機插密度和穗肥減量對優質食味水稻品種籽粒淀粉合成的影響
2021-06-09 13:16:16張宏路李思宇張亞軍韋佳利顧駿飛劉立軍楊建昌
作物學報 2021年8期
關鍵詞:水稻
陳 云 劉 昆 張宏路 李思宇 張亞軍 韋佳利 張 耗 顧駿飛 劉立軍,* 楊建昌
機插密度和穗肥減量對優質食味水稻品種籽粒淀粉合成的影響
陳 云1,2劉 昆1張宏路1李思宇1張亞軍2韋佳利2張 耗1顧駿飛1劉立軍1,*楊建昌1
1揚州大學江蘇省作物遺傳生理重點實驗室/ 江蘇省糧食作物現代產業技術協同創新中心 / 江蘇省作物基因組學和分子育種重點實驗室, 江蘇揚州 225009;2揚州大學生物科學與技術學院, 江蘇揚州 225009
機插密度對優質食味水稻品種籽粒淀粉合成的影響尚不明確。試驗以3個江蘇省代表性優質食味水稻品種南粳9108、南粳5055和南粳46為材料, 行距固定為30 cm, 通過設置10、12、14、16和18 cm株距, 形成33.3×104、27.8×104、23.8×104、20.8×104和18.5×104穴 hm–25種機插密度, 研究了機插密度對上述優質食味水稻品種產量和籽粒淀粉合成的影響, 并觀察了穗肥施氮量減半施用對優質食味水稻籽粒淀粉合成的調控效應。結果表明: (1) 3個優質食味水稻品種產量均在密度為27.8×104穴 hm–2時最高。成熟期稻米直鏈淀粉含量隨機插密度增加呈先下降后上升的趨勢, 支鏈淀粉含量則不斷下降。機插密度控制在27.5×104~29.9×104穴 hm–2時, 有利于提高上述優質食味水稻品種的食味值。(2) 隨機插密度增加, 顆粒結合態淀粉合成酶(granule-bound starch synthase, GBSS)和ADP-葡萄糖焦磷酸化酶(ADP-glucose pyrophosphorylase, AGP)在籽粒灌漿前中期(抽穗后8~20 d)的活性有所上升, 但在灌漿后期迅速下降, 而機插密度對淀粉分支酶(starch branching enzyme, SBE)活性無明顯影響。其中, 抽穗后12~16 d籽粒中GBSS和AGP活性與直鏈淀粉和總淀粉含量呈顯著或極顯著正相關。(3) 與正常穗肥相比, 穗肥減半處理能顯著降低優質食味水稻品種灌漿前中期籽粒中GBSS和AGP活性, 降低稻米直鏈淀粉含量, 提升優質食味稻米的食味值。上述結果表明, 機插密度可通過調控灌漿前中期籽粒中淀粉合成關鍵酶(GBSS和AGP)活性, 調控直鏈淀粉含量進而影響稻米食味值。合理的機插密度和穗肥施用量有助于進一步改善優質食味稻米的食味品質。
優質食味水稻; 機插密度; 淀粉合成關鍵酶; 食味值; 直鏈淀粉
中國是世界上最大的水稻生產國和消費[1-2]。近年來, 隨著社會經濟的不斷發展和產業結構的調整, 農村大量青壯年勞動力流失, 從事農業生產勞動力嚴重不足[3]。水稻生產機械化是實現水稻高效種植的重要途徑, 推進以機插秧為主的水稻機械化高產種植技術, 對穩定我國水稻生產、保障糧食安全具有重要意義[4]。機插密度影響水稻群體結構、產量形成和稻米品質, 確定合理的種植密度是高產優質栽培基本的因素[5]。由于粳稻多為中、小穗型, 產量往往受到基本苗數量不足的限制, 適當密植(23.8×104~ 27.8×104穴 hm-2)可有效提高粳稻產量[5-6]。
近年來, 隨著我國經濟的發展和人民生活水平的提高, 人們對稻米的需求逐漸從“吃得飽”向“吃得好”轉變, 迫切需要在穩定產量的同時提高稻米品質, 尤其是食味品質, 以滿足供給側結構性改革的需求[3]。近年來, 江蘇省農業科學院選育的南粳系列優良食味粳稻品種, 因優良的食味品質獲得廣大消費者的好評。而北方的“面食改米食”和南方的“秈米改粳米”趨勢明顯, 更加大了稻米市場對粳米, 特別是優質食味粳稻的需求量。但機插密度對優質食味水稻蒸煮食味品質的影響尚不清楚。
淀粉是稻米的主要組成部分, 是由葡萄糖多聚體構成的高度有序的晶狀質體, 是人類重要的熱量來源, 對稻米的理化性質和蒸煮性能起著基礎性作用[7-8]。根據淀粉結構不同, 分為直鏈淀粉和支鏈淀粉[9-10]。直鏈淀粉的分子量較小(105~106Da)、分支極少, 而支鏈淀粉分子分子量較大(107~109Da)、具有高度分支性[11]。直鏈淀粉的生物合成主要由ADP-葡萄糖焦磷酸酶(ADP-glucose pyrophosphorylase, AGP)和顆粒結合淀粉合成酶(granule-bound starch synthase, GBSS)控制, 而支鏈淀粉的反應則更為復雜, 需要在AGP、可溶性淀粉合成酶和淀粉分支酶(starch branching enzyme, SBE)等一系列酶的作用下生成。直鏈淀粉含量一直是衡量稻米蒸煮食味品質的關鍵指標, 一般認為食味優的水稻品種具有較低的直鏈淀粉含量[12-13]。直鏈淀粉含量不僅對米飯食味品質和質地起重要作用, 也與眾多的淀粉理化參數如黏滯性、糊化特性、回生特性密切相關[7]。直鏈淀粉含量除受主效基因位點控制外, 還受灌漿結實期溫光資源以及栽培措施的影響[7,14-16]。施用氮素穗肥除了會對水稻產量產生影響外, 亦會影響水稻籽粒淀粉結構和理化性質, 從而影響稻米的食味品質[17]。目前關于機插密度和氮素穗肥對優質食味水稻品種淀粉合成, 尤其是直鏈淀粉合成的影響研究較少, 其生理機制尚不明確。
本研究選用3個食味品質優良的水稻品種為材料, 研究了機插密度對其籽粒淀粉合成的影響及穗肥減量對優質食味水稻籽粒淀粉合成的調控。旨在進一步揭示機插密度影響優質食味水稻品種籽粒淀粉合成的生理機制, 為優質食味水稻品種選育以及栽培調控提供理論與實踐依據。
1 材料與方法
1.1 供試材料與試驗設計
以江蘇大面積推廣應用的優質食味水稻品種南粳9108、南粳5055和南粳46為試驗材料。上述水稻品種在生產中常用的機插密度通常為22.5×104~ 27×104穴 hm-2(國家水稻數據中心, http://www. ricedata.cn), 全生育期施氮量(折合純氮) 300 kg hm-2左右[18]。根據研究需要, 設置以下2組試驗。
試驗1: 機插密度試驗。采用常規塑盤毯苗育秧, 5月20日至21日播種, 播種量為每盤干谷120 g, 6月13日至14日采用洋馬VP6乘坐式水稻插秧機進行機械移栽, 每穴栽插3~4株苗。行距固定為30 cm, 通過設置10、12、14、16和18 cm株距, 形成33.3×104、27.8×104、23.8×104、20.8×104和18.5×104穴 hm–2等5種機插密度。小區面積400 m2, 隨機區組排列, 重復3次。全生育期施氮肥(折合純氮) 300 kg hm–2, 所施氮肥為尿素, 按基肥∶分蘗肥∶穗肥=4∶3∶3施用, 穗肥分促花肥、保花肥2次等量施用。基施過磷酸鈣(含P2O513.5%) 120 kg hm–2和氯化鉀(含K2O 62.5%) 120 kg hm–2。試驗重復2年(2017年和2018年)。
試驗2: 穗肥施氮量減半試驗。試驗于2019年5月22日播種, 6月16日機械移栽。試驗采用裂區設計, 品種為主區, 穗肥施氮量處理為裂區。行株距固定為30 cm ×12 cm (對應機插密度為27.8×104穴 hm-2), 設置2種穗肥施氮量處理: (1) 正常穗肥用量(normal N rate of panicle nitrogen fertilizer, NR):穗肥施氮量與試驗1相同, 為90 kg hm–2; (2) 穗肥用量減半(half normal N rate of panicle nitrogen fertilizer, HR): 穗肥施氮量為45 kg hm–2。穗肥分促花肥、保花肥2次等量施用。小區面積150 m2, 重復3次。供試材料、育秧和基蘗肥用量及施用方法等同試驗1。各小區作田埂覆膜分開, 單獨排灌, 以防水肥串灌。
以上2組試驗, 均在揚州大學江蘇省作物栽培生理重點實驗室校外試驗農場鎮江市新民洲農場(32°16′N, 119°33′E)進行。試驗地前茬作物均為小麥, 土壤類型為沙壤土, 耕作層有機質、有效氮、速效磷和速效鉀含量分別為22.4 g kg–1、106.2 mg kg–1、32.4 mg kg–1和87.5 mg kg–1。水分管理采用“淺—擱—濕”灌溉方式, 即薄水移栽, 活棵立苗后采取淺水勤灌促分蘗, 分蘗末期斷水擱田, 拔節后采取干濕交替灌溉, 抽穗至成熟前7 d濕潤灌溉, 之后斷水。全生育期嚴格控制病蟲草害。
1.2 取樣與測定
1.2.1 籽粒淀粉代謝相關酶活性 在抽穗期, 每品種每處理選擇生長狀況基本一致的抽穗單莖300個掛牌并標記日期。分別在抽穗后8、12、16、20、24、28和32 d, 取掛牌單莖20個, 摘下稻穗中部籽粒于液氮中冷凍30s, 置于-80℃冰箱保存待測。在測定酶活性時, 先將籽粒剝殼, 使用德國萊馳RETSCH MM400混合球磨儀充分研磨, 參照Yang等[19]方法測定顆粒結合態淀粉合成酶(GBSS)、ADP-葡萄糖焦磷酸化酶(AGP)以及淀粉分支酶(SBE)活性。
1.2.2 籽粒直鏈淀粉、總淀粉積累量 在上述相同時期和成熟期, 取掛牌單莖稻穗中部籽粒, 測定籽粒直鏈淀粉和總淀粉積累量的變化動態。
1.2.3 考種與計產 在成熟期, 各小區調查100株穗數, 并按平均穗數取樣10穴, 手工脫粒, 考查水稻每穗粒數、結實率和千粒重等產量構成因素。結實率采用水漂法測定。各小區實收5 m2, 機器脫粒曬干后測定重量和含水率, 按13.5%的含水率折算為實收產量。
1.2.4 稻米淀粉含量 取存放3個月后的實收稻谷樣本, 參照中華人民共和國國家標準《GB/T 17891-1999優質稻谷》測定稻米直鏈淀粉含量。總淀粉含量測定參照Zhu等[20]方法, 并以總淀粉含量與直鏈淀粉含量的差值計算支鏈淀粉含量。
1.2.5 米飯食味特性 稱取30 g精米, 放入不銹鋼罐中并加水浸泡30 min后接洗米裝置, 用水沖洗約30 s至水的濁度很小。按照米水1.0∶1.3的比例加水, 放入電飯鍋內預約1 h, 調理30 min, 燜飯10 min。冷卻2 h后采用日本佐竹公司生產的STA1B型米飯食味計(STA1B, SATAKE Co., Ltd, 日本)測定米飯食味值, 每個樣品重復3次。
1.3 數據計算與統計分析
用Microsoft Excel 2003與SPSS軟件統計分析數據, SigmaPlot 11.0和R繪圖。
2 結果與分析
2.1 方差分析
方差分析(表1)表明, 機插密度對南粳9108、南粳5055和南粳46三個優質食味水稻品種產量、整精米率、堊白粒率、直鏈淀粉含量、蛋白質含量和食味值的影響在密度和品種間存在差異顯著, 而上述指標在年份、年份×密度、年份×品種、密度×品種和年份×密度×品種間差異均不顯著。因此, 本文中試驗1重點以2018年的數據進行分析。
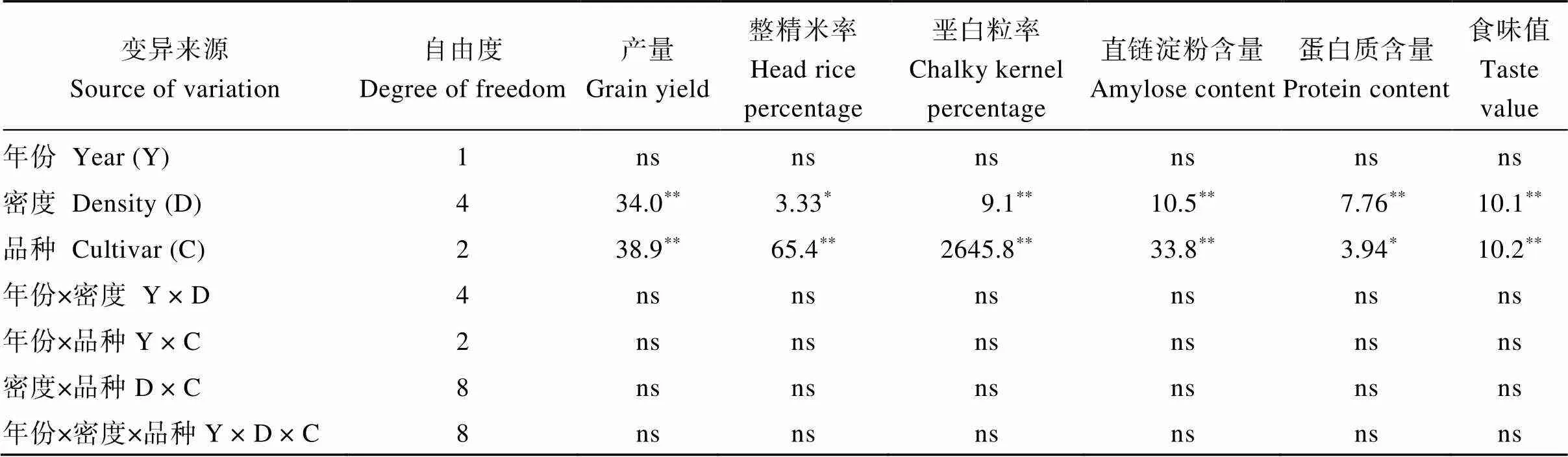
表1 機插密度對優質食味水稻品種產量和稻米主要品質指標的方差分析
*、**分別表示在0.05和0.01水平上顯著, ns表示差異不顯著(> 0.05)。
*and**mean significant differences at the 0.05 and 0.01 probability levels, respectively. ns: not significant at> 0.05.
2.2 機插密度對優質食味水稻品種產量及其構成因素的影響
機插密度對優質食味水稻品種南粳9108、南粳5055和南粳46產量及產量構成因素的影響趨勢大致相同(圖1)。隨機插密度的增加, 3個優質食味水稻品種產量, 均表現為先上升后下降的趨勢, 且均在密度為27.8×104穴 hm–2時產量最高。單位面積穗數、結實率總體隨機插密度增大呈先升后降, 而每穗粒數和千粒重則逐漸降低。
2.3 機插密度對優質食味水稻品種淀粉含量和食味值的影響
3個優質食味水稻品種籽粒直鏈淀粉含量隨機插密度的增加呈先降后升的趨勢, 支鏈淀粉、總淀粉含量基本隨機插密度的增加而降低。稻米食味值隨機插密度的增加呈先升后降的趨勢, 供試3個品種的表現一致。與南粳9108、南粳46相比, 南粳5055在相同機插密度條件下具有較低的直鏈淀粉含量與較高的食味值(圖2)。相關分析表明, 在不同機插密度下3個優質食味水稻品種的食味值與直鏈淀粉和總淀粉含量呈極顯著負相關, 但與支鏈淀粉含量之間的相關性不顯著(圖3)。對機插密度與直鏈淀粉含量進行方程擬合可以看出, 機插密度為27.5′104~29.9×104穴 hm–2時, 3個優質食味水稻品種直鏈淀粉含量最低, 食味值較高(表2)。
2.4 機插密度對優質食味水稻籽粒直鏈淀粉和總淀粉積累量的影響
不同機插密度條件下, 3個優質食味水稻品種籽粒直鏈淀粉和總淀粉積累量, 在抽穗后32 d即達最終積累量的69.7%~95.1%和80.3%~92.4%, 此后至成熟變化相對較小。隨機插密度增加, 水稻籽粒直鏈淀粉積累量呈先降后升的變化趨勢, 而總淀粉積累量則隨機插密度增加而逐漸降低(圖4)。
18.5、20.8、23.8、27.8和33.3分別表示機插密度為18.5′104、20.8′104、23.8′104、27.8′104和33.3′104穴 hm–2。
18.5, 20.8, 23.8, 27.8, and 33.3 represent the transplanting density of 18.5 × 104, 20.8 × 104, 23.8 × 104, 27.8 × 104, and 33.3 × 104hills hm–2, respectively.
*,< 0.05; **,< 0.01.
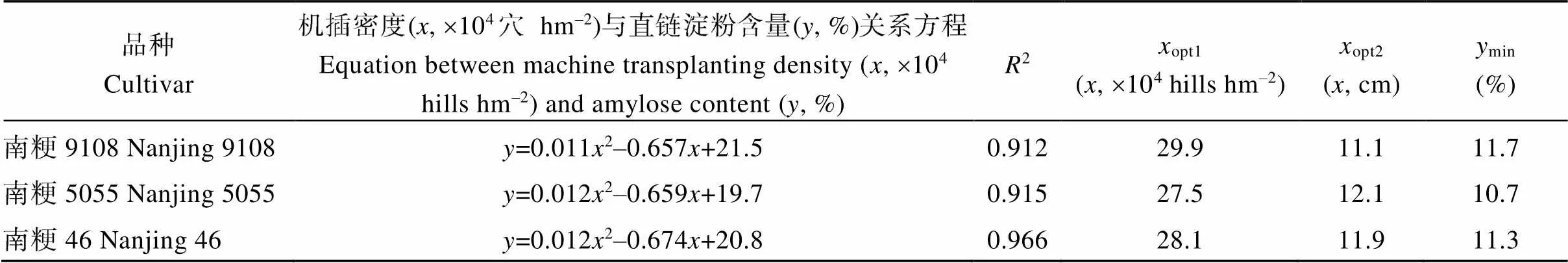
表2 優質食味水稻品種獲得最低直鏈淀粉含量時的機插密度(2018年)
opt1: 獲得最低直鏈淀粉含量時的機插密度;opt2: 獲得最低直鏈淀粉含量時的機插株距;min: 最低直鏈淀粉含量。
opt1: the optimal machine transplanting density for minimum amylose content;opt2: the optimal plant spacing for minimum amylose content;min: minimum amylose content.
18.5、20.8、23.8、27.8和33.3分別表示機插密度為18.5′104、20.8′104、23.8′104、27.8′104和33.3′104穴 hm–2。
18.5, 20.8, 23.8, 27.8, and 33.3 represent the transplanting density of 18.5′104, 20.8′104, 23.8′104, 27.8′104, and 33.3′104hills hm–2, respectively.
2.5 機插密度對優質食味水稻籽粒淀粉合成關鍵酶活性的影響
在不同機插密度下, 3個優質食味水稻品種灌漿籽粒中的淀粉合成關鍵酶(GBSS、SBE和AGP)活性皆呈先上升后下降的趨勢, 且基本都在抽穗后20 d達到峰值。機插密度對3個優質食味水稻品種SBE活性的影響較小(圖5), 而對GBSS和AGP活性的影響則表現為: 抽穗前中期(抽穗后0~20 d)機插密度越大, 酶活性越低, 后期則相反(圖6和圖7)。相關分析表明(表3), 稻米直鏈淀粉含量與抽穗后12~16 d籽粒中GBSS和AGP活性呈顯著或極顯著正相關(= 0.523*~0.865**), 而總淀粉含量與抽穗后12~20 d籽粒中上述2種酶活性均呈極顯著正相關(= 0.659**~0.771**)。
18.5、20.8、23.8、27.8和33.3分別表示機插密度為18.5′104、20.8′104、23.8′104、27.8′104和33.3′104穴 hm–2。
18.5, 20.8, 23.8, 27.8, and 33.3 represent the transplanting density of 18.5′104, 20.8′104, 23.8′104, 27.8′104, and 33.3′104hills hm–2, respectively.
18.5、20.8、23.8、27.8和33.3分別表示機插密度為18.5′104、20.8′104、23.8′104、27.8′104和33.3′104穴 hm–2。
18.5, 20.8, 23.8, 27.8, and 33.3 represent the transplanting density of 18.5′104, 20.8′104, 23.8′104, 27.8′104, and 33.3′104hills hm–2, respectively.
18.5、20.8、23.8、27.8和33.3分別表示機插密度為18.5′104、20.8′104、23.8′104、27.8′104和33.3′104穴 hm–2。
18.5, 20.8, 23.8, 27.8, and 33.3 represent the transplanting density of 18.5′104, 20.8′104, 23.8′104, 27.8′104, and 33.3′104hills hm–2, respectively.
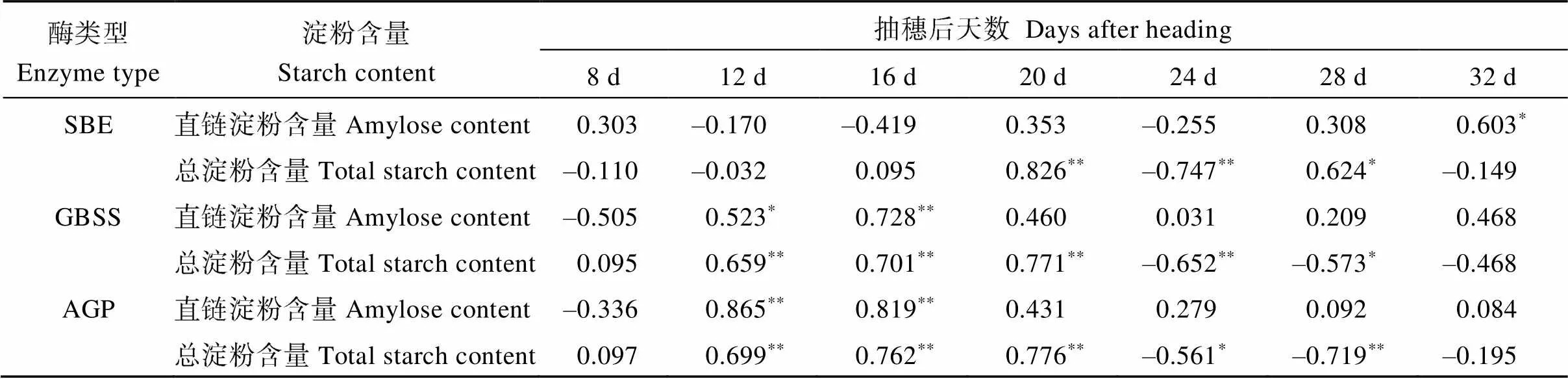
表3 優質食味水稻抽穗后不同時期籽粒中淀粉合成關鍵酶活性與成熟期稻米直鏈淀粉、總淀粉含量的相關(2018年)
*和**分別表示在0.05和0.01水平上顯著。SBE: 淀粉分支酶; GBSS: 顆粒結合態淀粉合成酶; AGP: ADP-葡萄糖焦磷酸化酶。
*and**mean significant difference at the 0.05 and 0.01 probability levels, respectively. SBE: starch branching enzyme; GBSS: granule-bound starch synthase; AGP: ADP-glucose pyrophosphorylase.
2.6 穗肥減半對優質食味水稻產量和籽粒淀粉的調控
與正常穗肥用量(NR)相比, 穗肥用量減半處理(HR)使南粳9108、南粳5055和南粳46的產量略有降低, 但均未達顯著水平。HR顯著降低了3個品種的直鏈淀粉含量(降幅達5.5%~8.7%), 提高了其食味值(增幅達4.9%~7.2%)。HR對支鏈淀粉和總淀粉含量影響較小(表4)。與NR相比, HR對抽穗后12~16 d籽粒中SBE活性無顯著影響, 但顯著降低了GBSS和AGP活性(圖8)。
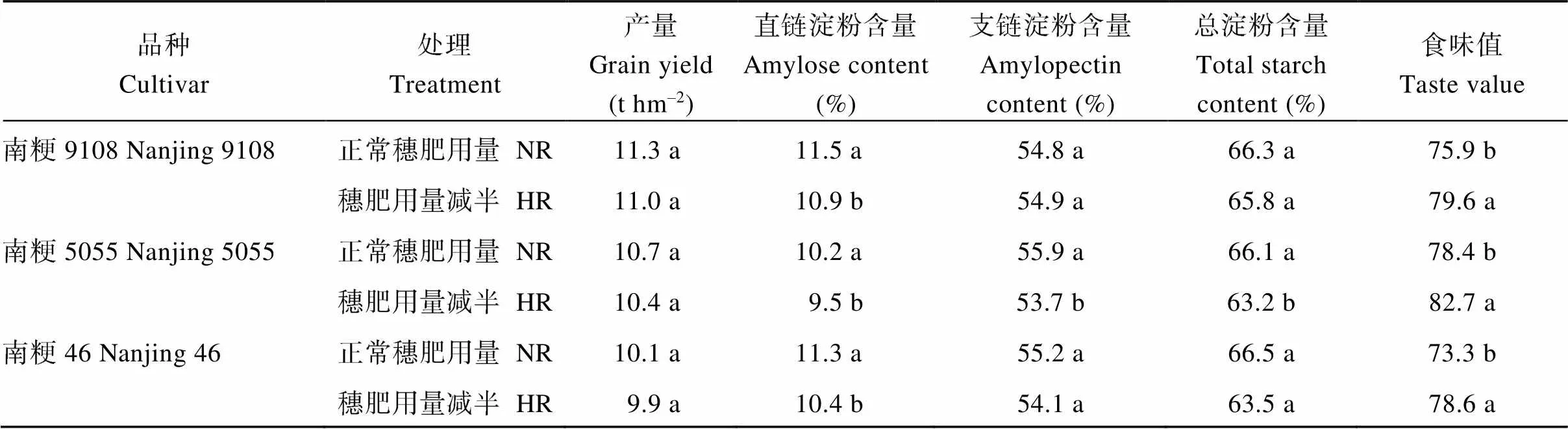
表4 穗肥減半對優質食味水稻品種產量、直鏈淀粉含量、總淀粉含量和食味值的影響(2019年)
NR: 正常穗肥用量; HR: 穗肥用量減半。標以不同字母表示在同一品種內比較0.05水平上差異顯著。
NR: normal N rate of panicle nitrogen fertilizer; HR: half normal N rate of panicle nitrogen fertilizer. Different lowercase letters within the same column indicate statistical significance at the= 0.05 level within the same cultivar.
NR: 正常穗肥用量; HR: 穗肥用量減半。SBE: 淀粉分支酶; GBSS: 顆粒結合態淀粉合成酶; AGP: ADP-葡萄糖焦磷酸化酶。酶活性以抽穗后12 d和16 d兩次測定的平均值表示。
NR: normal N rate of panicle nitrogen fertilizer; HR: half normal N rate of panicle nitrogen fertilizer. SBE: starch branching enzyme; GBSS: granule-bound starch synthase; AGP: ADP-glucose pyrophosphorylase. The activities of enzymes were means of two repetitions at 12 and 16 days after heading.
3 討論
水稻生產機械化是實現水稻高效種植的重要途徑, 機插密度是影響水稻群體結構、產量形成和稻米品質的重要調控因素, 確定合理的種植密度是水稻高產優質栽培最基本、最關鍵的因素[5]。淀粉是水稻胚乳的主要成分, 是種子萌發和幼苗生長過程中碳水化合物的主要來源。直鏈淀粉含量一直是衡量稻米蒸煮食味品質的關鍵指標, 一般認為食味優的水稻品種具有較低的直鏈淀粉含量[12-13]。直鏈淀粉含量降低是中國秈稻食味品質得以改善的一個主要原因[21]。在本試驗條件下, 隨機插密度的增加, 3個優質食味水稻品種(南粳9108、南粳5055和南粳46)籽粒直鏈淀粉含量呈先降后升的趨勢, 而食味值與之相反。食味值與直鏈淀粉含量呈極顯著負相關關系, 即降低直鏈淀粉含量有助于提升稻米的食味值。以往對何種機插密度利于降低水稻直鏈淀粉含量, 進而提高稻米食味值研究結果不一。胡雅杰等[22]研究表明, 適當降低機插密度(19.5×104~25.5×104穴 hm–2時)能夠降低不同穗型水稻品種蛋白質和直鏈淀粉含量, 改善淀粉RVA譜特征。但亦有研究者指出, 增加密度有利于降低粳稻直鏈淀粉含量, 提高稻米食味值[23]。本研究觀察到, 將機插密度控制在27.5×104~29.9×104穴 hm–2時, 3個優質食味水稻品種直鏈淀粉含量最低, 此時食味值較高。
GBSS、SBE和AGP對水稻籽粒淀粉合成具有關鍵作用, GBSS和AGP不僅參與水稻籽粒灌漿, 還與直鏈淀粉含量密切相關[24-25], AGP還被認為是淀粉合成相關途徑的限速酶[26-27], 而SBE主要參與促進形成支鏈淀粉[28]。以往對玉米、小麥大田作物籽粒淀粉代謝相關途徑的研究較多[29-30], 而對水稻尤其是優質食味稻種相關代謝途徑調控的研究鮮有涉及[17], 或多以溫度[29]、水肥[17,31]等調控措施為主。已有研究表明, 灌漿期高溫可提高非優質食味水稻品種淀粉的糊化溫度, 降低直鏈淀粉含量, 增加支鏈淀粉中相對較長的葡聚糖鏈的比例[32]。非優質食味水稻品種直鏈淀粉含量降低的原因在于GBSS酶的高表達[33-34]。但亦有相反的觀點認為, 非優質食味水稻品種淀粉代謝關鍵酶(GBSS、SBE和AGP)活性降低能夠減少直鏈淀粉含量, 進而提高稻米食味值[35]。因此, 非優質食味水稻品種籽粒上述淀粉代謝關鍵酶活性與直鏈淀粉含量的關系仍需進一步深入研究。可溶性淀粉合成酶(soluble starch synthase, SSS)可通過催化支鏈淀粉α-1,4糖苷鍵的形成, 參與支鏈淀粉的生物合成[36-37]。SSS基因發生突變或轉移反SSS基因, 其結構會發生改變、酶活性下降, 支鏈淀粉含量也會相應降低[36]。但本試驗觀察到, 稻米支鏈淀粉含量與食味值并無明顯相關(= 0.333)。因此, SSS對水稻, 尤其是優質食味水稻品種食味值的調控作用尚待進一步明確。
氮素是提高產量和蛋白質含量的重要養分, 但過量施氮會降低稻米品質, 尤其是蒸煮食味品質[31]。胡群等[38]研究表明, 適當增加穗肥比例雖能有效改善稻米的加工品質和營養品質, 但顯著降低了稻米的食味品質。沈鵬等[39-40]以不同品質類型的粳稻品種為材料研究認為, 隨著施氮量的增加, 籽粒中蛋白質含量逐漸提高, 淀粉譜特性變劣, 味度值下降。在本試驗條件下, 與正常穗肥用量相比, 穗肥用量減半處理使南粳9108、南粳5055和南粳46三個優質食味水稻品種的直鏈淀粉含量降低了5.5%~8.7%, 食味值提高了4.9%~7.2%。這可能是由于減施氮肥降低了葉片和籽粒氮代謝相關酶活性, 抑制了氮素吸收和轉運, 導致籽粒中蛋白質含量和直鏈淀粉含量降低所致[41-42]。與正常穗肥用量相比, 穗肥用量減半處理顯著降低優質食味水稻品種抽穗后12~ 16 d籽粒GBSS和AGP活性, 這也進一步驗證了降低灌漿前中期籽粒中GBSS和AGP活性有利于降低稻米直鏈淀粉含量, 優化稻米食味值。
4 結論
優質食味水稻品種食味值與直鏈淀粉含量顯著負相關。灌漿前中期(抽穗后12~16 d)籽粒中GBSS和AGP活性與成熟期稻米直鏈淀粉含量密切相關。合理的機插密度(27.5×104~29.9×104穴 hm–2)有利于在保證優質食味水稻品種產量較高的前提下, 降低灌漿前中期籽粒中GBSS和AGP的活性, 進而降低直鏈淀粉含量, 進一步改善優質稻米的食味值。
[1] Wang Y, Xue Y, Li J. Towards molecular breeding and improvement of rice in China., 2005, 10: 610–614.
[2] Huang M, Chen J, Cao F, Zou Y. Increased hill density can compensate for yield loss from reduced nitrogen input in machine- transplanted double-cropped rice., 2018, 221: 333–338.
[3] Peng S, Tang Q, Zou Y. Current status and challenges of rice production in China., 2009, 12: 3–8.
[4] Xing Z, Hu Y, Qian H, Cao W, Guo B, Wei H, Xu K, Huo Z, Zhou G, Dai Q, Zhang H. Comparison of yield traits in rice among three mechanized planting methods in a rice-wheat rotation system., 2017, 16: 1451–1466.
[5] Hu Q, Jiang W Q, Qiu S, Xing Z P, Hu Y J, Guo B W, Liu G D, Gao H, Zhang H C, Wei H Y. Effect of wide-narrow row arrangement in mechanical pot-seedling transplanting and plant density on yield formation and grain quality ofrice., 2020, 19: 1197–1214.
[6] Liu Q, Zhou X, Li J, Xin C. Effects of seedling age and cultivation density on agronomic characteristics and grain yield of mechanically transplanted rice., 2017, 7: 1–10.
[7] Zhu D, Zhang H, Guo B, Xu K, Dai Q, Wei C, Zhou G, Huo Z. Physicochemical properties of-hybrid rice starch from Chinese varieties., 2017, 63: 356–363.
[8] Chung H J, Liu Q, Lee L, Wei D. Relationship between the structure, physicochemical properties and in vitro digestibility of rice starches with different amylose contents., 2011, 25: 968–975.
[9] Li H, Prakash S, Nicholson T M, Fitzgerald M A, Gilbert R G. The importance of amylose and amylopectin fine structure for textural properties of cooked rice grains., 2016, 196: 702–711.
[10] Yang Y, Lin G, Yu X, Wu Y, Xiong F. Rice starch accumulation at different endosperm regions and physical properties under nitrogen treatment at panicle initiation stage., 2020, 160: 328–339.
[11] Gilbert R G, Witt T, Hasjim J. What is being learned about starch properties from multiple-level characterization., 2013, 90: 312–325.
[12] Hori K, Suzuki K, Iijima K, Ebana K. Variation in cooking and eating quality traits in Japanese rice germplasm accessions., 2016, 66: 309–318.
[13] Bhat F M, Riar C S. Physicochemical, cooking, and textural characteristics of grains of different rice (L.) cultivars of temperate region of India and their interrelationships., 2016, 48: 160–170.
[14] Bao J S. Toward understanding the genetic and molecular bases of the eating and cooking qualities of rice., 2012, 57: 148–156.
[15] Patindol J A, Siebenmorgen T J, Wang Y J. Impact of environmental factors on rice starch structure: a review., 2015, 67: 42–54.
[16] Bello B K, Hou Y, Zhao J, Jiao G, Wu Y, Li Z, Wang Y, Tong X, Wang W, Yuan W, Wei X, Zhang J. NF-YB1-YC12-bHLH144 complex directly activatesto regulate grain quality in rice (L.)., 2019, 17: 1222–1235.
[17] Zhu D, Zhang H, Guo B, Xu K, Dai Q, Wei C, Zhou G, Huo Z. Effects of nitrogen level on structure and physicochemical pro-perties of rice starch., 2017, 63: 525–532.
[18] 劉立軍, 王康君, 卞金龍, 熊溢偉, 陳璐, 王志琴, 楊建昌. 水稻產量對氮肥響應的品種間差異及其與根系形態生理的關系. 作物學報, 2014, 40: 1999–2007. Liu L J, Wang K G, Bian J L, Xiong Y W, Chen L, Wang Z Q, Yang J C. Differences in yield response to nitrogen fertilizer among rice cultivars and their relationship with root morphology and physiology., 2014, 40: 1999–2007 (in Chinese with English abstract).
[19] Yang J, Zhang J, Wang Z, Zhu Q, Liu L. Activities of enzymes involved in sucrose-to-starch metabolism in rice grains subjected to water stress during filling., 2003, 81: 69–81.
[20] Zhu D, Wei H, Guo B, Dai Q, Wei C, Gao H, Hu Y, Cui P, Li M, Huo Z, Xu K, Zhang H. The effects of chilling stress after anthesis on the physicochemical properties of rice (L.) starch., 2017, 237: 936–941.
[21] Feng F, Li Y, Qin X, Liao Y, Siddique K H M. Changes in rice grain quality ofandtype varieties released in China from 2000 to 2014., 2017, 8: 1–7.
[22] 胡雅杰, 吳培, 邢志鵬, 錢海軍, 張洪程, 戴其根, 霍中洋, 許軻, 魏海燕, 郭保衛. 機插方式和密度對水稻主要品質性狀及淀粉RVA譜特征的影響. 揚州大學學報(農業與生命科學版), 2017, 38(3): 73–82. Hu Y J, Wu P, Xing Z P, Qian H J, Zhang H C, Dai Q G, Huo Z Y, Xu K, Wei H Y, Guo B W. Effect of different mechanical transplanted methods and plant density on grain quality and characteristic of starch RVA of rice.(Agric Life Sci Edn), 2017, 38(3): 73–82 (in Chinese with English abstract) .
[23] 季紅娟, 張小祥, 趙步洪, 鄭青松, 陳剛, 李育紅, 肖寧, 潘存紅, 吳云雨, 蔡躍, 李愛宏. 不同播期和密度對直播粳稻揚粳3012產量及品質的影響. 揚州大學學報(農業與生命科學版), 2020, 41(1): 85–90. Ji H J, Zhang X X, Zhao B H, Zheng Q S, Chen G, Li Y H, Xiao N, Pan C H, Wu Y, Cai Y, Li A H. Effect of different sowing date and density on yield and quality of Yangjing 3012 in direct-seeding rice.(Agric Life Sci Edn), 2020, 41(1): 85–90 (in Chinese with English abstract).
[24] Zhou T, Zhou Q, Li E, Yuan L, Wang W, Zhang H, Liu L, Wang Z, Yang J, Gu J. Effects of nitrogen fertilizer on structure and physicochemical properties of ‘super’ rice starch., 2020, 239: 1–11.
[25] Smith A M, Denyer K, Martin C R. What controls the amount and structure of starch in storage organs?, 1995, 107: 673–677.
[26] Smidansky E D, Meyer F D, Blakeslee B, Weglarz T E, Greene T W, Giroux M J. Expression of a modified ADP-glucose pyrophosphorylase large subunit in wheat seeds stimulates photosynthesis and carbon metabolism., 2007, 225: 965–976.
[27] Devi T A, Sarla N, Siddiq E A, Sirdeshmukh R. Activity and expression of adenosine diphosphate glucose pyrophosphorylase in developing rice grains: varietal differences and implications on grain filling., 2010, 178: 123–129.
[28] Mizuno K, Kimura K, Arai Y, Kawasaki T, Shimada H, Baba T. Starch branching enzymes from immature rice seeds., 1992, 112: 643–651.
[29] Koo S H, Lee K Y, Lee H G. Effect of cross-linking on the physicochemical and physiological properties of corn starch., 2010, 24: 619–625.
[30] Hurkman W J, McCue K F, Altenbach S B, Korn A, Tanaka C K, Kothari K M, Johnson E L, Bechtel D B, Wilson J D, Anderson O D, DuPont F M. Effect of temperature on expression of genes encoding enzymes for starch biosynthesis in developing wheat endosperm., 2003, 164: 873–881.
[31] Gu J F, Chen J, Chen L, Wang Z Q, Zhang H, Yang J C. Grain quality changes and responses to nitrogen fertilizer ofrice cultivars released in the Yangtze River Basin from the 1950s to 2000s., 2015, 3: 285–297.
[32] Asaoka M, Okuno K, Sugimoto Y, Kawakami J, Fuwa H. Effect of environmental temperature during development of rice plants on some properties of endosperm starch., 1984, 36: 189–193.
[33] Ahmed N, Tetlow I J, Nawaz S, Iqbal A, Mubin M, Nawazul Rehman M S, Butt A, Lightfoot D A, Maekawa M. Effect of high temperature on grain filling period, yield, amylose content and activity of starch biosynthesis enzymes in endosperm of basmati rice., 2015, 95: 2237–2243.
[34] Ahmed N, Maekawa M, Tetlow I J. Effects of low temperature on grain filling, amylose content, and activity of starch biosynthesis enzymes in endosperm of basmati rice., 2008, 59: 599–604.
[35] Cheng F, Zhong L, Zhao N, Liu Y, Zhang G. Temperature induced changes in the starch components and biosynthetic enzymes of two rice varieties., 2005, 46: 87–95.
[36] 劉鑫燕, 李娟, 劉雪菊, 張昌泉, 顧銘洪, 劉巧泉. 可溶性淀粉合成酶與稻米淀粉精細結構關系的研究進展. 植物生理學報, 2014, 50: 1453–1458. Liu X Y, Li J, Liu X J, Zhang C Q, Gu M H, Liu Q. Progress in the relationship between soluble starch synthases and starch fine structure in rice., 2014, 50: 1453–1458 (in Chinese with English abstract).
[37] Pandey M K, Rani N S, Madhav M S, Sundaram R M, Varaprasad G S, Sivaranjani A K, Bohra A, Kumar G R, Kumar A. Different isoforms of starch-synthesizing enzymes controlling amylose and amylopectin content in rice (L.)., 2012, 30: 1697–1706.
[38] 胡群, 夏敏, 張洪程, 曹利強, 郭保衛, 魏海燕, 陳厚存, 韓寶富. 氮肥運籌對缽苗機插優質食味水稻產量及品質的影響. 作物學報, 2017, 43: 420–431. Hu Q, Xia M, Zhang H C, Cao L Q, Guo B W, Wei H Y, Chen H C, Han B F. Effect of nitrogen application regime on yield and quality of mechanical pot-seedlings transplanting rice with good taste quality., 2017, 43: 420–431 (in Chinese with English abstract).
[39] 沈鵬, 羅秋香, 金正勛. 稻米蛋白質與蒸煮食味品質關系研究. 東北農業大學學報, 2003, 34: 378–381. Shen P, Luo Q X, Jin Z X. Relationship between protein content and the cooking and eating quality properties of rice grain., 2003, 34: 378–381 (in Chinese with English abstract).
[40] 沈鵬, 金正勛, 羅秋香, 金學泳, 孫艷麗. 氮肥對水稻籽粒淀粉合成關鍵酶活性及蒸煮食味品質的影響. 東北農業大學學報, 2005, 36: 561–566. Shen P, Jin Z X, Luo Q X, Jin X Y, Sun Y L. Effects of nitrogen fertilizer on the enzyme activities involved in the starch synthesis and eating and cooking quality in rice grain., 2005, 36: 561–566 (in Chinese with English abstract)
[41] 潘圣剛, 翟晶, 曹湊貴, 蔡明歷, 王若涵, 黃勝奇, 李進山. 氮肥運籌對水稻養分吸收特性及稻米品質的影響. 植物營養與肥料學報, 2010, 16: 522–527. Pan S G, Zhai J, Cao C G, Cai M L, Wang R H, Huang S Q, Li J S. Effects of nitrogen management practices on nutrition uptake and grain qualities of rice., 2010, 16: 522–527 (in Chinese with English abstract).
[42] Borrell A K, Garside A L, Fukai S, Reid D J. Grain quality of flooded rice is affected by season, nitrogen rate, and plant type., 1999, 50: 1399–1408.
Effects of machine transplanting density and panicle nitrogen fertilizer reduction on grains starch synthesis in good taste rice cultivars
CHEN Yun1,2, LIU Kun1, ZHANG Hong-Lu1, LI Si-Yu1, ZHANG Ya-Jun2, WEI Jia-Li2, ZHANG Hao1, GU Jun-Fei1, LIU Li-Jun1,*, and YANG Jian-Chang1
1Jiangsu Key Laboratory of Crop Genetics and Physiology / Jiangsu Co-Innovation Centre for Modern Production Technology of Grain Crops / Jiangsu Key Laboratory of Crop Genomics and Molecular Breeding, Yangzhou University, Yangzhou 225009, Jiangsu, China;2College of Bioscience and Biotechnology, Yangzhou University, Yangzhou 225009, Jiangsu, China
The effect of machine transplanting density on grain starch synthesis in good taste rice cultivars (GTRC) is still unclear. In the experiment, three representative GTRC of Nanjing 9108, Nanjing 5055, and Nanjing 46 in Jiangsu province were used as materials. Five machine transplanting densities of 33.3 × 104, 27.8 × 104, 23.8 × 104, 20.8 × 104, and 18.5 × 104hills hm–2were formed with the row spacing at 30 cm and the plant spacing of 10, 12, 14, 16, and 18 cm, respectively. The effects of machine transplanting density on yield and grain starch synthesis in above three rice cultivars were studied, and the regulation mechanism of halving normal nitrogen (N) rate of panicle nitrogen fertilizer on the starch synthesis was also observed. The results were as follows: (1) The grain yield of the three GTRC were highest at the transplanting density of 27.8 × 104hills hm–2. The amylose content at mature stage of the three GTRC decreased first and then increased with the increase in machine transplanting density, while the amylopectin content continued to decrease. When the machine transplanting density was controlled at 27.5 × 104–29.9 × 104hills hm–2, it was beneficial to improve the taste value of the above-mentioned GTRC. (2) With the increase in machine transplanting density, the activities of granule-bound starch synthase (GBSS) and ADP-glucose pyrophosphorylase (AGP) were increased in the early and middle stages of grain filling (8–20 days after heading), and were decreased rapidly at later grain filling stage. Machine transplanting density had no significant effect on starch branching enzyme (SBE) activity. Among them, the activities of GBSS and AGP in grains at 12–16 days after heading were significantly and positively correlated with the contents of amylose and total starch. (3) Compared with normal N rate of panicle nitrogen fertilizer, half normal N rate could significantly decrease the activities of GBSS and AGP in grains during the early and middle stages of grain filling, reduce the amylose content of rice, and further improve the taste value of GTRC. These results indicated that machine transplanting density could regulate the amylose content due to the activities variations of key enzymes involved in starch synthesis (GBSS and AGP) in the grains during the early and middle stages of grain filling, thus resulting in the difference of rice taste value. Reasonable machine transplanting density and panicle nitrogen fertilizer rate are helpful to further improve the eating quality of GTRC.
good taste rice cultivar; machine transplanting density; key enzymes involved in starch synthesis; taste value; amylose
10.3724/SP.J.1006.2021.02069
本研究由國家重點研發計劃項目(2016YFD0300502, 2017YFD0301206), 國家自然科學基金項目(31871557, 31671614), 江蘇省農業科技自主創新資金項目(cx(18)3007), 江蘇省作物遺傳生理重點實驗室開放課題(YSCL201807)和江蘇高校優勢學科建設工程項目資助。
This study was supported bythe National Key Research and Development Program of China (2016YFD0300502, 2017YFD0301206), the National Natural Science Foundation of China (31871557, 31671614), the Jiangsu Agriculture Science and Technology Innovation Fund (cx(18)3007), the Open Project of Jiangsu Key Laboratory of Crop Genetics and Physiology, and the Priority Academic Program Development of Jiangsu Higher Education Institutions.
劉立軍, E-mail: ljliu@yzu.edu.cn, Tel: 0514-87972133
E-mail:yunchen@yzu.edu.cn, Tel: 0514-87979320
2020-10-22;
2021-01-13;
2021-02-18.
URL: https://kns.cnki.net/kcms/detail/11.1809.S.20210216.1438.006.html
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
