低溫脅迫下北方強冬性區(qū)甘藍型冬油菜的低溫光合生理特征
2021-06-01 06:46:40鄒婭米文博徐明霞徐春梅劉自剛趙彩霞米超
甘肅農業(yè)大學學報 2021年2期
鄒婭,米文博,徐明霞,徐春梅,劉自剛,趙彩霞,米超
(1.甘肅農業(yè)大學農學院,干旱生境作物生物學國家重點實驗室,甘肅省作物遺傳改良與種質創(chuàng)新重點實驗室,甘肅 蘭州 730070;2.西藏自治區(qū)農牧科學院農業(yè)研究所,西藏 拉薩 850002)
我國北方旱寒區(qū)光照充足,氣溫冷涼,非常有利于油菜籽生產.目前我國北方旱寒區(qū)冬油菜種植以白菜型品種為主,種植效益低是目前北方寒旱區(qū)冬油菜發(fā)展的主要問題.白菜型油菜植株生長勢弱,籽粒產量低,品質差,莖稈軟,易倒伏,不宜機械化收獲;相對而言,甘藍型冬油菜,莖稈堅硬,抗倒伏,適合機械化收獲,且其生長勢強,籽粒產量高,品質優(yōu).然而,甘藍型冬油菜品種/系抗寒性較弱,不能抵抗北方強冬性區(qū)極端低溫,因此,培育甘藍型冬油菜抗寒品種/系是北方強冬性區(qū)能否種植甘藍型冬油菜的關鍵.冬性作物越冬是貯存的有機物質的消耗過程,冬前光合產物積累對越冬有重大影響.我國北方旱寒區(qū)冬油菜一般8月中下旬至9月上中旬播種,幼苗生長至 4~5 葉期即經歷明顯的降溫過程,隨后進入冬前低溫生長階段.冬油菜能否高效利用冬前低溫階段的光熱資源進行光合作用,積累充足的光合有機物,是越冬期根部抵御冬季嚴寒而安全越冬,以及翌年返青營養(yǎng)器官重建的關鍵.近年來,本課題組已育成多個強抗寒品種(系),在甘肅會寧(冬季極端低溫為-26 ℃)等強冬性區(qū)種植,越冬率可達90%以上,較好地解決了甘藍型冬油菜越冬問題.因此,研究低溫對甘藍型冬油菜光合生理變化,對北方旱寒區(qū)甘藍型冬油菜強抗寒品種應用有重要意義.
葉綠體是植物光合作用重要的細胞器,葉綠體結構的完整性會影響其光合功能.植物葉片受低溫脅迫后光合機構受損,光能利用率、光化學轉化效率下降,光合電子傳遞活性減弱.抗寒植物在一定低溫下光合膜較穩(wěn)定,相關保護性酶活性較高,光合能力較強,可為植物生長發(fā)育提供能量[1-3].研究表明,凈光合速率直接體現了植物光合能力,也是光合系統(tǒng)正常的檢驗指標之一[4],對于強抗寒品種,在低溫脅迫下葉綠體外膜完整清晰,基質片層結構較完整,其內仍有少量淀粉粒存在,而抗寒性弱的品種相鄰葉綠體融合,葉綠體外膜破裂,基質片層斷裂[5].
常溫條件下,植物細胞處于正常水分狀態(tài),活性氧產生后可被抗氧化酶系統(tǒng)迅速清除,從而整個代謝處于平衡狀態(tài)[6].大量研究表明,植物抗寒性強弱與抗氧化酶活性密切相關[7-8].低溫脅迫會引起植物膜脂過氧化,從而使植物受到冷傷害[9],研究表明丙二醛的含量可以作為衡量逆境對生物膜危害程度的指標[10-11],丙二醛含量與抗寒性密切相關.低溫脅迫下,白菜型冬油菜抗氧化酶活性升高,滲透調節(jié)物質含量增加,并且抗寒性不同的材料生理特性變化不同[12].
利用冬前低溫階段有限光熱資源進行光合產物積累,是作物抵御冬季嚴寒,安全越冬的關鍵.本研究以4份甘藍型冬油菜為材料,研究低溫脅迫對甘藍型冬油菜幼苗葉片生理特性、形態(tài)結構、葉綠體超微結構、光合特性的影響,為甘藍型冬油菜抗寒種質挖掘、抗寒性評價及品種篩選提供理論依據.
1 材料與方法
1.1 試驗設計
以TSG10、天油2266、天油2288、Vision 4個甘藍型油菜為材料(均由甘肅農業(yè)大學提供),選取籽粒飽滿、大小均一、無病蟲害的甘藍型冬油菜種子,用70%酒精消毒1 min,10%氯化鈉溶液消毒1 min,后用無菌水沖洗干凈,播于裝有育苗基質的盆缽中,每盆定苗4株.幼苗五葉期放入低溫光照培養(yǎng)箱進行低溫處理.光照條件為光/暗(25 ℃/20 ℃),常溫處理作為對照,-4 ℃處理24 h作為低溫處理組,分別觀察各品種幼苗形態(tài)及葉綠體超微結構變化,測定生理指標、光合氣體交換等指標.試驗設置3個生物學重復.
1.2 試驗方法
1.2.1 甘藍型冬油菜形態(tài)特征觀察 觀察并拍照記錄低溫脅迫前后甘藍型冬油菜幼苗葉片形態(tài)變化及受凍情況.
1.2.2 甘藍型冬油菜生理指標的測定 常溫作為對照組(CK),-4℃處理24 h作為低溫處理組.參照鄒琦[13]的方法,測定不同品系甘藍型冬油菜葉片超氧化物歧化酶 (superoxide dismutase,SOD)、過氧化物酶 (peroxidase,POD)、過氧化氫酶 (catalase,CAT)、APX、可溶性蛋白(soluble protein,SP)、可溶性糖(soluble sugar,SS)、丙二醛(malondialdehyde,MDA)、脯氨酸(proline,Pro)[14]、電導率等生理指標.
1.2.3 甘藍型冬油菜葉綠體超微結構的觀察 葉綠體超微結構觀察方法參照 Lichtenthaler等[15]和趙云等[16]的方法.分別取對照組和低溫處理組幼苗的第4片葉片觀察細胞超微結構.將葉片切成(2 mm×4 mm)碎片浸入2.5%戊二醛中,4 ℃下固定24 h.然后用0.1 mol/L磷酸緩沖液沖洗3次,置于4 ℃下1% 鋨酸中固定5 h,并置于0.1 mol/L磷酸鈉中漂洗25 min;將固定好的組織樣品依次放入50%、70%、80%、90%、100%丙酮中脫水;嵌入到Spurr培養(yǎng)基中,然后用Epon 812進行聚合后樹脂包埋.用含有醋酸鈾酰和雷諾檸檬酸鉛雙重染色8 min,TEM-1200EX透射電鏡觀察葉綠體超微結構.
1.2.4 光合氣體交換參數測定 低溫處理前后,采用 LI-6400 便攜式光合儀測定每個處理幼苗的第3片葉的葉片凈光合速率(Pn)、氣孔導度(Gs)、胞間CO2濃度(Ci)、蒸騰速率(Tr)等,葉室(2 cm×3 cm)內設定溫度 20 ℃,測定時間為上午 8∶30~10∶00,光強為 600 μmol/(m2·s);每個處理測定3株,每株測3次.
1.3 統(tǒng)計分析
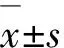
2 結果與分析
2.1 低溫對甘藍型冬油菜幼苗形態(tài)特征的影響
常溫下,甘藍型冬油菜葉片平展,直立生長;低溫處理(-4 ℃)24 h后,強抗寒品種TSG10植株呈半匍匐狀態(tài),葉色變化較小,仍保持了深綠色;抗寒性較弱的品種天油2288、天油2266葉片邊緣卷曲,3個抗寒性較弱的品種部分葉面顏色呈現暗紅色,葉片出現一定程度的失綠現象,表明低溫下甘藍型冬油菜強抗寒品種能更好維持光合色素的穩(wěn)定性(圖1).
2.2 低溫對甘藍型冬油菜抗氧化酶活性的影響
研究結果表明,低溫處理后不同品種甘藍型冬油菜幼苗抗氧化酶活性均有不同程度的升高(圖2),有效清除低溫逆境下體內積累的活性氧自由基,增強幼苗對低溫的抵抗能力.低溫處理后,TSG10、天油2266、天油2288和Vision SOD活性分別較對照增加了13.5%、52.7%、107.9%和109.5%;POD活性分別較對照增加了26.9%、70.0%、31.5%和31.0%;CAT活性分別較對照增加了14.4%、29.3%、22.1%和35.6%;APX活性分別較對照增加了59.7%、62.7%、59.7%和77.5%.方差分析表明:低溫處理后,TSG10和天油2266 POD活性差異顯著,而天油2288和Vision差異不顯著,4個品種的CAT活性差異均不顯著.綜上,低溫脅迫后甘藍型冬油菜葉片抗氧化酶活性均呈現上升趨勢,各品種間達到顯著差異(P<0.05)水平,品種間抗氧化酶活性上升幅度大致為:TSG10>天油2266>天油2288>Vision.

圖1 低溫脅迫下不同品種甘藍型冬油菜幼苗形態(tài)變化Figure 1 Morphological changes of different winter Brassica napus varieties seedlings under low temperature stress

圖中不同英文字母表示在不同處理間差異顯著(P<0.05).Different letters above histogram graphs indicate significant difference among each treatment (P<0.05).圖2 低溫脅迫下不同甘藍型冬油菜品種抗氧化酶活性變化Figure 2 Effect of low temperature on antioxidant enzyme activities of different winter rapeseeds
2.3 低溫處理對甘藍型油菜滲透調節(jié)物質含量的影響
SS、SP和Pro作為植物體內重要的滲透調節(jié)物質,經過低溫脅迫后含量上升,植株通過增加它們的含量來調節(jié)機體的滲透調節(jié)機制,降低機體內自由水的含量,增加機體對寒冷的抵抗能力.低溫脅迫后甘藍型冬油菜SS、SP、Pro含量均較對照呈顯著上升趨勢(圖3).方差分析表明:低溫處理后,4個品種的可溶性糖含量差異顯著;TSG10與其他3個品種SP的差異顯著;Pro含量較對照大幅度上升,品種間差異顯著.低溫脅迫后,TSG10、天油2266、天油2288和Vision可溶性糖含量分別較對照增加了200.5%、173.2%、128.2%和118.6%;SP含量分別增加了85.0%、107.4%、98.3%和81.6%;Pro含量分別增加了279.2%、280.5%、305.5%和289.5%.綜上,各品種滲透調節(jié)物質活性表現為TSG10>天油2266>天油2288>Vision.

圖中不同英文字母表示在不同處理間差異顯著(P<0.05).Different letters above histogram graphs indicate significant difference among each treatment (P<0.05).圖3 低溫脅迫前后不同甘藍型冬油菜品種滲透調節(jié)物質含量的變化Figure 3 Osmotic adjustment substances changes in different winter rapeseed varieties under low temperature stress
2.4 低溫處理對甘藍型油菜膜氧化損傷的影響
常溫下,不同甘藍型冬油菜品種間電導率變化幅度較小(圖4).低溫處理后,4個品種的電導率均顯著上升.其中上升幅度最大的為Vision,較對照增加了82.6%,其次是天油2288(64.2%)和天油2266(55.9%),上升幅度最小的是TSG10(44.2%).表明,低溫脅迫下,強抗寒品種TSG10電解質滲出較少,質膜完整性及通透性較好,質膜穩(wěn)定性明顯優(yōu)于冷敏感品種Vision.
丙二醛作為膜脂氧化的主要產物,其含量可以直接反應植物受逆境脅迫的程度.常溫下,甘藍型冬油菜不同品系 MDA 含量差異不明顯(圖4).低溫脅迫下,不同品種丙二醛含量均有所增加,但TSG10、天油2266和天油2288丙二醛含量較常溫增加不顯著,Vision含量增加顯著.表明在低溫脅迫過程中,抗寒性品種膜脂過氧化程度較小,丙二醛含量較低,葉片受凍害程度小.
2.5 低溫脅迫對甘藍型冬油菜葉綠體超微結構的影響
常溫下,葉綠體緊密分布在細胞壁內側,液泡膜和質膜完整,葉綠體形狀規(guī)則呈橢圓形,內含2~3個淀粉粒,基質片層排列有序(圖5).低溫處理后,液泡膜和質膜出現破裂現象,部分細胞質基質滲出,葉綠體內淀粉顆粒增多,且體積變大,部分葉綠體膜出現破裂,基質片層也出現斷裂現象.低溫處理后,TSG10大部分葉綠體仍整齊排布在細胞壁內側,個別葉綠體膜出現破裂,表明其葉綠體膜質結構輕微受損;天油2266中大部分葉綠體仍整齊排布在細胞壁內側,且葉綠體形狀變?yōu)榧忓N形,葉綠體基質片層出現斷裂,細胞內膜質系統(tǒng)出現損傷,淀粉粒體積較對照增大;天油2288葉綠體基質片層斷裂較嚴重,葉綠體內淀粉粒數目和體積均較對照顯著增加,細胞內膜質系統(tǒng)損傷較重;Vision葉綠體膜質損傷,液體外滲,葉綠體基質片層紊亂,大部分斷裂,淀粉粒顯著積累(圖5).低溫脅迫造成甘藍型冬油菜葉綠體光合膜結構損傷,相較于弱抗寒品種,強抗寒品種受低溫脅迫時葉綠體結構損傷較小.綜上,葉綠體中淀粉粒的積累速率可直接反應品種的抗寒性,抗寒性強弱表現與生理特性相吻合.

圖中不同英文字母表示在不同處理間差異顯著(P<0.05).Different letters above histogram graphs indicate significant difference among each treatment (P<0.05).圖4 低溫脅迫前后不同甘藍型冬油菜品種電導率與丙二醛含量的影響Figure 4 The electrical conductivity and malondialdehyde content changes in different winter rapeseed varieties under low temperature stress

圖5 低溫下甘藍型冬油菜葉綠體超微結構變化Figure 5 Changes of chloroplast ultrastructural in winter rapeseed under low temperature stress
2.6 低溫脅迫對甘藍型冬油菜光合參數的影響
低溫脅迫下,不同甘藍型冬油菜葉片Gs、Tr、Pn較對照明顯降低,而Ci較對照呈上升趨勢(圖6),不同甘藍型冬油菜品系Pn存在顯著差異(P<0.05).TSG10葉片Pn顯著高于其他3個品種,低溫脅迫后Pn降低幅度表現為:TSG10(69.49%)<天油2266(86.56%) 低溫脅迫下,甘藍型冬油菜葉片Ci較對照明顯升高.Ci上升幅度表現為TSG10(32.92%)>天油2266(41.03%)>Vision(41.36%)>天油2288(43.92%);Tr較對照明顯降低,降幅度表現為TSG10(72.87%) 圖中不同英文字母表示在不同處理間差異顯著(P<0.05).Different letters above histogram graphs indicate significant difference among each treatment (P<0.05).圖6 低溫脅迫前后不同甘藍型冬油菜品種光合氣體交換參數變化Figure 6 Effects of low temperature on photosynthetic gas exchange parameters in different winter rapeseed varieties under low temperature stress 低溫脅迫下植物體內抗氧化酶活性在一定程度上可以反應植物的抗寒性[17].低溫脅迫下,膜系統(tǒng)受損與生物氧自由基有關,植物體內會積累大量的活性氧自由基,會對生物體產生過氧化損害[18];同時,植物體內也會合成一些物質,維持植物體內活性氧等自由基的生成與清除平衡.SOD、POD、CAT是細胞膜保護酶,在維持這種動態(tài)平衡過程中起著至關重要的作用.酶活性越高,消除氧自由基的能力愈強,植物的抗逆性也較強[19].本研究中,甘藍型冬油菜幼苗感受低溫脅迫時,強抗寒品種體內的SOD、POD、CAT活性較高.這一結論也在白菜型冬油菜[2,22]和扁桃[21]的研究中得到應證.MDA作為膜脂氧化的主要產物,其積累的多少可以反映植物受逆境脅迫程度[23].本研究顯示,低溫脅迫下,弱抗寒品種MDA含量升高顯著,說明弱抗寒品種受低溫損傷更嚴重.因此MDA含量可作為評判冬油菜抗寒性強弱的指標之一.低溫脅迫下,植物體內SS、SP和Pro含量增加與植物抗寒性密切相關[24].本研究中,SS、SP和Pro含量均增加.SP可以積累于細胞中,引起滲透壓加大,有助于植物對低溫脅迫的適應性[25].低溫脅迫下SS積累,提高了細胞滲透勢,有利于細胞滲透調節(jié),保護膜結構[26].本文結果也在章錦濤等[27]和孔祥會等[28]的研究中得到印證.也有研究指出,脯氨酸含量不僅與抗寒性有關,也隨植株的生長發(fā)育發(fā)生變化.抗艷紅等[29]對馬鈴薯的研究中得出,當進入花期后脯氨酸含量明顯上升.這一結論是否適用于油菜還需進一步論證.低溫脅迫下,植物體內膜系統(tǒng)發(fā)生破裂,膜內物質就會滲出到周圍環(huán)境,從而導致細胞的電導率增大.電導率大小也反映植物受逆境損害程度.羅寧等[30]對低溫脅迫下玉米根系電導率的研究得出,低溫脅迫下,強抗寒品種能夠保持較高的質膜流動性及通透性.本研究中,抗寒性不同的甘藍型冬油菜在低溫脅迫下電導率呈顯著上升趨勢,弱抗寒性的甘藍型冬油菜品種上升幅度高于強抗寒性品種.包玲玲[31]對水稻的抗寒性研究中得出,電導率在低溫脅迫下越高,表示水稻葉片受到的低溫傷害越大,細胞膜受破壞程度和細胞受氧化損傷程度越高. 光合作用是植物物質積累與生理代謝過程,本研究結果表明,低溫脅迫下甘藍型冬油菜葉片凈光合速率顯著降低,較于強抗寒品系,弱抗寒品系凈光合速率下降幅度更大,機體積累的有機物少;同時顯微鏡觀察發(fā)現葉綠體結構發(fā)生改變,基粒溶解、基質片層斷裂,膜有序結構被破壞;膜及結合分子的有序性、膜囊內外電荷規(guī)則分布和膜電勢形成,影響光能捕獲和轉換、電子傳遞和光合磷酸化等,進而影響光合作用的高效進行[32-33];基粒垛堞結構和基質片層損失導致膜結合分子排列有序性的喪失和跨膜電勢建立困難,最終導致葉片Pn的降低.北方強冬性區(qū)冬油菜入冬后地上部分全部枯萎,越冬期根部抵御冬季寒冷,以及翌年返青進行地上部營養(yǎng)體重建,都需要根部冬前積累足夠的貯藏物.在冬油菜種植中也發(fā)現,越冬前地下根部直徑> 1 cm,地上著生8~10葉的植株,有更大幾率通過北方寒冷的冬季而安全越冬;相反,冬前植株和根直徑較小的植株,通常難以安全越冬. 低溫脅迫后,甘藍型冬油菜強抗寒品種較好維持了葉片光合色素穩(wěn)定性,植株向半匍匐狀態(tài)過渡;有較強抗氧化和滲透調節(jié)能力,較好維持了葉綠體膜完整性和基質片層的連續(xù)性,保持了較高的光合合成能力.依據低溫形態(tài)、生理及光合特征等,甘藍型冬油菜不同品種抗寒性表現為:TSG10>天油2266>天油2288>Vision.
3 討論
4 結論
