小麥黃花葉病抗性鑒定和全基因組關聯分析
2021-06-09 03:45:18郭營孫俊生屈春艷郭寶晉趙巖李斯深
山東農業科學 2021年5期
郭營,孫俊生,屈春艷,2,郭寶晉,趙巖,李斯深
(1.山東農業大學/作物生物學國家重點實驗室,山東 泰安 271018;2.棗莊學院,山東 棗莊 277160)
小麥黃花葉病是由小麥黃花葉病病毒(wheat yellow mosaic virus,WYMV)引起的一種土傳病害,以土壤中的禾谷多黏菌(Polymyxa graminis)為傳毒介體。近年來,該病害在我國冬麥區發生日趨嚴重,已逐漸成為生產上的一種重要病害,主要發生在黃淮冬麥區、長江中下游冬麥區的部分地區和西南冬麥區的四川盆地等[1-3]。由于該病毒存在于禾谷多黏菌休眠孢子體內,而禾谷多黏菌休眠孢子堆壁厚,具有很強的抗逆性,因此很難用化學方法防治。鑒定篩選抗源、選育小麥黃花葉病抗性品種是控制該病害的根本途徑。國內學者已經進行了一些小麥抗黃花葉病種質的篩選工作,如孫炳劍等[4]對河南62個主推小麥品種進行了小麥黃花葉病的抗性評價,篩選出揚輻9311、寧麥9號等10個抗病品種;魏瑋等[5]對國內外527個小麥品種(系)進行小麥黃花葉病抗性鑒定,篩選出包括豐抗1號、石家莊72、鄭麥9023等10個抗病品種;劉麗娟等[6]對黃淮地區的145個小麥品種進行小麥黃花葉病抗性評價,篩選出濮優938、新麥208等70個免疫品種,豫麥47、邯6172等50個抗病品種。
小麥黃花葉病抗性是受多基因調控的數量性狀,遺傳基礎復雜,受基因型和環境效應影響顯著。連鎖分析和全基因組關聯分析已成為數量性狀基因定位和挖掘的重要途徑。前人利用F2、重組自交系(recombinant inbred lines,RIL)等遺傳分析群體,在2A、2D、3B、4D、5A和7B等染色體上檢測到多個小麥黃花葉病抗性的數量性狀位點(QTL)[7-12]。相較于連鎖分析,全基因組關聯分析(genome-wide association study,GWAS)以自然群體為材料,來源廣泛(如野生種、地方品種、現代品種和高代品系),基因多態性較高,無需構建雙親群體,具有發掘效率高、分辨率高且成本較低的優點,目前利用自然群體進行小麥黃花葉病抗性的研究較少。
單核苷酸多態性(single nucleotide polymorphism,SNP)是由單個核苷酸變異引起的DNA序列多態性,具有在基因組中數量多、分布廣、遺傳穩定性高、可自動化分析等優點。近年來,國內外科學家已開發出多款小麥SNP芯片,如9K、90K、55K、660K和820K芯片[13-18],為GWAS分析提供了高基因組覆蓋度的基因型分型數據,并在小麥產量、品質、抗病性等相關性狀基因的挖掘中得到了廣泛應用[19-23]。
本研究對我國黃淮麥區134份小麥品種(系)進行小麥黃花葉病抗性鑒定,以期篩選出抗病種質,為育種家提供優異抗源。結合90K SNP芯片基因型分型數據和病情指數,通過全基因組關聯分析,以期獲得顯著關聯的分子標記位點,為小麥黃花葉病抗性遺傳改良提供參考。
1 材料與方法
1.1 供試材料
試驗材料為我國黃淮麥區134個小麥品種(系)組成的自然品種群體(表1)。供試材料分別于2014—2015(E1)、2015—2016(E2)、2016—2017(E3)年度種植于山東省泰安市岱岳區小麥黃花葉病連續多年均勻發病的自然病圃。田間種植采用隨機區組設計,每年10月中上旬播種,每材料種植3行,行長1.5 m,行距25 cm,每行點播40粒種子,重復2次,常規田間管理。
1.2 小麥黃花葉病抗性鑒定
分別于2015年(3月14日)、2016年(3月4日、3月21日)、2017年(3月11日、3月21日)進行田間抗病性鑒定,每個材料調查全部植株。單株抗性鑒定分級參照劉偉華等[24]的指標:0級,無癥狀;1級,輕度花葉,葉片不產生梭狀條紋癥狀,植株不矮化;2級,花葉明顯,梭條或黃花葉癥狀占葉面積的1/2左右,植株輕度矮化;3級,嚴重花葉,梭條或黃花葉癥狀占葉面積的3/4左右,植株明顯矮化。根據單株病級鑒定結果,計算病情指數(disease index,DI),計算公式為:病情指數=Σ(各級病株數×各級代表值)/(調查總株數×最高級代表值)×100。每個供試材料在同一時間鑒定的2個重復DI抗性結果取平均值,同一年的兩次抗性結果以DI較大值作為該品種(系)該年度抗性結果。品種抗病性分級標準:抗病(R),DI<5%;中抗(MR),5%≤DI<15%;中感(MS),15%≤DI<25%;感病(S),DI≥25%[24]。
1.3 基因型分型
利用小麥90K SNP芯片[13]對品種群體進行基因型分型。在得到的多態性SNP中,去除最小等位基因頻率(MAF)小于5%、缺失率大于15%的SNP位點,7 357個SNP有遺傳位置信息,分布在21條染色體上,用于下一步的關聯分析[25]。
1.4 全基因組關聯分析
利用TASSEL 5.0軟件,采用混合線性模型(MLM),以主成分分析(PCA)和親緣關系(Kinship)作為協變量,結合7 357個SNP標記基因型數據,對134份小麥品種(系)的DI值進行關聯分析,設置閾值P<0.001,曼哈頓圖使用CMplot程序包完成(https://github.com/YinLiLin/RCMplot)。
1.5 數據分析
3年DI結果的最佳線性無偏預測(best linear unbiased prediction,BLUP)采用R版本3.6.1(https://www.r-project.org/)軟件進行限制最大似然(REML)分析。利用SPSS 17.0軟件對小麥黃花葉病發病指數進行表型數據分析。性狀廣義遺傳力計算公式為為遺傳方差和環境方差。
2 結果與分析
2.1 小麥黃花葉病的抗性鑒定
供試的134個小麥品種(系)對小麥黃花葉病的抗性存在明顯差異,并且同一品種(系)在不同年份發病情況也有一定差異(表1)。其中,抗病品種(系)27個,占供試材料的20.15%,包括煙農0428、煙99603、山農18、山農17、煙農836、濟麥20、煙農999、濟麥22、良星99、煙農21、藁優9618、山農23、泰農18、濟麥21、954(7)-8、濟南17、西農889、魯麥23、濰麥8號、LS3283、山農25、汶農5號、山農21、LS4942、郯麥98、臨旱822、昌樂5號;中抗品種(系)21個,占供試材料的15.67%,分別為陜627、山農45、萊州95021、汶農6號、山農29、M8008、新麥26、泰山21、魯麥21、泰山24、煙農19、山融3號、石新828、鑫麥289、山農664、煙農15、魯麥14、石麥15、石家莊8號、煙5072、山農8355;中感、感病品種(系)分別為21、65個,占供試材料的15.67%、48.51%。
通過對134份小麥品種(系)在不同環境下的病情指數進行分析,結果(表2)表明:變異系數范圍為87.18%~140.82%,廣義遺傳力為70.65%。方差分析結果表明,品種(系)間、不同年份間的差異是變異的主要來源,DI的基因型和環境效應值均在P≤0.001水平差異顯著。
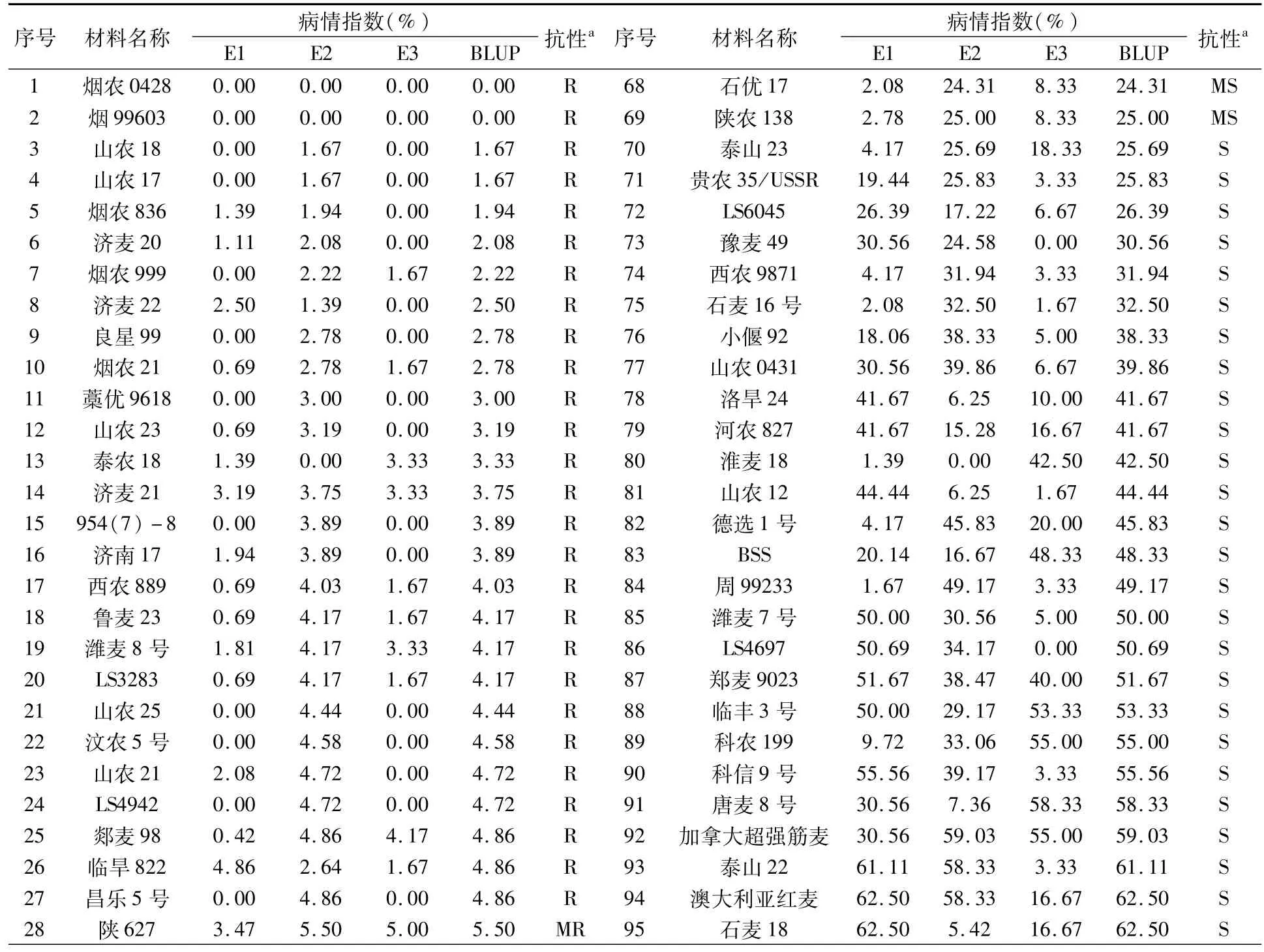
表1 134份小麥品種(系)的黃花葉病抗性表現

表1(續)

表2 134個品種(系)在不同環境下的病情指數統計分析
2.2 全基因組關聯分析
結果顯示,56個SNP與小麥黃花葉病病情指數顯著相關,分別位于2A(1個SNP)、2B(4個SNP)、2D(43個SNP)、3A(4個SNP)、4B(1個SNP)、7B(3個SNP)染色體上,可解釋8.8%~29.6%的表型變異(圖1,表3)。其中,在2D染色體上檢測到在3個環境和BLUP中均顯著關聯的SNP標記有40個,占所有關聯標記的71.4%。根據SNP遺傳位置[25],2D染色體的40個SNP位于76.57~86.28 cM區間(表3)。

圖1 全基因組關聯分析的曼哈頓圖

表3 與小麥黃花葉病病情指數顯著關聯的SNP標記
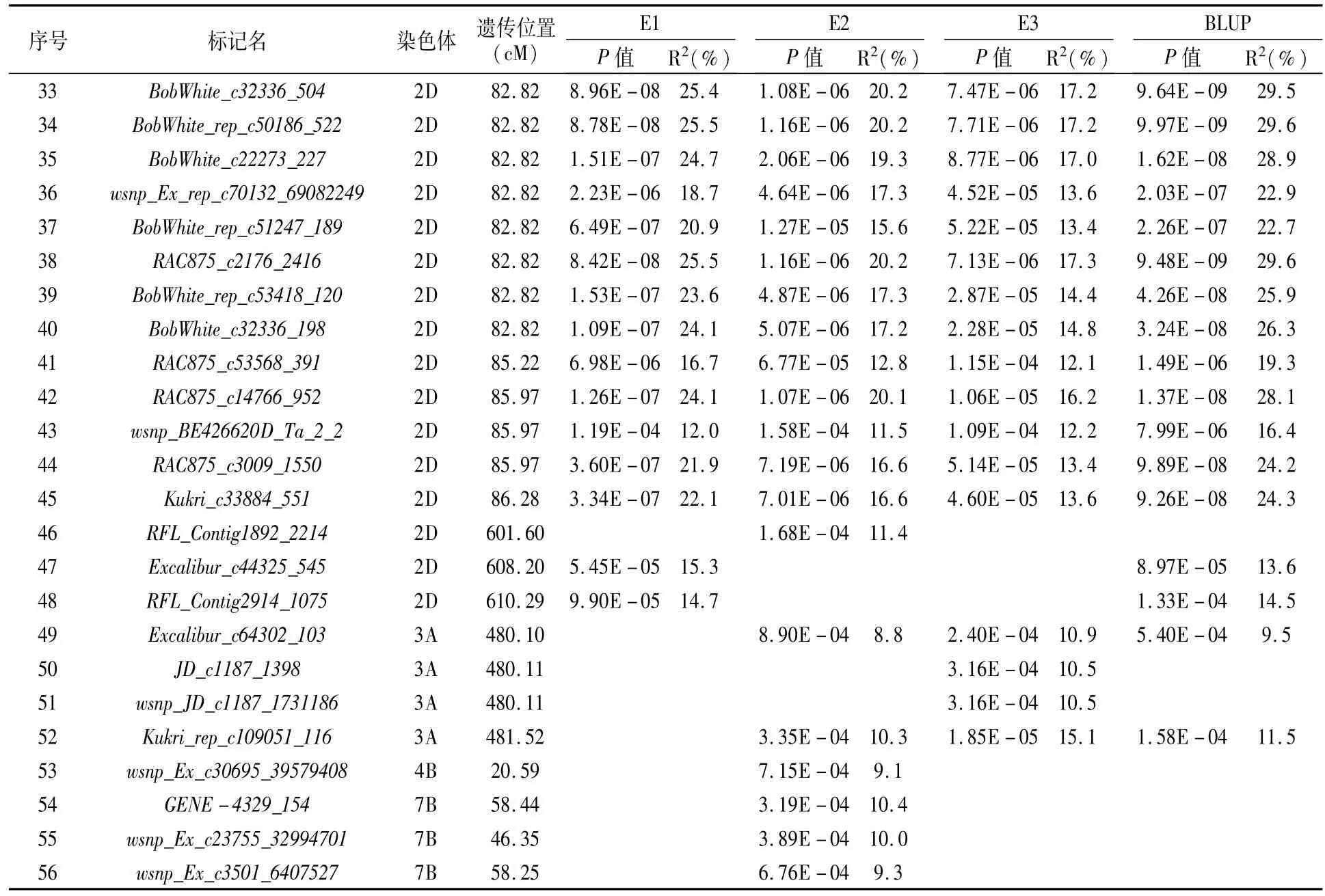
表3(續)
3 討論與結論
小麥黃花葉病廣泛發生于黃淮麥區、長江中下游麥區等冬小麥主要種植區域,由于感病品種的種植及農業機械的跨區作業,該病害的發病面積有逐漸擴大的趨勢。選育并推廣種植抗病品種是防治小麥黃花葉病大面積發生的最有效途徑。本研究連續3年對134個小麥品種(系)進行小麥黃花葉病的抗性鑒定,結果發現不同品種(系)對黃花葉病的抗性差異較大。本研究篩選出抗病品種(系)27個,包括目前生產上大面積種植和近年來審定的品種如山農18、山農17、煙農836、煙農999、濟麥22、良星99、煙農21、山農23、泰農18、濟南17、山農25、郯麥98等,建議在小麥黃花葉病主要發病麥區(黃淮麥區北片)推廣種植,也可作為抗病育種的親本材料加以利用。另外,篩選出21個中抗品種,可以通過與高抗品種進行輪作或者間作,達到延長抗病品種使用年限的目的。
本研究利用7 357個SNP標記與3年小麥黃花葉病病情指數及BLUP值進行關聯分析,共檢測到56個與小麥黃花葉病病情指數顯著關聯的SNPs,分別位于2A、2B、2D、3A、4B、7B染色體上;在2D染色體上檢測到在3個環境和BLUP中均顯著關聯的SNP標記40個,位于76.57~86.28 cM區間,表明上述標記基因的表達受環境影響較小,是穩定關聯的標記位點。前人的研究也在2D染色體檢測到抗病位點:如Takeuchi等[26]以高抗品種‘Madsen’和高感品種‘Hokushin’為親本構建F2群體,將‘Madsen’的抗病基因YmMD定位于2DL上,位于SSR標記wmc041和gwm349之間;Nishio等[8]用高抗品種‘Ibis’和高感品種‘Munstertaler’為親本構建DH群體,通過遺傳分析發現高抗品種‘Ibis’的抗病基因Ymlb位于2DL上,與SSR標記cfd16、wmc41、cfd168和wmc181緊密連鎖;Kobayashi等[12]用高抗品種‘Yumechikara’和高感品種‘Kitahonami’為親本構建DH群體,通過遺傳分析將抗病基因Q.Ymym定位于2D上,與SSR標記cfd233和gwm349緊密連鎖。由此可見,2D染色體含有較多抗黃花葉病基因,值得進一步深入研究利用。
猜你喜歡
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:30
當代陜西(2021年17期)2021-11-06 03:21:36
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
當代陜西(2019年15期)2019-09-02 01:52:00
電子制作(2018年18期)2018-11-14 01:48:24
學苑創造·A版(2018年11期)2018-02-01 06:29:20
讀者(2017年5期)2017-02-15 18:04:18
山東工業技術(2016年15期)2016-12-01 05:31:22
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
