藏羊肺臟中HIF-1α的表達及其高原適應性研究
2021-05-29 01:52:10李帥帥張雨欣王子懿阿依木古麗阿不都熱依木
現代畜牧獸醫 2021年5期
李帥帥,張 晨,張雨欣,鄧 茵,王子懿,阿依木古麗·阿不都熱依木
(西北民族大學生命科學與工程學院,甘肅蘭州730030)
藏羊又稱藏系羊(Tibetan sheep),是我國特有的牛科品種。藏羊體質強壯、抗嚴寒、耐粗飼,廣泛分布于海拔3 000~5 000 m的青藏高原地區。海拔高度與大氣含氧量呈負相關性變化,海拔越高,大氣含氧量越低[1]。藏羊長期生活在高海拔、低氧、嚴寒的環境中。藏羊經過長期的生物進化,對低氧、高寒、氣候多變的環境有很強的適應性,是高原牧區重要的經濟支柱。
缺氧誘導因子,也稱低氧誘導因子(hypoxia inducible factor,HIF),普遍存在于哺乳動物細胞內,是一種異源二聚體,主要由HIF-α和HIF-β2個亞單位組成。研究發現,缺氧誘導因子是細胞內對缺氧適應性反應的一種關鍵轉錄調控因子[2]。HIF-1α的激活在缺氧早期促進下游基因,如促紅細胞生成素(erythropoietin,EPO)、血管內皮生長因子(vascular endothelial growth factor,VEGF)等的表達,促進組織缺氧時細胞的存活;但嚴重缺氧或長期缺氧則導致下游基因如Bcl-2相互作用蛋白3(BNIP3)和NIX的表達增加,促進細胞凋亡和壞死;同時也能通過穩定p53的表達,引起細胞的凋亡。大鼠心肌細胞缺氧會導致HIF-1αmRNA及蛋白水平均顯著增加[3]。HIF-α在血管豐富的心、肺等組織中表達較強,在血管內皮細胞、胎肺成纖維細胞系等血流豐富的組織細胞中含量較高,而在白細胞系中含量較低[4]。HIF在動物低氧適應方面起著重要作用,然而具體的分子機制還不清楚。因此,本試驗擬通過組織學染色,觀察不同海拔生活的藏羊肺臟組織結構特點,并用免疫組織化學SP法和實時熒光定量PCR法檢測其HIF-1α的表達差異,為進一步研究藏羊高原適應性機制提供參考。
1 材料與方法
1.1 樣品采集
藏羊樣品采自青海主月畜牧業屠宰場(海拔高度3 900 m)和甘南鑫旺屠宰場(海拔高度2 900 m)。選擇健康、1~2歲藏羊各5只,斷頸處死,檢查胸腔和腹腔無可視病理變化,取肺組織1 cm3,4%多聚甲醛固定,24 h后更換固定液復固定,保存備用。
1.2 試驗方法
染色切片每組隨機抽取5張切片,每張切片在400倍視野隨機選擇10個不同區域,用Image Proplus6.0軟件測量肺被膜厚度、肺泡大小、單位面積(1 mm2)內肺泡與肺泡囊數量、并測量肺泡隔厚度。Gomori醛品紅染色切片中測量肺被膜彈性纖維厚度,按(肺彈性纖維厚度/肺被膜厚度)×100%計算肺被膜彈性纖維百分比。免疫組織化學SP染色切片檢測免疫陽性產物光密度值(IOD)。
1.2.1 組織學染色
各組肺組織常規梯度酒精脫水,二甲苯透明,石蠟包埋,連續切片,厚度為5μm;一部分切片用于常規H.E染色和Gomori醛品紅染色;另一部分用于免疫組織化學染色。
Gomori醛品紅染色:酸化高錳酸鉀氧化5 min,2%草酸漂白1 min,醛品紅染色15 min,70%酒精洗滌3次;橙黃G溶液滴染30 s,水洗終止染色,常規脫水、透明,中性樹膠封片。
1.2.2 免疫組織化學染色
切片脫蠟至水,采用微波檸檬酸抗原液修復法使抗原修復[5-6],滴加A液室溫避光孵育10 min,阻斷內源性過氧化物酶,PBS緩沖液洗滌3次,每次5 min;滴加B液,室溫孵育30 min,加入一抗室溫孵育1 h(陰性對照組用PBS緩沖液替代一抗),PBS緩沖液洗滌3次,每次5 min;最后滴加D液室溫孵育20 min,DAB顯色,顯微鏡觀察;最后蘇木素復染,自來水反藍(復染細胞核),常規脫水、透明,中性樹膠封片。陽性呈黃褐色,陰性對照不著色。
1.3 數據統計與分析
數據采用SPSS 22軟件SSR法統計分析,數據用“平均值±標準差”表示,P<0.05表示差異顯著,P<0.01表示差異極顯著。
2 結果與分析
2.1 不同海拔藏羊肺組織結構差異(見圖1、表1、表2)
由圖1可知,藏羊肺被覆漿膜,主要由疏松結締組織構成,富含彈性纖維。藏羊肺泡上皮主要以單層扁平上皮為主,肺泡隔中含大量毛細血管。
由表1可知,較高海拔藏羊肺被膜厚度與較低海拔藏羊比較差異不顯著(P>0.05),但較高海拔藏羊被膜中彈性纖維含量極顯著高于較低海拔藏羊(P<0.01)。
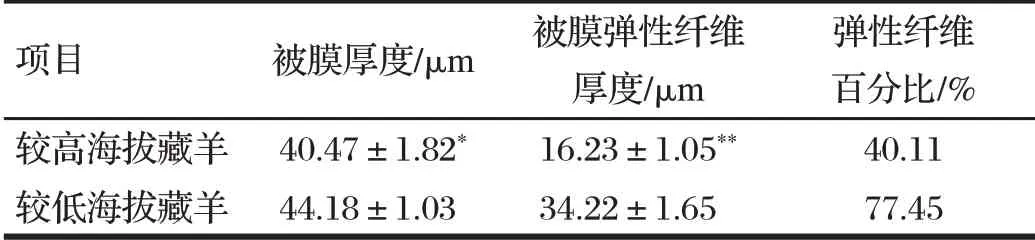
表1 不同海拔藏羊肺臟被膜厚度及彈性纖維含量比較Tab.1 Comparison of lung sac thickness and elastic fiber content of Tibetan sheep at different altitudes

表2 不同海拔藏羊肺泡及肺泡隔比較Tab.2 Comparison of alveolar and alveolar septum of Tibetan sheep at different altitudes
由表2可知,較高海拔藏羊與較低海拔藏羊肺泡大小、單位面積內肺泡數量均無顯著差異(P>0.05),但較高海拔藏羊單位面積內肺泡囊數量(1 mm2面積內)、肺泡隔厚度、肺泡隔中毛細血管數量均顯著高于較低海拔藏羊(P<0.05)。
2.2 不同海拔藏羊肺臟中HIF-1α的表達差異(見圖2)
由圖2可知,HIF-1α蛋白在肺臟細支氣管黏膜表達強度較強,微動脈內皮也有表達,肺泡隔中有少量表達。HIF-1α在細胞中主要位于細胞質中,細胞核內未見,較高海拔藏羊肺臟中HIF-1α蛋白的表達量高于較低海拔藏羊。
3 討論
高原動物在長期自然進化中為適應高原低氧環境,在形態學及生理學方面均表現出一些不同于平原動物的特點[7-8],尤其以心血管系統和呼吸系統的變化較為明顯。低氧是高原生存的不利因素之一。HIF是氧信號轉導系統的重要轉錄因子,在細胞的低氧應答中起著關鍵作用[9],通過調控靶基因,產生一系列應答反應,如增加紅細胞數量、調節葡萄糖代謝等。常氧下HIF蛋白也有表達,但很快被細胞內氧依賴性泛素蛋白酶降解,只有在缺氧條件下才會穩定表達[10]。動物在缺氧環境中,細胞內HIF-1α蛋白水平可在短時間內迅速上升,如牦牛[11]、藏雞[12]以及人類[13]等。肺被膜中豐富的彈性纖維可有效提高肺臟回縮力,有利于肺殘氣排出,提高肺通氣效率[14]。本試驗發現,較高海拔藏羊和較低海拔藏羊肺表面被膜厚度無顯著差異,但肺被膜中彈性纖維含量較高海拔藏羊顯著多于較低海拔藏羊,可能與藏羊能適應較高海拔有關。研究表明,肺泡中豐富的毛細血管可以顯著提高肺泡的交換效率[15]。本試驗發現,較高海拔藏羊肺泡隔和肺泡隔中毛細血管含量顯著高于較低海拔藏羊。
4 結論
試驗結果表明,HIF-1α蛋白大量分布于細支氣管黏膜上皮細胞和血管內皮細胞中,肺泡間隔有少量分布,且較高海拔藏羊中HIF-1α的表達顯著高于較低海拔藏羊,其參與調解低氧適應的機制仍需進一步研究。
