,* 周文彬,*
地膜覆蓋對黃土高原地區兩種種植密度下玉米葉片代謝組的影響
牛 麗1,2,3白文波1李 霞2段鳳瑩2侯 鵬2趙如浪4王永宏4趙 明2李少昆2宋吉青1,*周文彬2,*
1中國農業科學院農業環境與可持續發展研究所 / 農業農村部農膜殘留污染防治重點實驗室, 北京 100081;2中國農業科學院作物科學研究所, 北京 100081;3中國農業大學水利與土木工程學院, 北京 100083;4寧夏農林科學院農作物研究所, 寧夏永寧 750105
在黃土高原地區, 對覆膜條件下玉米葉片代謝組變化規律的研究是探索覆膜增產生理機制的重要方面。本研究采用氣相色譜-四極桿-飛行時間質譜(GC-QTOF)技術, 對2種覆膜方式(塑料膜覆蓋和不覆蓋)、2個密度(7.5×104株 hm–2和10.5×104株 hm–2)和2個品種(鄭單958和先玉335)的吐絲期玉米葉片進行了代謝組學分析。從差異代謝物個數來看, 先玉335對覆膜的響應大于鄭單958, 較高的種植密度縮小了覆膜與無覆膜處理間葉片代謝的差異。主成分分析表明, 覆膜處理和品種均對代謝物組成產生了重大影響。覆膜引起的代謝譜差異主要是由檸檬酸等有機酸和氨基酸造成的; 品種引起的代謝組差異主要是由辛酸等烷酸和酚類造成的。相關分析表明, 葉綠醇、白藜蘆醇、葡萄糖-6-磷酸與玉米籽粒產量呈顯著正相關關系, 而甘油與之呈顯著負相關關系。在覆膜條件下, 與呼吸作用及清除光呼吸產物相關的纈氨酸、異亮氨酸及蛋氨酸, 與三羧酸循環相關的異檸檬酸以及可減輕光抑制的蛋氨酸、N-乙酰基天冬氨酸等代謝物的水平整體呈升高趨勢。以上結果表明, 在覆膜條件下, 消除光呼吸產物及減少光抑制的相關代謝物的積累是葉片凈光合速率提高的代謝基礎; 抗氧化及能量代謝相關的代謝物在提高產量的過程中發揮了重要作用。
地膜覆蓋; 葉片; 代謝組; 產量; 抗氧化; 玉米
近年來, 氣候變化對世界主要農作物的產量產生了一些負面影響[1]。隨著全球氣候持續變暖, 干旱的發生頻率將會增加[2]。為保證糧食安全, 滿足人們日益增長的糧食需求, 農業生產需要迅速適應氣候變化, 以減緩或消減氣候變化帶來的不良效應。適當的作物管理措施是實現這一目標的主要途經[3]。黃土高原是我國典型的干旱和半干旱氣候區, 年均降雨量為250~600 mm, 當地農業以旱地雨養農業為主[4]。該區太陽輻射充足, 經常出現春季低溫, 作物生育期內降雨不足且難以預測。因此, 低溫、缺水和強蒸發是限制該地區作物生產的主要因素[5]。提高作物水分利用效率(water use efficiency, WUE)對當地農業的可持續發展和糧食安全至關重要[6]。在過去幾十年中, 已經發展形成了多種農田管理措施來提高水分利用效率, 如雨水收集措施[7]、地膜(塑料薄膜)覆蓋、農作物秸稈覆蓋[8]、礫石覆蓋[9]以及滴灌措施[10]等。地膜覆蓋作為最有效的措施之一, 可提高雨養農業區作物的水分利用效率, 在玉米、小麥、棉花和馬鈴薯等作物上得到廣泛應用[11-14]。
玉米是黃土高原的主要作物之一, 其種植面積占黃土高原耕地面積的27.3%[15]。近年來, 地膜覆蓋因其減少土壤水分蒸發、提高土壤含水量和表土溫度等作用已被廣泛應用于該地區的玉米種植[16-18]。此外, 地膜覆蓋可以通過增加陽光的反射率和膜下空氣流動的阻力來改善土壤表面的微氣候[19], 以及減少雜草和害蟲的危害[20]。在干旱和半干旱地區, 前人對地膜覆蓋在作物生產中的應用研究主要集中在土壤–植物–大氣連續體(soil-plant-atmosphere continuum, SPAC)的水熱傳輸情況、作物的生長發育、產量、品質和水分利用效率等指標的量化上。研究表明, 地膜覆蓋通過提高土壤含水量和土壤溫度, 提高了作物產量, 改善了作物品質, 同時提高了水分利用效率[11,21]。在生理水平上, 地膜覆蓋條件下的作物比無地膜覆蓋的作物表現出, 更高的葉綠素含量[21]、核酮糖-1,5-二磷酸羧化酶(RubisCO)活性[22]、抗氧化活性[23]、電子傳遞速率[24]和熱耗散[21], 故具有較強的光合能力。
作物原初代謝與產量密切相關。近年來, 特定環境條件下作物代謝標記物篩選的研究引起了人們的關注[25-27]。前人采用289個玉米自交系對與玉米農藝性狀相關的代謝組進行全基因組關聯分析發現, 木質素前體含量與木質素含量、植株高度及干物質產量顯著相關, 這表明代謝組可用來預測農藝性狀, 建立基因型和表型之間的關聯[25]。Witt等[26]研究表明, 玉米在干旱脅迫下, 氨基酸、糖、糖醇和三羧酸循環的中間體代謝物含量普遍增加, 這種趨勢并沒有因為品種耐旱性的改變而變化。水稻代謝組研究也鑒定到了抗旱性的代謝標記候選物絲氨酸、蘇氨酸和蘇糖酸[27]。大田中小麥和玉米的農藝性狀和代謝數量性狀位點定位在相同的基因組區域, 表明了代謝性狀在品種選育中的重要作用[25,28]。
玉米產量既取決于葉片對籽粒的同化物供應(源), 也取決于籽粒容納這些同化物的潛力(庫)[29]。目前, 玉米抗逆品種的選育主要關注庫潛力的提升[30],而源潛力提升機制仍需要進一步深入研究。水分脅迫影響葉片的生理和代謝, 尤其影響光合作用, 從而降低葉片的源能力[31]。與其他器官相比, 水分脅迫對葉片的代謝產物組成的影響最大[26]。葉片代謝和源能力密切相關, 故葉片代謝組與籽粒產量之間存在密切關系。在葉片結構上, 地膜覆蓋條件下生長的玉米葉片, 具有更高的氣孔密度, 因此具有較高的凈光合速率[32]。密植是玉米增產的重要農藝措施, 但密植通常使玉米葉片葉綠體形態發生變化, 葉綠體及基粒數量減少, 葉綠素、含量降低, 凈光合速率降低, 葉肉細胞的膜結構逐漸受損[33]。綜上所述, 從代謝組學水平分析栽培措施對作物的影響, 可以系統地揭示特定栽培措施條件下作物產量形成的生理代謝基礎, 獲得產量的代謝標記物。然而, 地膜覆蓋對作物代謝組的影響還鮮有報道。本文通過研究不同密度和品種的玉米葉片代謝組對地膜覆蓋的響應, 以期探明地膜覆蓋提高玉米產量和水分利用效率的代謝基礎, 篩選出與產量密切相關的代謝標記物, 為黃土高原旱作區玉米栽培和品種選育提供理論依據和研究方向。
1 材料與方法
1.1 試驗地點
田間試驗于2018年在寧夏固原市原州區進行。該地屬黃土高原丘陵地區(36°18'53''N, 106°22'20''E), 海拔1769 m, 屬中溫帶半干旱氣候區, 年平均降水量約為400 mm, 且季節分布不均, 約60%的降水集中在7月至9月。年日照時數、年平均氣溫和無霜期分別為2518.3 h、6.2°C和151 d。試驗地土壤類型為褐土, 0~40 cm土層土壤容重為1.27 g cm?3, 土壤有機質7.86 g kg?1, 總氮0.81 g kg?1, 堿解氮55.6 mg kg?1, 速效磷12.8 mg kg?1, 速效鉀158.0 mg kg?1。
1.2 試驗設計和田間管理
試驗采用裂區試驗設計, 主區為覆膜方式, 裂區為種植密度, 裂裂區為品種。主區覆膜方式為: (1)寬(80 cm)窄(40 cm)行壟溝全膜覆蓋(FM, 圖1-A), (2) 寬(80 cm)窄(40 cm)行壟溝無覆蓋(NM, 圖1-B); 裂區種植密度為: (1) 低密度7.5×104株 hm–2; (2) 高密度10.5×104株hm–2; 裂裂區玉米品種為: (1)鄭單958 (Zhengdan 958, ZD958); (2) 先玉335 (Xianyu 335, XY335)。鄭單958和先玉335是在中國廣泛種植的2個玉米栽培品種, 低密度7.5×104株 hm–2目前在中國許多地區被推薦種植, 而高密度10.5×104株hm–2通常可以獲得更高的產量[34]。本試驗以覆膜方式為基礎, 設置不同的栽培品種和密度組合, 共包括以下8個處理: (1) FMZL, 地膜覆蓋低密度種植的ZD958; (2) FMZH, 地膜覆蓋高密度種植的ZD958; (3) FMXL, 地膜覆蓋低密度種植的XY335; (4) FMXH, 地膜覆蓋高密度種植的XY335; (5) NMZL, 無覆蓋低密度種植的ZD958; (6) NMZH, 無覆蓋高密度種植的ZD958; (7) NMXL, 無覆蓋低密度種植的XY335; (8) NMXH, 無覆蓋高密度種植的XY335。
玉米生長在寬窄行壟溝結構的溝中。A: FM, 寬(80 cm)窄(40 cm)行壟溝全膜覆蓋; B: NM, 寬(80 cm)窄(40 cm)行壟溝無覆蓋。
The maize was grown in the furrows of ridge-furrow construction with wide - narrow rows. A: FM, ridge-furrow construction with alternating wide (80 cm) and narrow (40 cm) ridges and full plastic film mulching; B: NM, ridge-furrow construction with alternating wide (80 cm) and narrow (40 cm) ridges and no mulching.
供試小區行長9.6 m, 寬4 m, 小區面積38.4 m2, 每個處理設3次重復, 共24個小區。在取樣和測量時, 將每個小區1 m的邊緣排除在外。試驗前對地塊進行旋耕、平整, 播種前9 d 進行施肥、起壟和地膜覆蓋。于2018年4月27日, 用點播器將2粒玉米種子播于溝槽底部5 cm深處。播種后1個月進行間苗, 每穴留1株幼苗。播種前施用尿素、磷酸氫二銨和硫酸鉀作為基肥, 折合純N 123 kg hm?2, P2O5138 kg hm?2, K2O 75 kg hm?2。拔節期追施尿素和磷酸氫二銨以防止養分脅迫, 折合純氮200 kg hm?2, P2O569 kg hm?2。按照當地常規管理措施進行雜草和病蟲害防控。玉米整個生育期內除自然降水外無人工灌水, 玉米于2018年10月3日收獲。
1.3 取樣與測量方法
1.3.1 葉片代謝組測定 于玉米吐絲期, 每小區隨機選擇6株玉米植株, 在晴朗無云的上午9:00—9:30, 取穗位葉快速放入液氮中, 用干冰運回實驗室, 存放于-80°C超低溫冰箱。將約50 mg冷凍研磨的玉米葉片樣品與700 μL甲醇(100%)和30 μL核糖醇內標儲備液(0.2 mg mL–1)混合后, 在70°C恒溫混合器(Eppendorf)中震蕩15 min。以14,000轉 min–1離心10 min后, 將上清液轉移至新離心管中。加入400 μL氯仿和800 μL水, 然后將離心管小心渦旋15 s, 以14,000轉 min–1離心15 min。從上部極性相中吸取2等份50 μL上清液至1.5 mL新離心管中, 放置真空濃縮儀(Eppendorf)室溫干燥過夜。制備完成的極性初級代謝物經衍生化后, 通過氣相色譜-四極桿-飛行時間質譜儀(GC-QTOF)進行代謝組分析[35]。所得的質譜保留時間(RI)使用TagFinder軟件以及數據庫Golm Metabolome Database (GMD)進行代謝物的鑒定及量化[36-37]。原始檢測豐度通過內標(核糖醇)和樣品鮮重進行標準化。將所有代謝組學數據除以所有品種和處理中每種代謝物的中值對數據進行歸一化, 并對得到的數值進行log2轉化, 得到的數據用于進一步的分析。篩選組間差異代謝物時采用單維分析和多維分析相結合的方法。在正交偏最小二乘方-判別分析(OPLS-DA分析)中, 變量權重值(VIP)可用來表示各代謝物的含量對組間樣本分類判別的解釋能力和影響強度, 一般認為VIP>1的代謝物為差異代謝物。同時利用T檢驗對組間差異代謝物進行檢驗。故篩選差異代謝物的標準為VIP>1且-value<0.05。采用具有數據分析、數據可視化及數據處理功能的軟件MultiExperiment Viewer (version 4.8.1, the Institute of Genomic Research, Rockville, MD, USA)基于Pearson相關性進行層次聚類。使用R software (http://www.r-project.org/; R package pca Methods)進行概率主成分分析(PCA)。使用SPSS 12.5 (SPSS Inc., Chicago, IL, USA)進行相關性分析。
1.3.2 單株產量 在成熟期, 每個小區排除1 m的邊界, 隨機選取15株收獲測產。以14%的籽粒含水量來計算籽粒產量, 用整株干重來表示生物量。
2 結果與分析
2.1 地膜覆蓋對玉米葉片代謝組的影響
將經過標準化且log2對數轉換后的代謝組數據進行主成分分析(PCA)以得到不同處理間代謝組差異的總體輪廓。由圖1可知, 第一主成分(PC1)和第二主成分(PC2)共同貢獻了總方差的48.1%。其中ZD958和XY335兩個品種間的差異主要是通過PC1來區分(34.5%), 覆膜和無覆膜處理的玉米葉片代謝組差異主要通過PC2被區分開來(13.6%), 而不同種植密度間的差異不明顯。以上結果表明代謝組之間的差異主要來源于品種間差異以及覆膜和無覆膜處理的差異(圖2-A)。
載荷代表不同代謝物對于每個主成分的貢獻, 較高的正載荷和負載荷分別代表相應的代謝物與某一主成分的得分具有較高的正負相關性。圖2-B~C顯示了PC1 (區分品種)和PC2 (區分覆膜和無覆膜處理)的前10個正負載荷。PC1正載荷的代謝物在ZD958中水平較高, 而在XY335中水平較低, 而負載荷的代謝物水平表現出相反的趨勢。同時, PC2正載荷的代謝物水平在覆膜處理中較高, 在無覆膜處理中較低, 而負載荷的代謝物水平表現出相反的趨勢。在第一主成分上導致代謝譜分離的代謝物主要有辛酸等烷酸和對苯二酚等酚類, 在第二主成分上導致代謝組分離的代謝物主要有檸檬酸等有機酸和蛋氨酸等氨基酸。
為了深入了解不同處理對玉米葉片代謝組的影響, 通過對覆膜或不覆膜處理、不同密度及不同品種之間進行兩兩比較, 以VIP >1且< 0.05為閾值, 篩選出具有顯著差異的代謝物。從差異代謝物數量來看, 地膜覆蓋因素和品種因素造成的差異代謝物數目較多, 而密度因素造成的差異代謝物的數目較少。XY335對地膜覆蓋的響應大于ZD958。密度對代謝物的影響在覆膜條件下較大(圖3)。
ZD958和XY335對覆膜的響應存在一定的共性, 但這種共性會因密度而異。低密度條件下, 與無覆膜相比, ZD958和XY335覆膜處理后產生25個共同差異代謝物; 高密度條件下, 這 2個品種在覆膜后與無覆膜相比產生14個共同的差異代謝物。在低密度和高密度情況下, ZD958品種進行覆膜處理時產生了26個共同的差異代謝物; 而XY335品種在這2種密度下進行覆膜處理時, 產生了23個共同的差異代謝物。綜合不同密度和不同品種的共性, 覆膜會引起10種代謝物的差異, 其中對苯二酚、3,4-二羥基苯甲酸、核糖酸、香草酸、尿嘧啶和1,3-二羥基吡啶含量降低, 而纈氨酸、蛋氨酸、異檸檬酸和N-乙酰基天冬氨酸含量升高(圖4)。
A: 第一主成分(PC1)和第二主成分(PC2)的得分圖; B: PC1前十位正載荷和負載荷的代謝物; C: PC2前十位正載荷和負載荷的代謝物。FMZL: 地膜覆蓋低密度種植的ZD958; FMZH: 地膜覆蓋高密度種植的ZD958; FMXL: 地膜覆蓋低密度種植的XY335; FMXH: 地膜覆蓋高密度種植的XY335; NMZL: 無覆蓋低密度種植的ZD958; NMZH: 無覆蓋高密度種植的ZD958; NMXL: 無覆蓋低密度種植的XY335; NMXH: 無覆蓋高密度種植的XY335。其中綠色色塊代表較低的含量, 紅色色塊代表較高的含量。
A: scores of the first principal component (PC1) and the second principal component (PC2). B: the top ten negative and metabolites of PC1. C: the top ten negative and metabolites of PC2. FMZL: ZD958 with low planting density and plastic film mulching; FMZH: ZD958 with high planting density and plastic film mulching; FMXL: XY335 with low planting density and plastic film mulching; FMXH: XY335 with high planting density and plastic film mulching; NMZL: ZD958 with low planting density and no mulching; NMZH: ZD958 with high planting density and no mulching; NMXL: XY335 with low planting density and no mulching; NMXH: XY335 with high planting density and no mulching. Among them, green represents lower expression abundance, and red represents higher expression abundance.
處理同圖2。Treatments are the same as those given in Fig. 2.
以低密度種植的ZD958品種為例, 地膜覆蓋使41種代謝物的含量發生了變化。對這些代謝物進行層次聚類分析發現, 代謝物的含量在地膜覆蓋處理和無覆蓋處理之間存在著較大的差異。此外, 在不同品種和密度之間也存在一定的差異(圖5)。根據不同處理間代謝物含量的變化趨勢, 可將代謝物分為以下3類: 第一類是在進行地膜覆蓋處理后含量升高的代謝物, 主要為氨基酸、糖酵解過程產物及抗氧化物質, 包括纈氨酸、蛋氨酸、葡萄糖-6-磷酸、異檸檬酸、磷酸烯醇式丙酮酸、白藜蘆醇、葉綠醇和異亮氨酸等物質。總體來說, XY335的此類代謝物含量低于ZD958; 高密度條件下此類代謝物的含量低于低密度條件。第二類是在進行地膜覆蓋處理后, 含量變化不大的代謝物, 主要為一些糖類, 包括巖藻糖、3,6-脫水-D-葡萄糖和核糖酸, 總體趨勢為XY335的含量高于ZD958。第三類是在進行地膜覆蓋處理后含量降低的代謝物, 主要為有機酸和滲透調節相關的物質, 包括檸檬酸、谷氨酸、琥珀酸、對苯二酚、甘油和肌酐等物質。在地膜覆蓋條件下, XY335的此類代謝物含量高于ZD958; 高密度條件下的此類代謝物含量高于低密度。在無地膜覆蓋條件下, XY335的此類代謝物含量低于ZD958。
處理同圖2。數字表示2組處理間具有顯著差異的代謝物數目。
Treatments are the same as those given in Fig. 2. The numbers indicate the numbers of metabolites with significant differences between the two treatments.
處理同圖2。其中綠色色塊代表較低的含量, 紅色色塊代表較高的含量。
Treatments are the same as those given in Fig. 2. Among them, green represents lower expression abundance, and red represents higher expression abundance.
2.2 地膜覆蓋對玉米葉片代謝網絡的影響
以低密度種植的ZD958品種為例, 在主要代謝通路中, 覆膜使16種已注釋代謝物的含量產生了變化。綜合所有處理來看, 在覆膜條件下整體上調的代謝物有8個, 分別是葡萄糖-6-磷酸、蛋氨酸、磷酸烯醇丙酮酸、異亮氨酸、多巴、纈氨酸、N-乙酰基天冬氨酸和異檸檬酸。其中, 葡萄糖-6-磷酸和磷酸烯醇丙酮酸是和糖酵解過程密切相關的代謝物。在覆膜條件下整體下調的代謝物有8個, 分別是甘油、O-磷酸-L-絲氨酸、檸檬酸、對苯二酚、琥珀酸、N-乙酰鳥氨酸、肌酐和谷氨酸, 其中肌酐和脯氨酸代謝相關, 而甘油和脯氨酸都屬于滲透調節物質; O-磷酸-L-絲氨酸與光呼吸的產物絲氨酸可以相互轉化(圖6)。
2.3 玉米產量與代謝物水平的相關性分析
對單株籽粒產量及產量構成的分析表明, 地膜覆蓋條件下, ZL、ZH、XL和XH的單株籽粒產量分別比無覆蓋條件下增加了12.70%、22.14%、23.37%和4.87%。從產量組成要素來看, 地膜覆蓋對產量的提高主要是通過增加每穗籽粒數來實現的, 其對ZL、ZH、XL和XH每穗籽粒數的提高幅度分別為17.02%、10.61%、16.87%和18.32%。方差分析表明, 覆膜、密度和品種均對單株產量產生顯著影響。覆膜和密度、密度和品種及覆膜、密度和品種三者對單株籽粒產量的影響存在交互作用。對于鄭單958品種, 覆膜對高密度下的單株籽粒產量的提高幅度較大, 而對于先玉335品種, 則是對低密度下單株籽粒產量的提高幅度較大(表1)。
紅色圓代表在地膜覆蓋處理下總體上調的物質, 綠色圓代表在地膜覆蓋處理下總體下調的物質。8個不同的色塊從左到右依次代表FMZL、FMZH、FMXL、FMXH、NMZL、NMZH、NMXL、NMXH, 處理同圖2。其中綠色色塊代表較低的含量, 紅色色塊代表較高的含量。
Red circles represent substances that are generally up-regulated under plastic film mulching treatment, and green circles represent substances that are generally down-regulated under plastic film mulching treatment. Eight different color blocks from left to right represent FMZL, FMZH, FMXL, FMXH, NMZL, NMZH, NMXL, and NMXH, respectively. Treatments are the same as those given in Fig. 2. Among them, green block represents lower expression abundance, and red represents higher expression abundance.
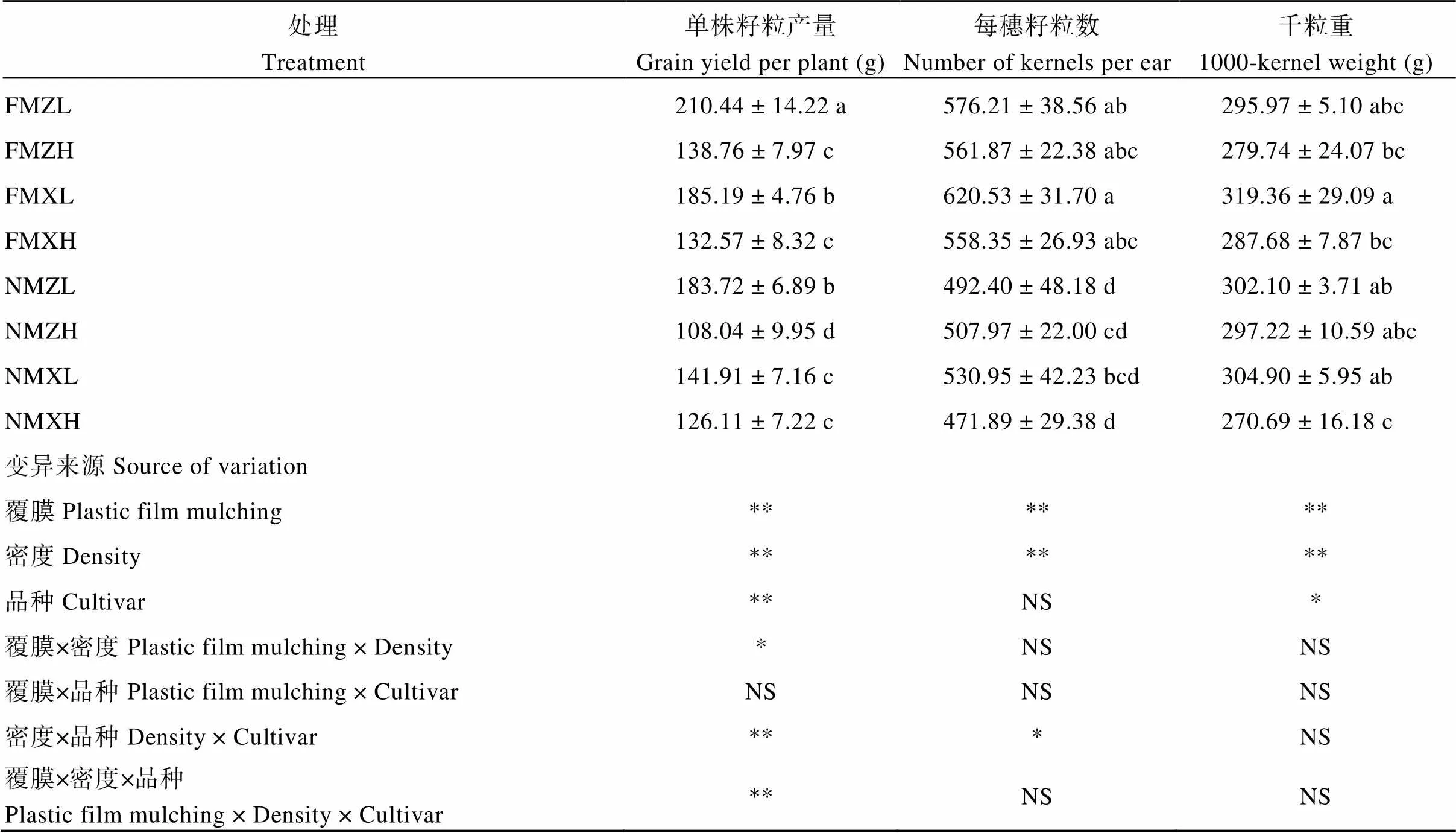
表1 玉米單株籽粒產量及產量構成要素
數據代表平均值±標準差, 采用Duncan’s多重比較, 每一列中不同的小寫字母表示處理之間在0.05水平上差異顯著。處理同圖2; 方差分析表中*表示顯著差異(*< 0.05; **< 0.01); NS, 不顯著。
Values represent means ± SD, values within a column followed by different lowercase letters indicate significant differences at< 0.05 by Duncan’s multiple range test. Treatments are the same as those given in Fig. 2. * and ** indicate significant differences at the0.05 and 0.01 probability levels, respectively. NS, not significant.
進一步將玉米籽粒產量及生物量與其代謝物含量進行相關性分析, 以期篩選出與產量直接相關的代謝標記物。結果表明, 白藜蘆醇、葉綠醇和葡萄糖-6-磷酸與玉米籽粒產量呈顯著正相關關系, 而甘油與之呈顯著負相關關系(圖7)。此外, 甘油與玉米生物量呈顯著負相關關系(圖8)。說明在產量較高的玉米植株中, 白藜蘆醇、葉綠醇和葡萄糖-6-磷酸這3種代謝物的含量也高度積累, 其可作為潛在的產量相關代謝標記物, 用于優良品種的選育。
3 討論
3.1 農藝措施與代謝組之間的關系
已有研究提出采用相關標記物的策略用于分子標記輔助育種[38]。基于GC-MS的代謝組具有高通量、穩健性以及分析主要代謝物的能力[39], 在分子標記輔助育種方面具有良好的前景。本文采用這一方法對不同處理的玉米葉片代謝組進行了研究, 在覆膜、密度和品種這3個因素中, 對代謝組影響最大的是品種, 其次是覆膜, 影響最弱的是密度(圖2和圖3)。這從代謝層面說明了選育抗逆高產品種的重要性。從差異代謝物的數目來看, 較高的種植密度縮小了覆膜與無覆膜處理間的差異(圖3和圖4), 這可能是由于較高的種植密度使單株玉米葉片光合能力變差、膜結構受損, 從而導致代謝變差導致[33]。長期以來, 人們認為氨基酸積累是對非生物脅迫的標志性反應[40]。干旱脅迫期間氨基酸譜的變化歸因于各種機制, 包括蛋白質合成減少[41], 蛋白質降解增加[42], 以及受底物和酶活性調節的生物合成。氨基酸在植物抗逆性中起著重要的作用[43], 研究者在干旱脅迫[39]和滲透脅迫[42]條件下的植物葉片中發現許多氨基酸的積累。在本研究中, 無地膜覆蓋條件下, 玉米葉片普遍積累的氨基酸有O-磷酸-L-絲氨酸、N-乙酰基鳥氨酸和谷氨酸(圖5)。O-磷酸-L-絲氨酸可以和絲氨酸相互轉化, 而絲氨酸是光呼吸的產物, 可在一定程度上反映光呼吸的強弱[44]。葉片中O-磷酸-L-絲氨酸的積累表明葉片光呼吸較強。N-乙酰基鳥氨酸是從谷氨酸到精氨酸的酶促生物合成過程中間體。谷氨酸在植物體內含氮化合物的轉化中具有重要作用, 它可以轉化為脯氨酸。在無地膜覆蓋的玉米葉片中觀察到了谷氨酸的積累(圖5), 一方面谷氨酸可作為滲透調節物質起作用, 另一方面谷氨酸可作為前體物質轉化成脯氨酸[45]。脯氨酸可以調節滲透壓, 清除自由基, 作為親水物質減少蛋白和酶類的降解, 從而緩解干旱對葉片細胞的傷害[46]。
和分別是通過Pearson相關分析得出的相關系數和值。
andare the correlation coefficient and-value from Pearson correlation analysis, respectively.
和分別是通過Pearson相關分析得出的相關系數和值。
andare the correlation coefficient and-value from Pearson correlation analysis, respectively.
3.2 地膜覆蓋使葉片脅迫保護氨基酸積累、光抑制減輕
支鏈氨基酸(纈氨酸和異亮氨酸)以及其他與支鏈氨基酸共享合成途徑的氨基酸(蛋氨酸)通常會在各種非生物脅迫條件下積累[39]。它們被認為是相溶性溶質[47]以及呼吸電子傳遞鏈中的電子供體[48]。本研究中, FM處理下纈氨酸、異亮氨和蛋氨酸的含量增加(圖5), 這與FM處理下葉片呼吸作用的增加有關[32]。琥珀酸是連接三羧酸循環和谷氨酸分流的代謝物[49], 因此這2種途徑之間的平衡可能影響琥珀酸的含量。在FM處理下, 琥珀酸和谷氨酸的含量均呈現降低的趨勢, 而三羧酸循環中異檸檬酸含量呈升高的趨勢(圖5和圖6), 表明在FM處理下葉片可能通過降低谷氨酸代謝來維持三羧酸循環。FM處理使玉米葉片中蛋氨酸含量普遍增加(圖5和圖6), 蛋氨酸可能與脅迫保護有關, 干旱脅迫條件下, 葉片蛋氨酸含量較高的小麥品種具有較高的產量[50], 由此可見, 蛋氨酸對FM處理下玉米產量的提高也有一定的貢獻。
在蛋氨酸和其他天冬氨酸衍生氨基酸的生物合成過程中會消耗還原力三磷酸腺苷(ATP)和還原型輔酶I (II)——NAD(P)H, 這有利于維持能量穩態, 防止光系統II (PSII)因過度還原而產生氧化應激損傷(即光抑制)[50-51]。天冬氨酸家族氨基酸的生物合成可能會起到一種能量庫的作用來緩解葉綠體能量失衡。研究表明, 高絲氨酸脫氫酶是控制天冬氨酸從天冬酰胺向天冬氨酸家族氨基酸生物合成的酶, 在干旱脅迫條件下, 葉綠體NADPH/NADP+比值通常會升高, 此時蘇氨酸對該酶的變構反饋抑制顯著減輕從而可以消耗過多的NADPH[52]。在FM處理條件下, 蛋氨酸和N-乙酰基天冬氨酸含量增加(圖5和圖6), 這意味著FM減輕了葉片的光抑制, 從而提高了葉片的凈光合速率[32]。蛋氨酸是絲氨酸的下游產物, 它的生物合成對植物的另一種應激保護機制可能是對光呼吸產物絲氨酸的消耗, 蛋氨酸合成會使光呼吸的反應平衡向右移動(遠離2-磷酸乙醇酸、乙醇酸、乙醛酸、甘氨酸和絲氨酸), 從而維持卡爾文循環, 抑制光呼吸中間體(2-磷酸乙醇酸、乙醇酸和乙醛酸)的產生[50], 在本研究中, 地膜覆蓋條件下玉米葉片中O-磷酸-L-絲氨酸的含量降低(圖5和圖6)而該物質可與絲氨酸相互轉化, 這進一步表明地膜覆蓋使玉米葉片光呼吸減弱。
3.3 產量與代謝物之間的相關性
篩選代謝標記物用于大田作物產量的指示物對抗逆品種的選育具有重要意義[53-54]。在冷脅迫條件下, 玉米的生物量與葉片棉子糖含量呈負相關[53]; 在花期干旱脅迫條件下, 玉米葉片中的甘氨酸和肌醇含量與玉米籽粒呈顯著負相關[54]。在本試驗中, 和玉米產量密切相關的代謝物主要包括: 葉綠醇、白藜蘆醇、葡萄糖-6-磷酸和甘油(圖7)。相比于無覆蓋處理, 地膜覆蓋提高了作物產量[32]。白藜蘆醇可以清除活性氧自由基, 減輕膜脂的過氧化程度[55], 從而有利于產量的提高(表1)。葉綠醇作為葉綠素的組分, 可在一定程度上反映葉綠素的變化趨勢。此外, 葉綠醇也具有抗氧化作用[56]。作為糖酵解和磷酸戊糖途徑的重要參與者, 葡萄糖-6-磷酸在為生命活動提供能量的過程中發揮了重要作用。因此, 以上代謝物可能在栽培措施帶來的增產中發揮了重要作用, 是不同栽培措施條件下與產量相關的重要代謝指示物。通過逐步回歸分析, 得到了籽粒產量(GY)與白藜蘆醇含量(MLRes)的模型關系GY=120.6× MLRes-52.6 (=0.829,=0.011)。由于樣品是從難以控制生長條件的大田獲得, 會受到多種因素的干擾, 通常認為絕對值在0.72到0.88之間,低于0.05或0.01是高度可信的, 之前在大田中進行的一些代謝組研究也得到了類似的結果[27]。甘油可防止羥基自由基的產生, 清除活性氧中間體, 從而保護細胞膜免受氧化傷害, 在脅迫條件下是一種理想的胞內抗脅迫保護劑[57]。在本研究中, 甘油含量與籽粒產量和生物量呈顯著負相關(圖8), 這表明, 較少的抗脅迫保護劑的生成有利于產量的提高。目前來看, 代謝組研究通常用于評估代謝物組成中明顯的自然差異, 即用于尋找代謝標記物的植物通常處于同一種生長條件下[25-27]。然而, 加入其他的因素會不會使原來的產量代謝標記物發生改變尚不清楚。本研究中的栽培管理措施涉及地膜覆蓋、密度和品種, 這些因素帶來的差異不僅僅屬于自然差異, 在這種情況下鑒定到的代謝物與產量之間的相關性則具有較高的保守性; 另一方面, 此類研究也可為栽培措施調控作物生理代謝的相關研究提供一定的參考。
4 結論
本文對黃土高原地區2個覆膜處理、2個種植密度及2個品種的玉米葉片代謝組進行了研究。先玉335對覆膜的響應程度(差異代謝物數目)大于鄭單958, 較高的種植密度縮小了覆膜與無覆膜處理間葉片代謝組的差異。另外, 在覆膜條件下, 可消除光呼吸產物及可減輕光抑制的蛋氨酸、N-乙酰基天冬氨酸等代謝物的水平整體呈升高趨勢, 這一現象在代謝水平上解釋了覆膜條件下玉米葉片凈光合速率提高的原因, 進一步豐富了覆膜提高玉米產量的生理機制。通過相關性分析, 發現與籽粒產量及生物量密切相關的代謝標記物主要是與抗氧化作用及能量代謝相關的代謝物, 從而揭示了此類代謝物在提高產量過程中的重要貢獻。
[1] Lobell D B, Schlenker W, Costa-Roberts J. Climate trends and global crop production since 1980., 2011, 333: 616–620.
[2] Sherwood S, Fu Q. Climate change: a drier future?, 2014, 343: 737–739.
[3] Iglesias A, Garrote L. Adaptation strategies for agricultural water management under climate change in Europe., 2015, 155: 113–124.
[4] Li F M, Wang P, Wang J, Xu J Z. Effects of irrigation before sowing and plastic film mulching on yield and water uptake of spring wheat in semiarid Loess Plateau of China., 2004, 67: 77–88.
[5] Li S X, Wang Z H, Malhi S S, Li S Q. Nutrient and water management effects on crop production, and nutrient and water use efficiency in dryland areas of China., 2009, 102: 223–265.
[6] Liu Q F, Chen Y, Liu Y, Wen X X, Liao Y C. Coupling effects of plastic film mulching and urea types on water use efficiency and grain yield of maize in the Loess Plateau, China., 2016, 157: 1–10.
[7] Mzirai O B, Tumbo S D. Macro-catchment rainwater harvesting systems: challenges and opportunities to access runoff., 2010, 7: 789–800.
[8] 張萬恒, 張恒嘉, 王澤義, 高佳. 秸稈覆蓋方式對西北旱作馬鈴薯產量效益的影響. 農業工程, 2018, 8(12): 85–88. Zhang W H, Zhang H J, Wang Z Y, Gao J. Effect of straw mulching on yield of potato in drought region of Northwest China., 2018, 8(12): 85–88 (in Chinese with English abstract).
[9] 唐學芬, 趙文舉, 李宗禮, 郁文. 壓砂地土壤鹽分空間變異規律. 水土保持通報, 2016, 36(3): 18–23. Tang X F, Zhao W J, Li Z L, Yu W. Variation pattern of soil Sali-nity of gravel-sand mulched field., 2016, 36(3): 18–23 (in Chinese with English abstract).
[10] 李憑峰, 譚煌, 王嘉航, 楊培嶺. 滴灌水肥條件對櫻桃產量、品質和土壤理化性質的影響. 農業機械學報, 2017, 48(7): 236–246. Li P F, Tan H, Wang J, Yang P L. Effects of drip irrigation water and fertilizer conditions on cherry yield, quality and soil physical and chemical properties., 2017, 48(7): 236–246 (in Chinese with English abstract).
[11] Zhang X D, Yang L C, Xue X K, Kamran M, Ahmad I, Dong Z Y, Liu T N, Jia Z K, Zhang P, Han Q F. Plastic film mulching stimulates soil wet-dry alternation and stomatal behavior to improve maize yield and resource use efficiency in a semi-arid region., 2019, 233: 101–113.
[12] 王星垚, 李援農, 銀敏華, 徐袁博, 任全茂, 徐路全. 不同地膜對土壤水熱狀況和冬小麥植株氮素的影響. 節水灌溉, 2018, (2): 40–46. Wang X Y, Li Y N, Yin M H, Xu Y B, Ren Q M, Xu L Q. Effects of different mulch films on soil hydrothermal regimes and the plant nitrogen accumulation and translocation of winter wheat., 2018, (2): 40–46 (in Chinese with English abstract).
[13] 李昊, 李世平, 南靈, 李河, 郭清卉. 中國棉花地膜覆蓋產量效應的meta分析. 農業機械學報, 2017, 48(7): 228–235. Li H, Li S P, Nan L, Li H, Guo Q H. Meta-analysis of effect of plastic film mulching on cotton yield in China., 2017, 48(7): 228–235 (in Chinese with English abstract).
[14] Hou F Y, Dong S X, Xie B T, Zhang H Y, Li A X, Wang Q M. Mulching with plastic film improved the root quality of summer-sown sweet potato ((L.) Lam.) in Northern China., 2019, 18: 982–991.
[15] 薛吉全, 張仁和, 李鳳艷, 張興華. 陜西玉米育種現狀、問題與對策. 玉米科學, 2008, 16(2): 144–146. Xue J Q, Zhang R H, Li F Y, Zhang X H. Current status, problem and strategy of maize breeding in Shananxi Province., 2008, 16(2): 139–141 (in Chinese with English abstract).
[16] Gan Y T, Siddique K H M, Turner N C, Li X G, Niu J Y, Yang C, Liu L P, Chai Q. Ridge-furrow mulching systems-an innovative technique for boosting crop productivity in semiarid rain-fed environments., 2013, 118: 429–476.
[17] Hou P, Cui Z L, Bu L D, Yang H S, Zhang F S, Li S K. Evaluation of a modified hybrid-maize model incorporating a newly developed module of plastic film mulching., 2014, 54: 2796–2804.
[18] Wang Y J, Xie Z K, Malhi S S, Vera C L, Zhang Y B, Guo Z H. Effects of gravel-sand mulch, plastic mulch and ridge and furrow rainfall harvesting system combinations on water use efficiency, soil temperature and watermelon yield in a semi-arid Loess Plateau of northwestern China., 2011, 101: 88–92.
[19] Mo F, Wang J Y, Zhou H, Luo C L, Zhang X F, Li X Y, Li F M, Xiong L B, Kavagi L, Nguluu S N, Xiong Y C. Ridge-furrow plastic-mulching with balanced fertilization in rainfed maize (L.): an adaptive management in east African Plateau., 2017, 236: 100–112.
[20] Mckenzie C L, Lapointe S L, Duncan L W. Landscape fabric as a physical barrier to neonate(Coleoptera: Curculionidae)., 2001, 84: 721–722.
[21] Wang C B, Wang H, Zhao X M, Chen B H, Wang F L. Mulching affects photosynthetic and chlorophyll a fluorescence characteristics during stage III of peach fruit growth on the no mulching semiarid Loess Plateau of China.(Amsterdam), 2015, 194: 246–254.
[22] Hou F Y, Zhang L M, Xie B T, Dong S X, Zhang H Y, Li A X, Wang Q M. Effect of plastic mulching on the photosynthetic capacity, endogenous hormones and root yield of summer-sown sweet potato ((L.) Lam.) in Northern China., 2015, 37: 1–10.
[23] Ali S, Xu Y Y, Jia Q M, Ahmad I, Ma X C, Henchiri M, Ren X L, Zhang P, Cai T, Zhang J H, Jia Z K. Ridge-furrow mulched with plastic film improves the anti-oxidative defence system and photosynthesis in leaves of winter wheat under deficit irrigation., 2018, 13: e0200277.
[24] Ali S, Xu Y Y, Jia Q M, Ma X C, Ahmad I, Adnan M, Gerard R, Ren X L, Zhang P, Cai T, Zhang J H, Jia Z K. Interactive effects of plastic film mulching with supplemental irrigation on winter wheat photosynthesis, chlorophyll fluorescence and yield under simulated precipitation conditions., 2018, 207: 1–14.
[25] Riedelsheimer C, Lisec J, Czedik-Eysenberg A, Sulpice R, Flis A, Grieder C, Altmann T, Stitt M, Willmitzer L, Melchinger A E. Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize., 2012, 109: 8872–8877.
[26] Witt S, Galicia L, Lisec J, Cairns J, Tiessen A, Araus J L, Palacios-Rojas N, Fernie A R. Metabolic and phenotypic responses of greenhouse-grown maize hybrids to experimentally controlled drought stress., 2012, 5: 401–417.
[27] Degenkolbe T, Do P T, Kopka J, Zuther E, Hincha D K, K?hl K I. Identification of drought tolerance markers in a diverse population of rice cultivars by expression and metabolite profiling., 2013, 8: e63637.
[28] Hill C B, Taylor J D, Edwards J, Mather D, Langridge P, Bacic A, Roessner U. Detection of QTL for metabolic and agronomic traits in wheat with adjustments for variation at genetic loci that affect plant phenology., 2015, 233: 143–154.
[29] Jones R J, Simmon S R. Effect of altered source-sink ratio on growth of maize kernels., 1983, 23: 129–134.
[30] Tollenaar M, Lee E A. Dissection of physiological processes underlying grain yield in maize by examining genetic improvement and heterosis., 2006, 51: 399–408.
[31] Osakabe Y, Yamaguchi-Shinozaki K, Shinozaki K, Tran L P. ABA control of plant macroelement membrane transport systems in response to water deficit and high salinity., 2014, 202: 35–49.
[32] Niu L, Yan Y, Hou P, Bai W, Zhao R, Wang Y, Li S, Du T, Zhao M, Song J, Zhou W. Influence of plastic film mulching and planting density on yield, leaf anatomy, and root characteristics of maize on the Loess Plateau., 2020, 8: 548–564.
[33] Ren B Z, Liu W, Zhang J W, Dong S T, Liu P, Zhao B. Effects of plant density on the photosynthetic and chloroplast characteristics of maize under high-yielding conditions., 2017, 104: 12.
[34] 明博, 謝瑞芝, 侯鵬, 李璐璐, 王克如, 李少昆. 2005–2016年中國玉米種植密度變化分析. 中國農業科學, 2017, 50: 1960–1972. Ming B, Xie R Z, Hou P, Li L L, Wang K R, Li S K. Changes of maize planting density in China. 2017, 50: 1960–1972 (in Chinese with English abstract).
[35] Glaubitz U, Erban A, Kopka J, Hincha D K, Zuther E. High night temperature strongly impacts TCA cycle, amino acid and polyamine biosynthetic pathways in rice in a sensitivity-dependent manner., 2015, 66: 6385–6397.
[36] Kopka J, Schauer N, Krueger S, Birkemeyer C, Usadel B, Bergmuller E, Dormann P, Weckwerth W, Gibon Y, Stitt M, Willmitzer L, Fernie A R, Steinhauser D. GMD@CSB.DB: the golm metabolome database., 2005, 21: 1635–1638.
[37] Hummel J, Strehmel N, Selbig J, Walther D, Kopka J. Decision tree supported substructure prediction of metabolites from GC-MS profiles., 2010, 6: 322–333.
[38] Jannink J L, Lorenz A J, Iwata H. Genomic selection in plant breeding: from theory to practice., 2010, 9: 166–177.
[39] Obata T, Fernie A R. The use of metabolomics to dissect plant responses to abiotic stresses., 2012, 69: 3225–3243.
[40] Stewart G R, Larher F. Accumulation of Amino Acids and Related Compounds in Relation to Environmental Stress. In: Miflin B J, eds. Amino Acids and Derivatives. New York: Academic Press, 2001. pp 609–635.
[41] Good A G, Zaplachinski S T. The effects of drought stress on free amino acid accumulation and protein synthesis in., 1994, 90: 9–14.
[42] Huang T F, Jander G. Abscisic acid-regulated protein degradation causes osmotic stress-induced accumulation of branched-chain amino acids in., 2017, 246: 737–747.
[43] Rai V K. Role of amino acids in plant responses to stresses., 2002, 45: 481–487.
[44] Bauwe H, Hagemann M, Kern R, Timm S. Photorespiration has a dual origin and manifold links to central metabolism., 2012, 15: 269–275.
[45] 秦嶺, 陳二影, 張艷亭, 楊延兵, 孔清華, 張華文, 王海蓮, 王潤豐, 管延安, 阮穎. 干旱及復水對谷子脯氨酸積累和基因表達的影響. 分子植物育種, 2018, 16: 7460–7465. Qin L, Chen E Y, Zhang Y T, Yang Y B, Kong Q H, Zhang H W, Wang H L, Wang R F, Guan Y A, Ruan Y. Effects of drought and rehydration on proline accumulation andgene expression from foxtail millet ((L.) Beauv)., 2018, 16: 7460–7465 (in Chinese with English abstract).
[46] Filippou P, Antoniou C, Fotopoulos V. The nitric oxide donor sodium nitroprusside regulates polyamine and proline metabolism in leaves ofplants., 2013, 56: 172–183.
[47] Joshi V, Joung J, Fei Z J, Jander G. Interdependence of threonine, methionine and isoleucine metabolism in plants: accumulation and transcriptional regulation under abiotic stress., 2010, 39: 933–947.
[48] Araújo W L, Tohge T, Ishizaki K, Leaver C J, Fernie A R. Protein degradationan: an alternative respiratory substrate for stressed plants., 2011, 16: 489–498.
[49] Fait A, Fromm H, Walter D, Galili G, Fernie A R. Highway or byway: the metabolic role of the GABA shunt in plants., 2008, 13: 14–19.
[50] Yadav A K, Carroll A J, Estavillo G M, Rebetzke G J, Pogson B J. Wheat drought tolerance in the field is predicted by amino acid responses to glasshouse-imposed drought., 2019, 70: 4931–4948.
[51] Ravanel S, Gakiere B, Job D, Douce R. The specific features of methionine biosynthesis and metabolism in plants., 1998, 95: 7805–7812.
[52] Bryan J K. Differential regulation of maize homoserine dehydrogenase under physiological conditions., 1990, 92: 785–791.
[53] Lamari N, Zhendre V, Urrutia M, Bernillon S, Maucourt M, Deborde C, Prodhomme D, Jacob D, Ballias P, Rolin D, Sellier H, Rabier D, Gibon Y, Giauffret C, Moing A. Metabotyping of 30 maize hybrids under early-sowing conditions reveals potential marker-metabolites for breeding., 2018, 14: 132.
[54] Obata T, Witt S, Lisec J, Palacios-Rojas N, Florez-Sarasa I, Araus J L, Cairns J E, Yousfi S, Fernie A R. Metabolite profiles of maize leaves in drought, heat and combined stress field trials reveal the relationship between metabolism and grain yield., 2015, 169: 2665–2683.
[55] Mahal H S, Mukherjee T. Scavenging of reactive oxygen radicals by resveratrol: antioxidant effect., 2006, 32: 59–71.
[56] Islam M T, Ali E S, Uddin S J, Shaw S, Islam M A, Ahmed M I, Chandra Shill M, Karmakar U K, Yarla N S, Khan I N, Billah M M, Pieczynska M D, Zengin G, Malainer C, Nicoletti F, Gulei D, Berindan-Neagoe I, Apostolov A, Banach M, Yeung A W K, El-Demerdash A, Xiao J, Dey P, Yele S, Jó?wik A, Strza?kowska N, Marchewka J, Rengasamy K R R, Horbańczuk J, Kamal M A, Mubarak M S, Mishra S K, Shilpi J A, Atanasov A G. Phytol: a review of biomedical activities., 2018, 121: 82–94.
[57] Eastmond P J. Glycerol-insensitivemutants:seedlings lack glycerol kinase, accumulate glycerol and are more resistant to abiotic stress., 2004, 37: 617–625.
Effects of plastic film mulching on leaf metabolic profiles of maize in the Loess Plateau with two planting densities
NIU Li1,2,3, BAI Wen-Bo1, LI Xia2, DUAN Feng-Ying2, HOU Peng2, ZHAO Ru-Lang4, WANG Yong-Hong4, ZHAO Ming2, LI Shao-Kun2, SONG Ji-Qing1,*, and ZHOU Wen-Bin2,*
1Key Laboratory of Prevention and Control of Residual Pollution in Agricultural Film, Ministry of Agriculture and Rural Affairs / Institute of Environment and Sustainable Development in Agriculture, Chinese Academy of Agricultural Sciences, Beijing 100081, China;2Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China;3College of Water Resources and Civil Engineering, China Agricultural University, Beijing 100083, China;4Ningxia Academy of Agriculture and Forestry Sciences, Crops Research Institute, Yongning 750105, Ningxia, China
In the Loess Plateau, the study of leaf metabolomics under plastic mulching is an important step to explore the mechanism of plastic film mulching to increase grain yield in maize. Metabolomics analysis was performed on the leaves of two maize cultivars (Zhengdan 958 and Xianyu 335) under two planting densities (7.5×104and 10.5×104plant hm–2) and two mulching modes (plastic film mulching and no mulching ) at silking stage using gas chromatography-quadrupole-time of flight mass spectrometry (GC-QTOF) technology. In term of the number of differential metabolites, the response of Xianyu 335 to plastic film mulching was greater than that of Zhengdan 958. The higher planting density reduced the difference in leaf metabolism between plastic film mulching and no mulching. The principal component analysis revealed that both the mulching treatment and the cultivar had significant impacts on the composition of the metabolites. The separation of metabolic spectrum along plastic film mulching was mainly driven by organic acids such as citric acid and amino acids; the separation along cultivar was mainly driven by alkanoic acids and phenols. Correlation analysis indicated that resveratrol, phytol and glucose-6-phosphate had significant positive correlations with maize grain yield, whereas glycerol had a significant negative correlation with it. Under plastic film mulching condition, the levels of valine, isoleucine and methionine related to respiration and elimination of photorespiration products; isocitrate related to tricarboxylic acid cycle, methionine, N-acetylaspartic acid and other metabolites that could reduce photoinhibition showed an overall increasing trend. These results indicated that, under plastic film mulching condition, the antioxidant and energy metabolism-related metabolites played important roles in increasing grain yield. The elimination of photorespiration products and the accumulation of metabolites to alleviate photoinhibition were the metabolic bases for the increase of net photosynthetic rate in maize leaves.
plastic film mulching; leaf; metabolome; yield; antioxidant; maize
10.3724/SP.J.1006.2021.03053
本研究由國家重點研發計劃項目(2016YFD0300102)資助。
This study was supported by the National Key Research and Development Program of China (2016YFD0300102).
周文彬, E-mail: zhouwenbin@caas.cn, Tel: 010-82108609; 宋吉青, E-mail: songjiqing@caas.cn
E-mail: niulipipi@163.com
2020-09-11;
2020-12-01;
2021-01-04.
URL: https://kns.cnki.net/kcms/detail/11.1809.S.20210104.1347.008.html
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農業(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
聚氯乙烯(2018年9期)2018-02-18 01:11:34
